Chapter 5: Food, Fibre and Other Ecosystem Products

This chapter should be cited as:
Bezner Kerr, R., T. Hasegawa, R. Lasco, I. Bhatt, D. Deryng, A. Farrell, H. Gurney-Smith, H. Ju, S. Lluch-Cota, F. Meza, G. Nelson, H. Neufeldt, and P. Thornton, 2022: Food, Fibre, and Other Ecosystem Products. In: Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [H.-O. Pörtner, D.C. Roberts, M. Tignor, E.S. Poloczanska, K. Mintenbeck, A. Alegría, M. Craig, S. Langsdorf, S. Löschke, V. Möller, A. Okem, B. Rama (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 713–906, doi:10.1017/9781009325844.007.
Executive Summary 
Current Impacts
Climate change impacts are stressing agriculture, forestry, fisheries and aquaculture, increasingly hindering efforts to meet human needs (high confidence1 ). Human-induced warming has slowed growth of agricultural productivity over the past 50 years in mid and low latitudes (medium confidence). Crop yields are compromised by surface ozone (high confidence). Methane emissions have negatively impacted crop yields by increasing temperatures and surface ozone concentrations (medium confidence). Warming is negatively affecting crop and grassland quality and harvest stability (high co nfidence). Warmer and drier conditions have increased tree mortality and forest disturbances in many temperate and boreal biomes (high confidence), negatively impacting provisioning services (medium confidence). Ocean warming has decreased sustainable yields of some wild fish populations (high confidence). Ocean acidification and warming have already affected farmed aquatic species (high co nfidence). {5.2.1, 5.4.1, 5.5.1, 5.6.1, 5.7.1, 5.8.1, 5.9.1}
Warming has altered the distribution, growing area suitability and timing of key biological events, such as flowering and insect emergence, impacting food quality and harvest stability (high confidence). It is very likely 2 that climate change is altering the distribution of cultivated, wild terrestrial, marine and freshwater species. At higher latitudes, warming has expanded potential area but has also altered phenology (high confidence), potentially causing plant–pollinator and pest mismatches (medium confidence). At low latitude, temperatures have crossed upper tolerance thresholds, more frequently leading to heat stress (high co nfidence). {5.4.1, 5.7.4, 5.8.1, Cross-Chapter Box MOVING PLATE this chapter , 5.12.3.4}
Climate-related extremes have affected the productivity of all agricultural and fishery sectors, with negative consequences for food security and livelihoods (high confidence). The frequency of sudden food production losses has increased since at least mid-20th century on land and sea (medium evidence, high agreement ). Droughts, floods and marine heatwaves contribute to reduced food availability and increased food prices, threatening food security, nutrition and livelihoods of millions (high confidence). Droughts induced by the 2015–2016 El Niño, partially attributable to human influences (medium confidence), caused acute food insecurity in various regions, including eastern and southern Africa and the dry corridor of Central America (high confidence). In the northeast Pacific, a recent 5-year warm period impacted the migration, distribution and abundance of key fish resources (high confidence). Increasing variability in grazing systems has negatively affected animal fertility, mortality and herd recovery rates, reducing livestock keepers’ resilience (medium confidence). { 5.2.1, 5.4.1, 5.4.2, 5.5.2,5.8.1, 5.9.1, 5.12.1, 5.14.2, 5.14.6, Cross-Chapter Box MOVING PLATE this chapter; WGI AR6 Sections 11.2–11.8}
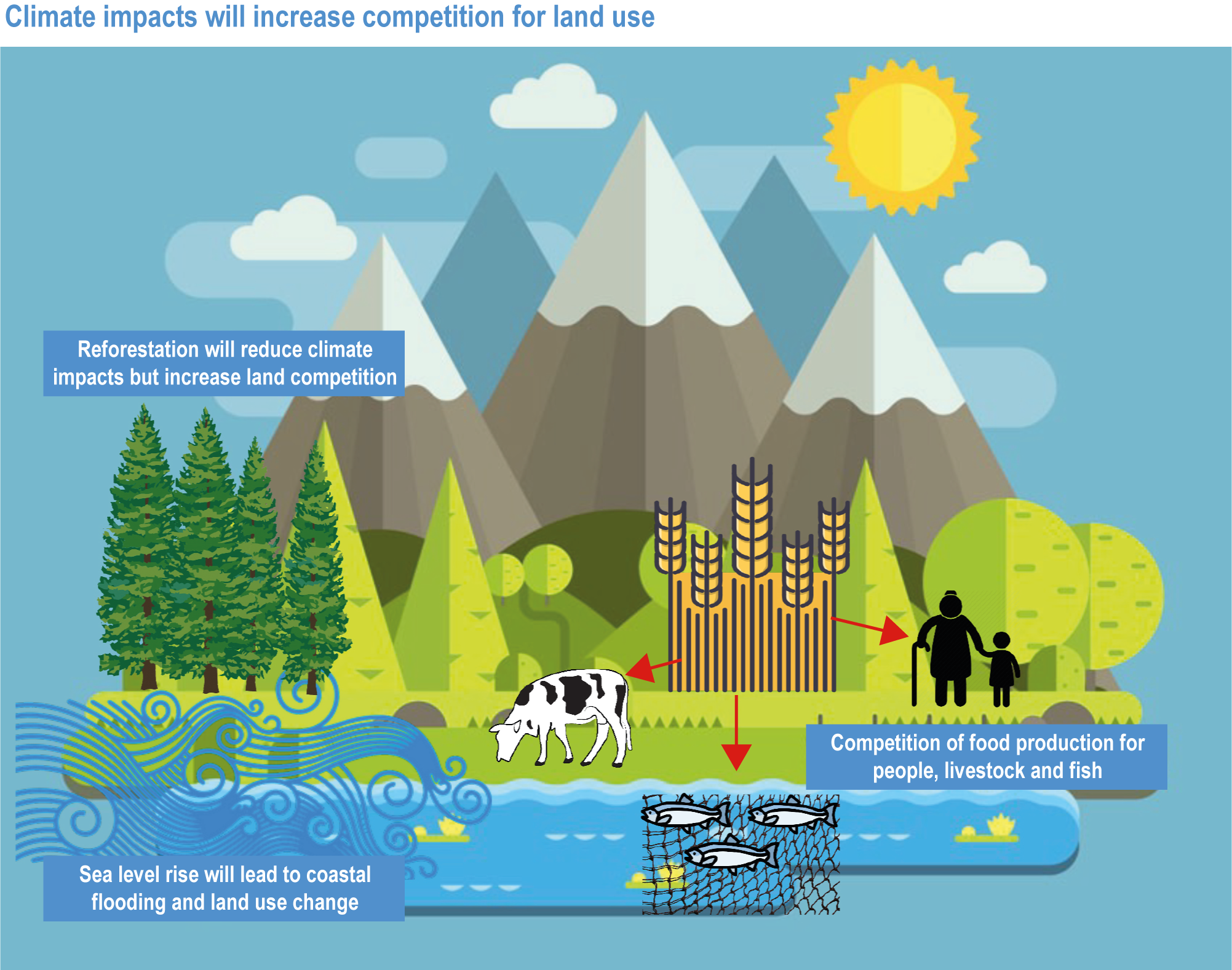
Climate change impacts everybody, but vulnerable groups, such as women, children, low-income households, Indigenous or other minority groups and small-scale producers, are often at higher risk of malnutrition, livelihood loss, rising costs and competition over resources (high confidence). Increasing competition for land, energy and water exacerbates impacts of climate change on food security (high confidence). {5.4.2.2, 5.5.2.6; 5.8.2.2, 5.9.2.1, 5.12.2, 5.12.3.1; 5.12.3.2; 5.12.3.3; 5.13.1, 5.13.3, 5.13.4}
Projected Impacts
Climate change will make some current food production areas unsuitable (high confidence). Current global crop and livestock areas will increasingly become climatically unsuitable under a high-emission scenario (high confidence) (e.g., 10% by 2050, over 30% by 2100 under SSP-8.5 versus below 8% by 2100 under SSP1-2.6). Increased, potentially concurrent climate extremes will periodically increase simultaneous losses in major food-producing regions (medium confidence). {5.2.2, 5.4.1, 5.4.3, 5.5.2, 5.5.3, Cross-Chapter Box MOVING PLATE in this chapter, <a class='section-link' data-title='Projected Impacts on Food Security' href='/report/ar6/wg2/chapter/chapter-5#5.12.4'>Section 5.12.4</a>; WGI Section 11.8}
Impacts on food availability and nutritional quality will increase the number of people at risk of hunger, malnutrition and diet-related mortality (high confidence). Climate change will increase the number of people at risk of hunger in mid-century, concentrated in Sub-Saharan Africa, South Asia and Central America (high confidence) (e.g., between 8 million under SSP1-6.0 and 80 million people under SSP3-6.0). Increased CO2 concentrations will reduce nutrient density of some crops (high confidence). Climate change will increase loss of years of full health 3 by 10% in 2050 under Representative Concentration Pathway (RCP) 8.5 because of undernutrition and micronutrient deficiencies (medium evidence, high agreement ). {5.2.2, 5.4.2, 5.4.3, 5.12.1.2, 5.12.4; Cross-Chapter Box MOVING PLATE this chapter}
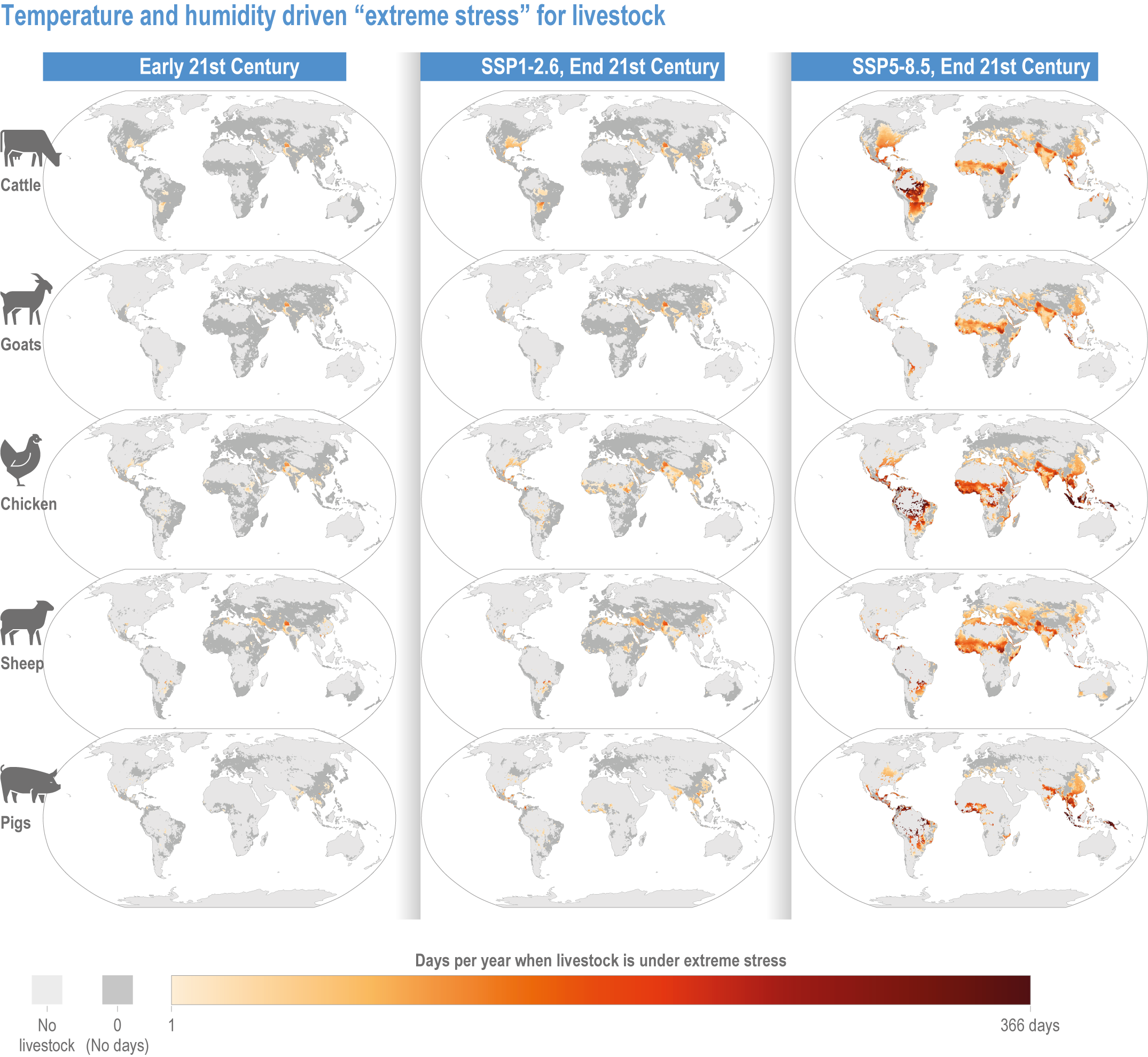
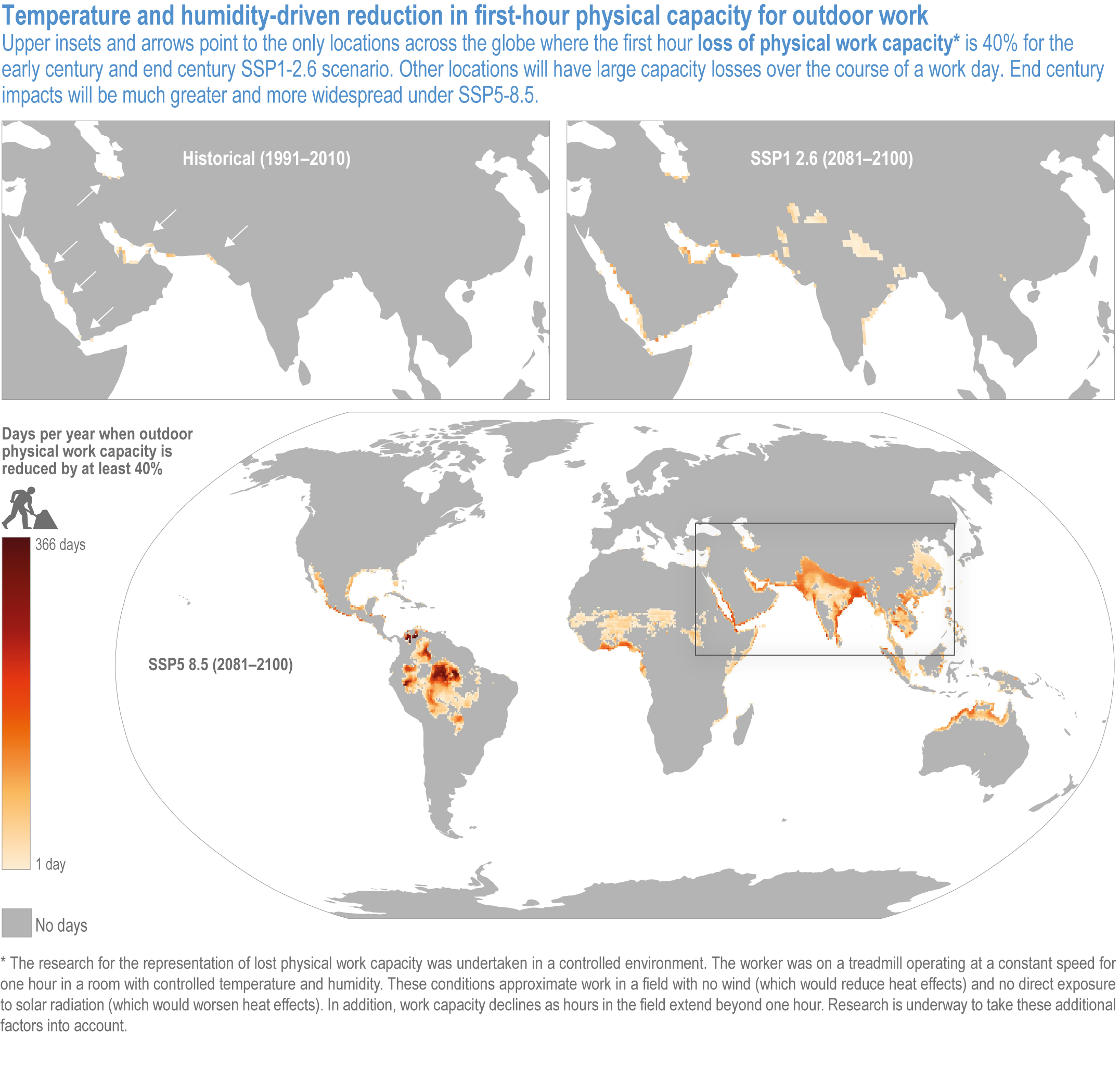
Climate change will increasingly expose outdoor workers and animals to heat stress, reducing labour capacity, animal health, and dairy and meat production (high confidence). The number of days with climatically stressful conditions for outdoor workers will increase by up to 250 workdays per year by century’s end in some parts of South Asia, tropical sub-Saharan Africa and parts of Central and South America under Shared Socioeconomic Pathway (SSP) 5-8.5, with negative consequences such as reduced food productivity, higher costs and prices (medium confidence). From early- to end-century, cattle, sheep, goats, pigs and poultry in the low latitudes will face 72–136 additional days per year of extreme stress from high heat and humidity under SSP5-8.5. Meat and milk productivity will be reduced (medium confidence). {5.5.3.4; 5.12.4}
Climate change will further increase pressures on terrestrial ecosystem services supporting global food systems (highconfidence). Climate change will reduce the effectiveness of pollinator agents as species are lost from certain areas, or the coordination of pollinator activity and flower receptiveness is disrupted in some regions (high confidence). Greenhouse-gas emissions will negatively impact air, soil and water quality, exacerbating direct climatic impacts on yields (high co nfidence). {5.4.3, Box 5.3, Box 5.4, 5.5.3.4; 5.7.1, 5.7.4, 5.10.3}
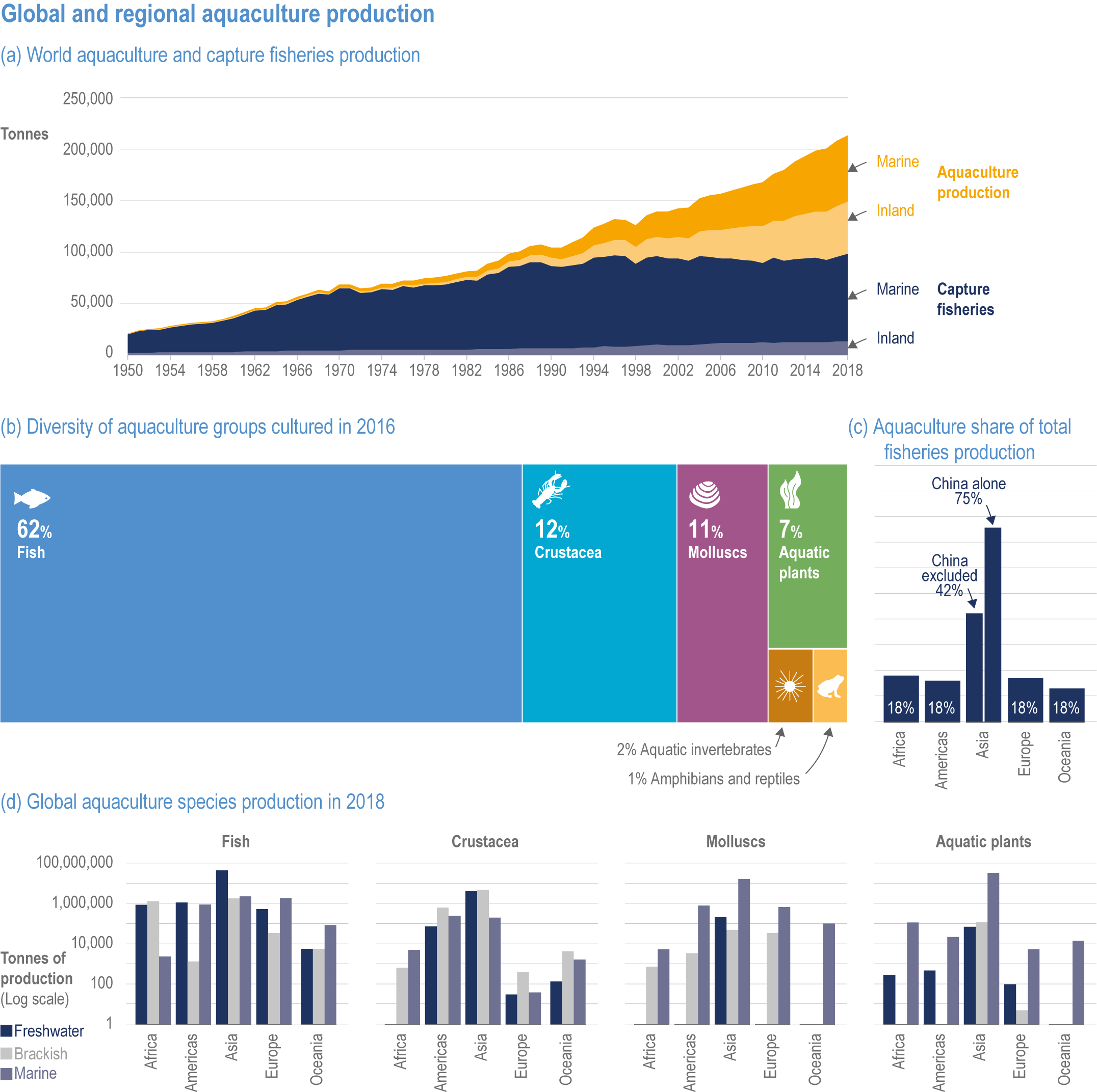
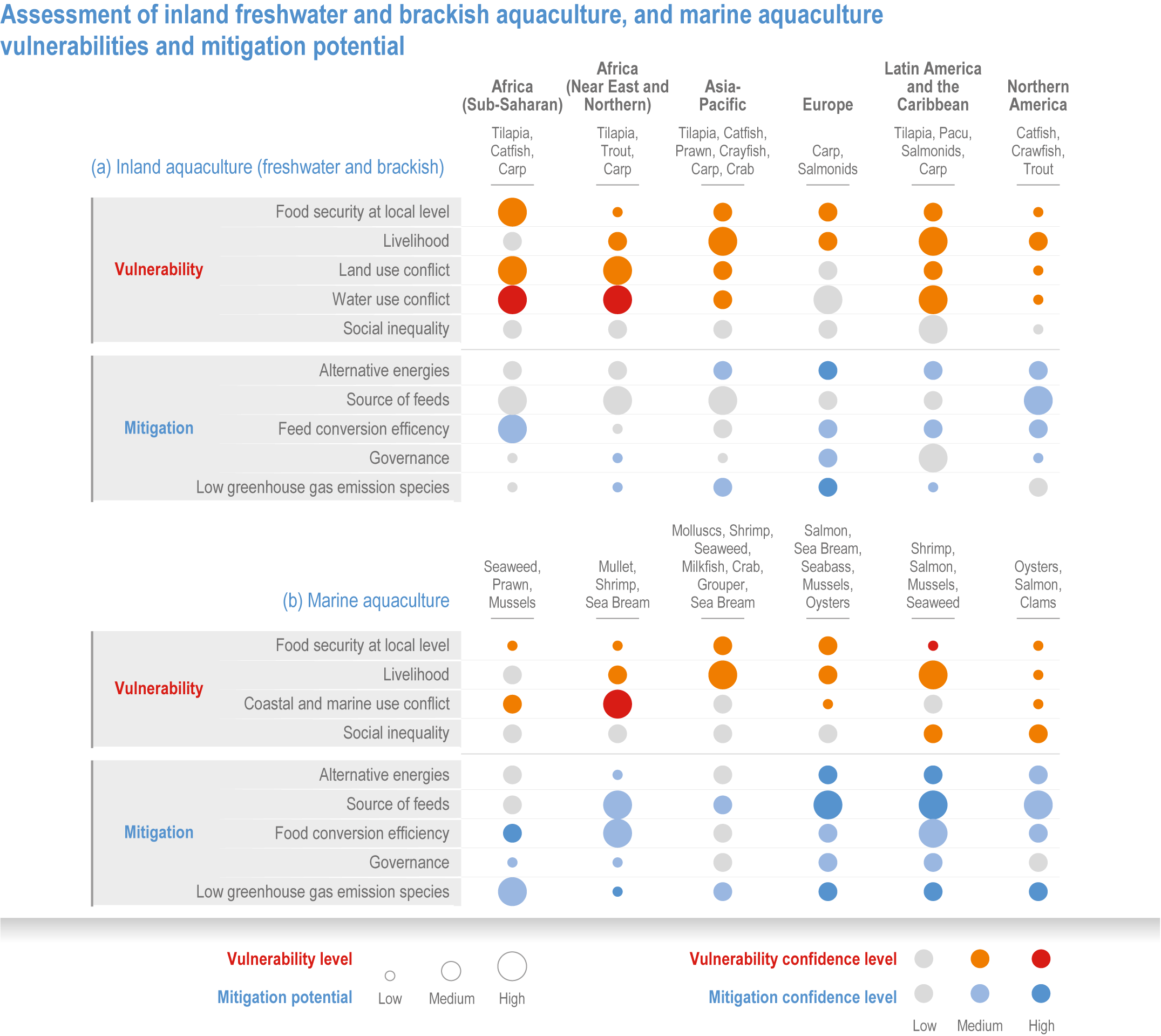
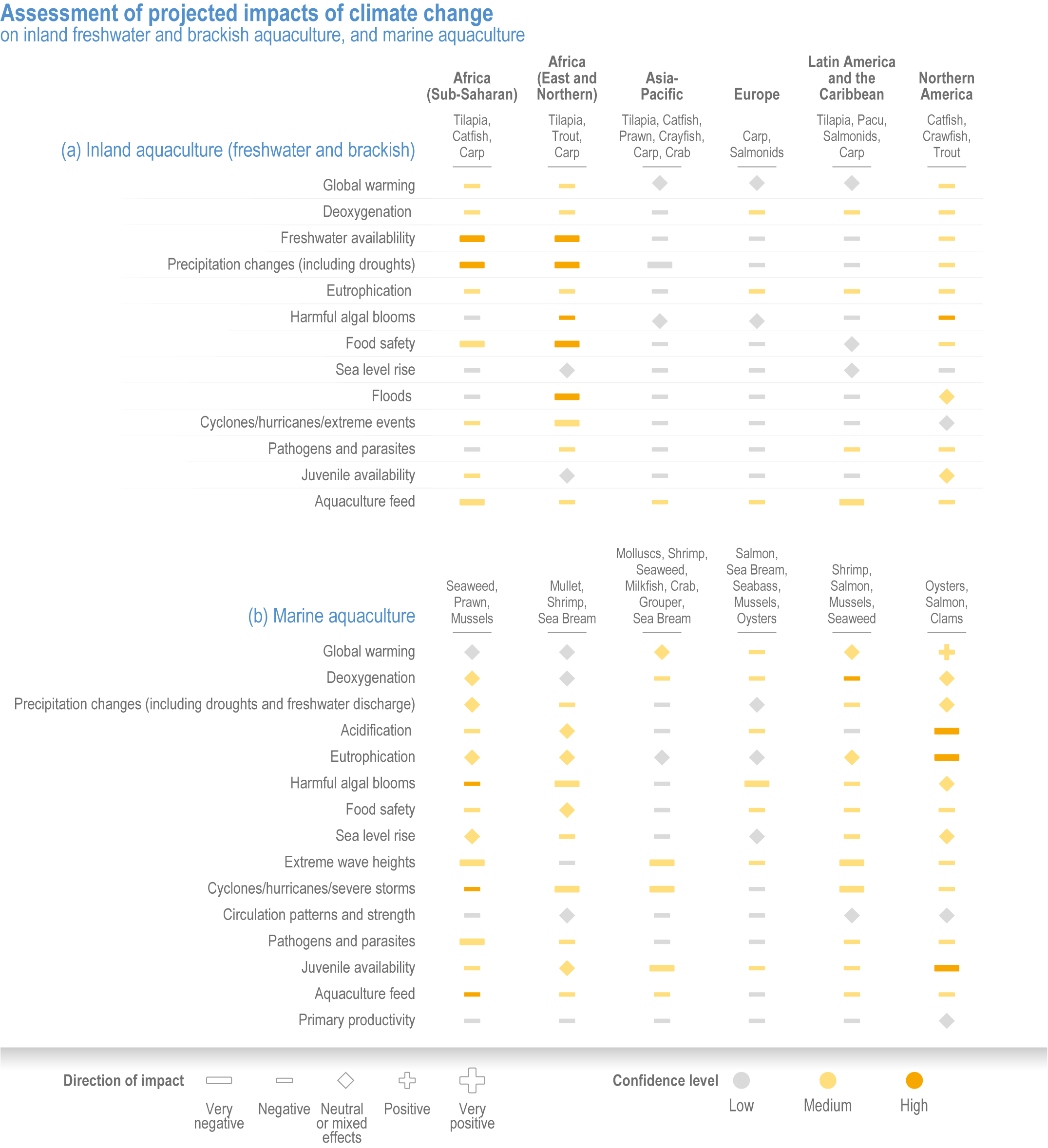
Climate change will significantly alter aquatic food provisioning services and water security with regional variances (high confidence). Climate change will reduce marine fisheries and aquaculture productivity, altering the species that will be fished or cultured, and reducing aquaculture habitat in tropical and subtropical areas (high confidence). Global ocean animal biomass will decrease by 5–17% under RCP2.6 and 8.5, respectively, from 1970 to 2100 with an average decline of 5% for every 1°C of warming, affecting food provisioning, revenue value and distribution (medium confidence). Global marine aquaculture will decline under warming and acidification from 2020 to 2100, with potential short-term gains for temperate finfish and overall negative impacts on bivalve aquaculture from habitat reduction (50–100% for some countries in the Northern Hemisphere) (medium confidence). Changes in precipitation, sea level, temperature and extreme climate events will affect food provisioning from inland and coastal aquatic systems (high confidence). Sea level rise and altered precipitation will increase coastal inundation and water conflicts between water-dependent sectors, such as rice production, direct human use and hydropower (medium confidence). {5.8.3, 5.9.3, 5.13, Cross-Chapter Box SLR in Chapter 3}
The occurrence and distribution of pests, weeds and diseases, including zoonoses, in agricultural, forest and food systems (terrestrial and aquatic) will be altered, and their control will become costlier (medium confidence). Changes in the rates of reproduction and distribution of weeds, insect pests, pathogens and disease vectors will increase biotic stress on crops, forests and livestock, and will increase the risk of biodiversity loss and ecosystem degradation (medium evidence, high agreement ). Risks will increase for climate-driven emerging zoonoses (medium evidence, high agreement ). {5.4.1.3, 5.9.4, Cross-Chapter Box MOVING PLATE this chapter}
Forest production systems will have variable responses to climate change across regions, with negative effects being more predominant in tropical forests (high confidence). In temperate and boreal regions, some productivity gains are projected, but tree mortality will increase in some areas (high confidence). In tropical forests, change in species composition and forest structure will lower production (medium confidence). Some models project a possible increase in global wood supply and lowering of average wood prices, but they do not account for the negative impacts of extreme events and thus possibly overestimate the wood supply (medium confidence). {5.6.2}
Climate change will negatively impact food safety (high confidence). Higher temperatures and humidity will favour toxigenic fungi, plant and animal-based pathogens, and harmful algal blooms (HABs) (high confidence). More frequent and intense flood events and increased melting of snow and ice will increase food contamination (high confidence). Incidence and severity of HABs and water-borne diseases will increase, as will indirect effects from infrastructure damage during extreme events (high confidence). {5.4.3, 5.5.2.3, 5.8.1, 5.8.2, 5.8.3, 5.9.1, 5.11.1, 5.11.3, 5.12.3; Cross-Chapter Box ILLNESS in Chapter 2}
Adaptation
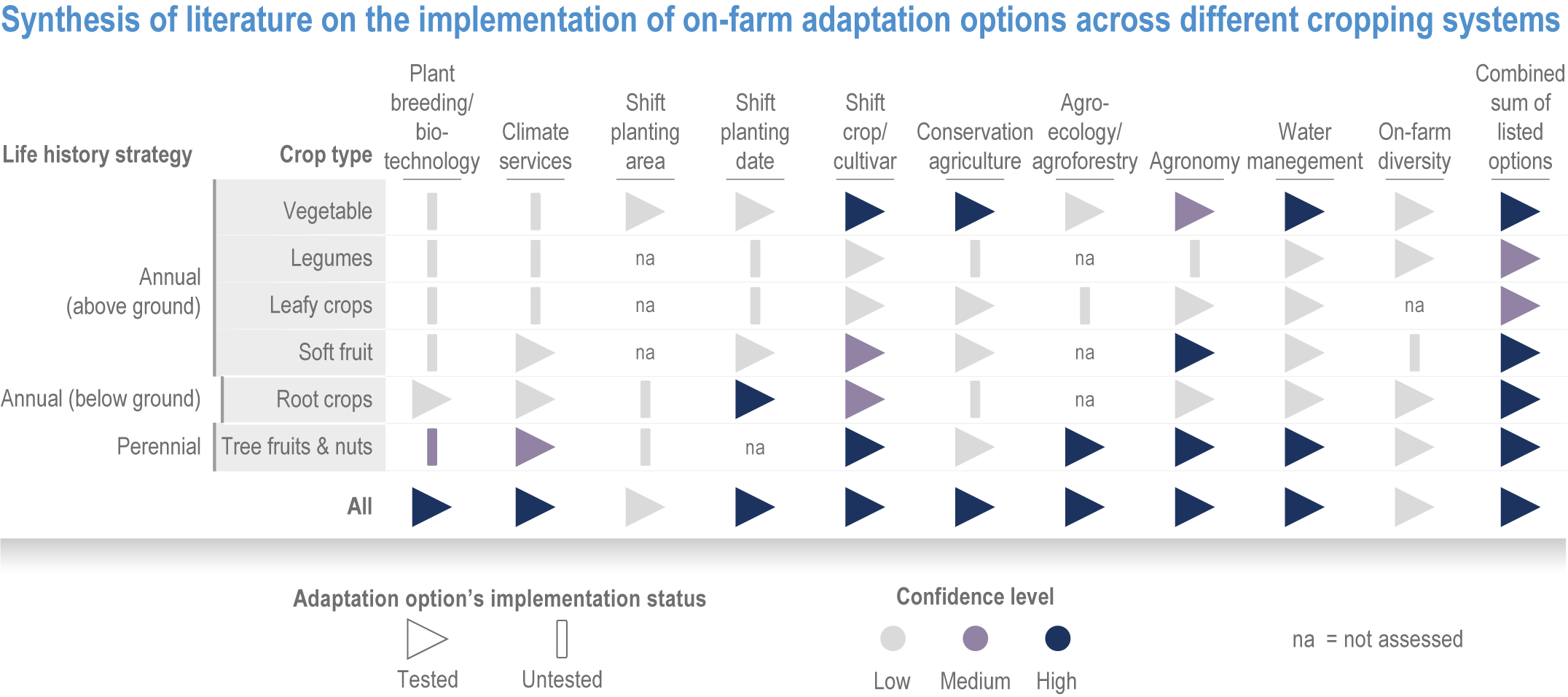
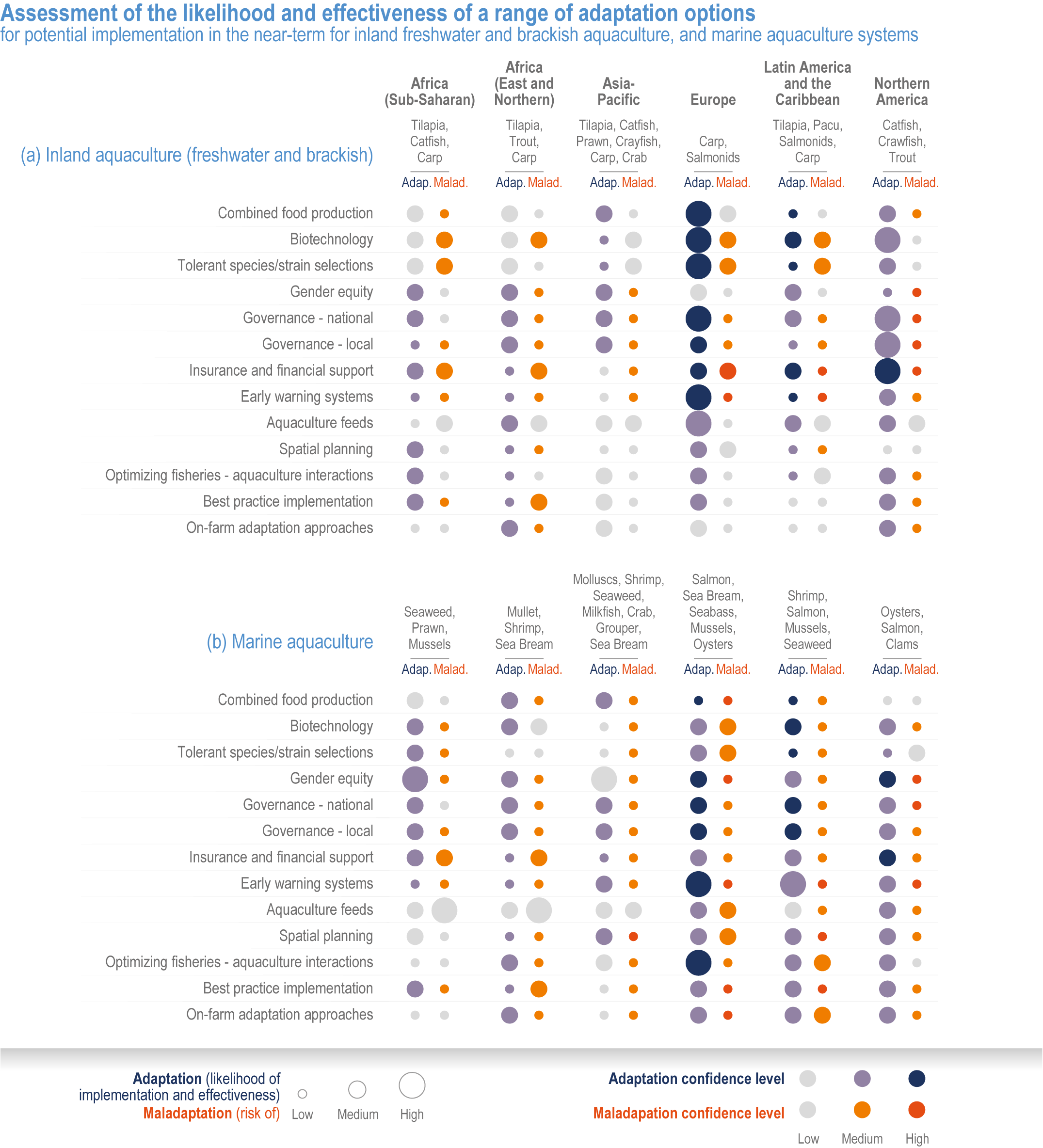
Many autonomous adaptation options have been implemented in both terrestrial and aquatic systems, but on-farm adaptations are insufficient to meet Sustainable Development Goal (SDG) 2 (high confidence). Autonomous responses include livestock and farm management, switching varieties/species and altered timing of key farm activities such as planting or stocking (high confidence). However, because of limited adaptive capacities and non-climatic compounding drivers of food insecurity, SDG2 will not be met (high confidence). {Table 5.1, 5.4.4; 5.5.4, 5.9.4, 5.10.4; 5.12.4}
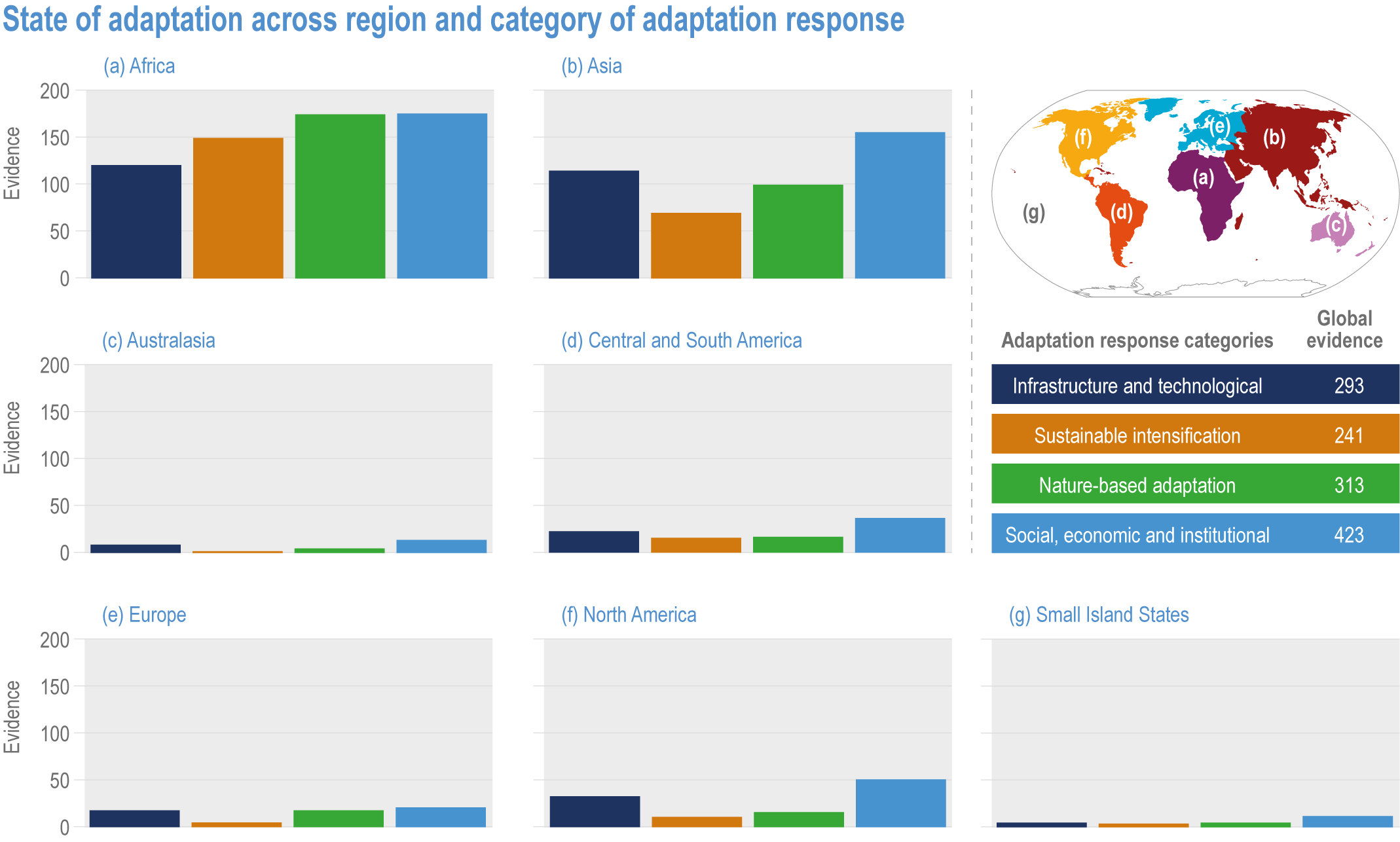
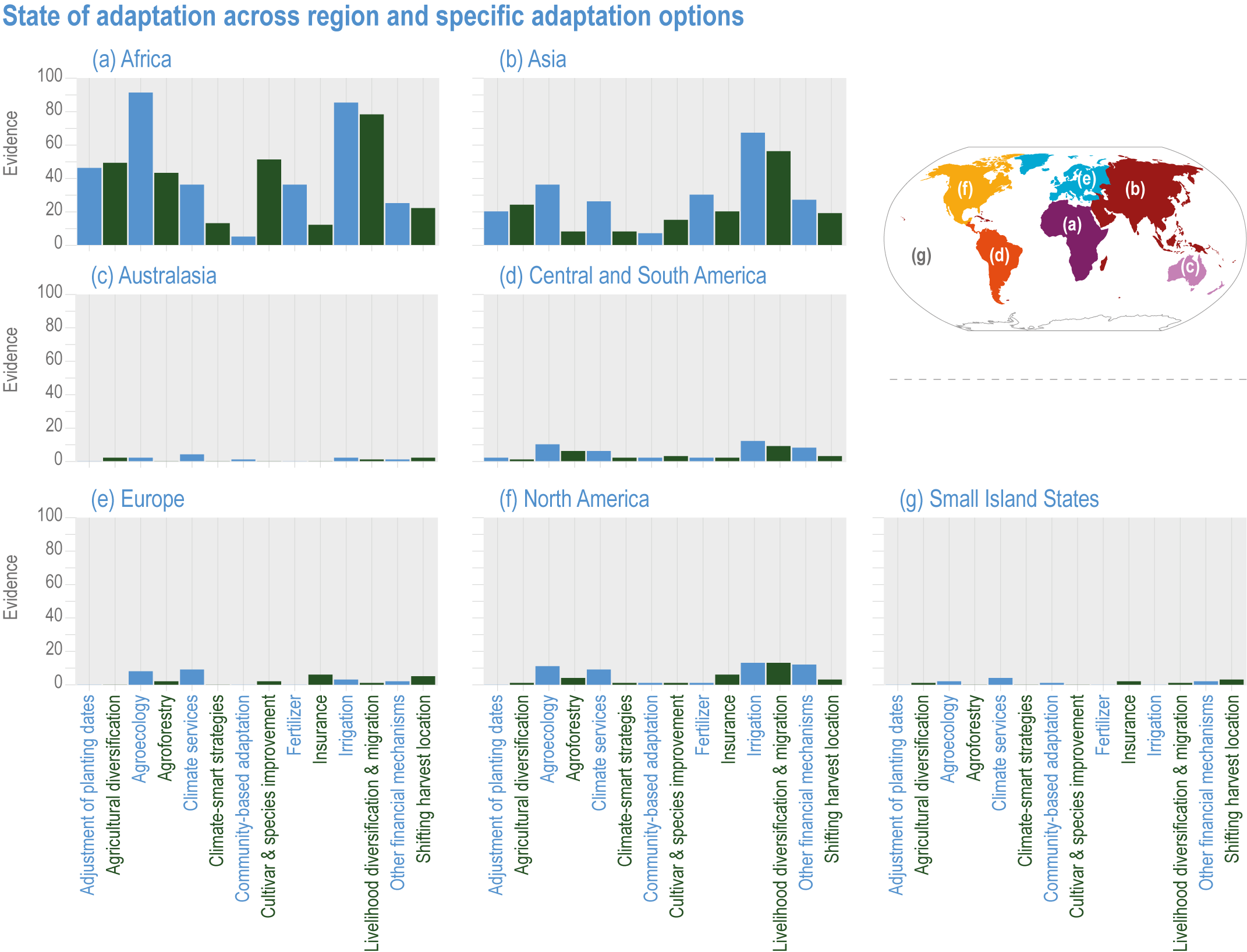
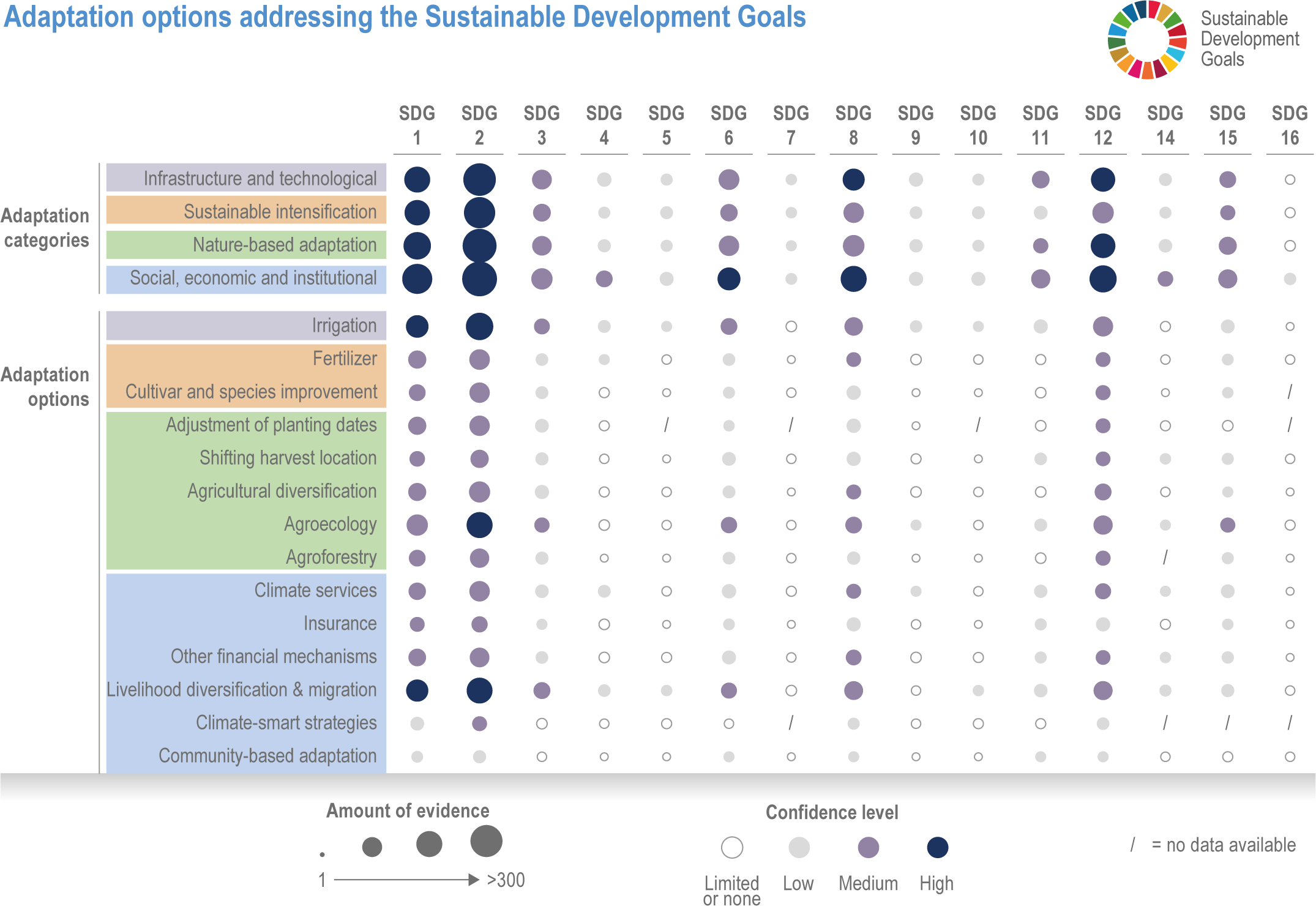
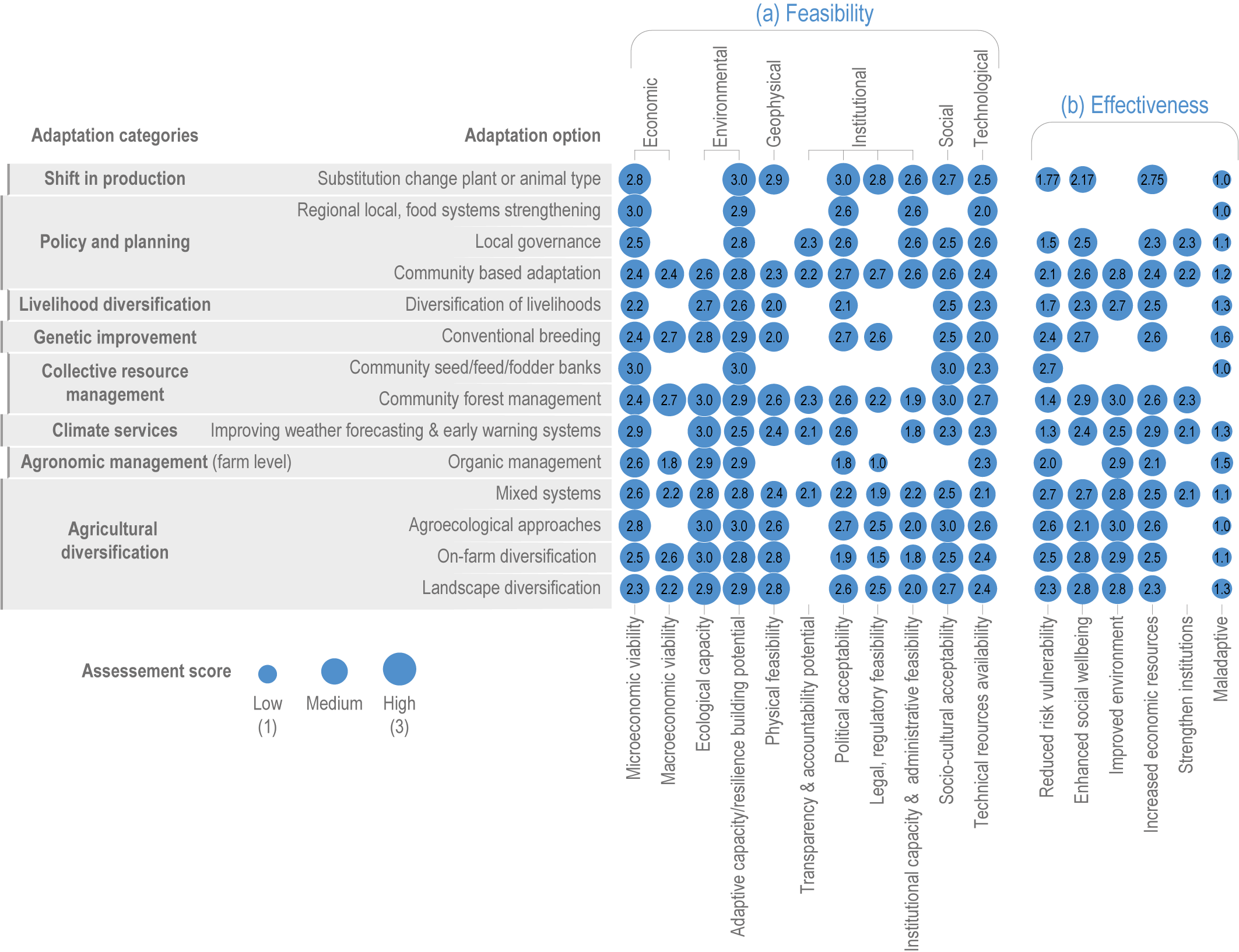
Various adaptation options are currently feasible and effective at reducing climate impacts in different socio-cultural, economic and geographical contexts (high confidence), but some lack adequate economic or institutional feasibility or information on limits (medium confidence). Feasible and effective options include cultivar improvements, community-based adaptation, agricultural diversification, climate services, adaptive eco-management in fisheries and aquaculture. There is limited evidence, medium agreement on the institutional feasibility or cost effectiveness of adaptation activities, and the limits to such adaptations. {5.4.4, 5.5.4, 5.6.3, 5.8.4, 5.9.4, 5.10.4, 5.11.4, 5.12.4, 5.14.1}
Ecosystem-based approaches such as diversification, land restoration, agroecology and agroforestry have the potential to strengthen resilience to climate change with multiple co-benefits, but trade-offs and benefits vary with socio-ecological context (high confidence). Ecosystem-based approaches support long-term productivity and ecosystem services such as pest control, soil health, pollination and buffering of temperature extremes (high confidence), but potential and trade-offs vary by socioeconomic context, ecosystem zone, species combinations and institutional support (medium confidence). {5.4.4.4, 5.6.3, 5.10.4, 5.14.1, Cross-Chapter Box NATURAL in Chapter 2; Cross-Working Group Box BIOECONOMY this chapter}
Bio-based products as part of a circular bioeconomy have potential to support adaptation and mitigation, with sectoral integration, transparent governance and stakeholder involvement key to maximising benefits and managing trade-offs (high confidence). A sustainable bioeconomy relying on bioresources will need to be supported by technology innovation and international cooperation and governance of global trade to disincentivise environmental and social externalities (medium confidence). {Cross-Working Group Box BIOECONOMY this chapter}
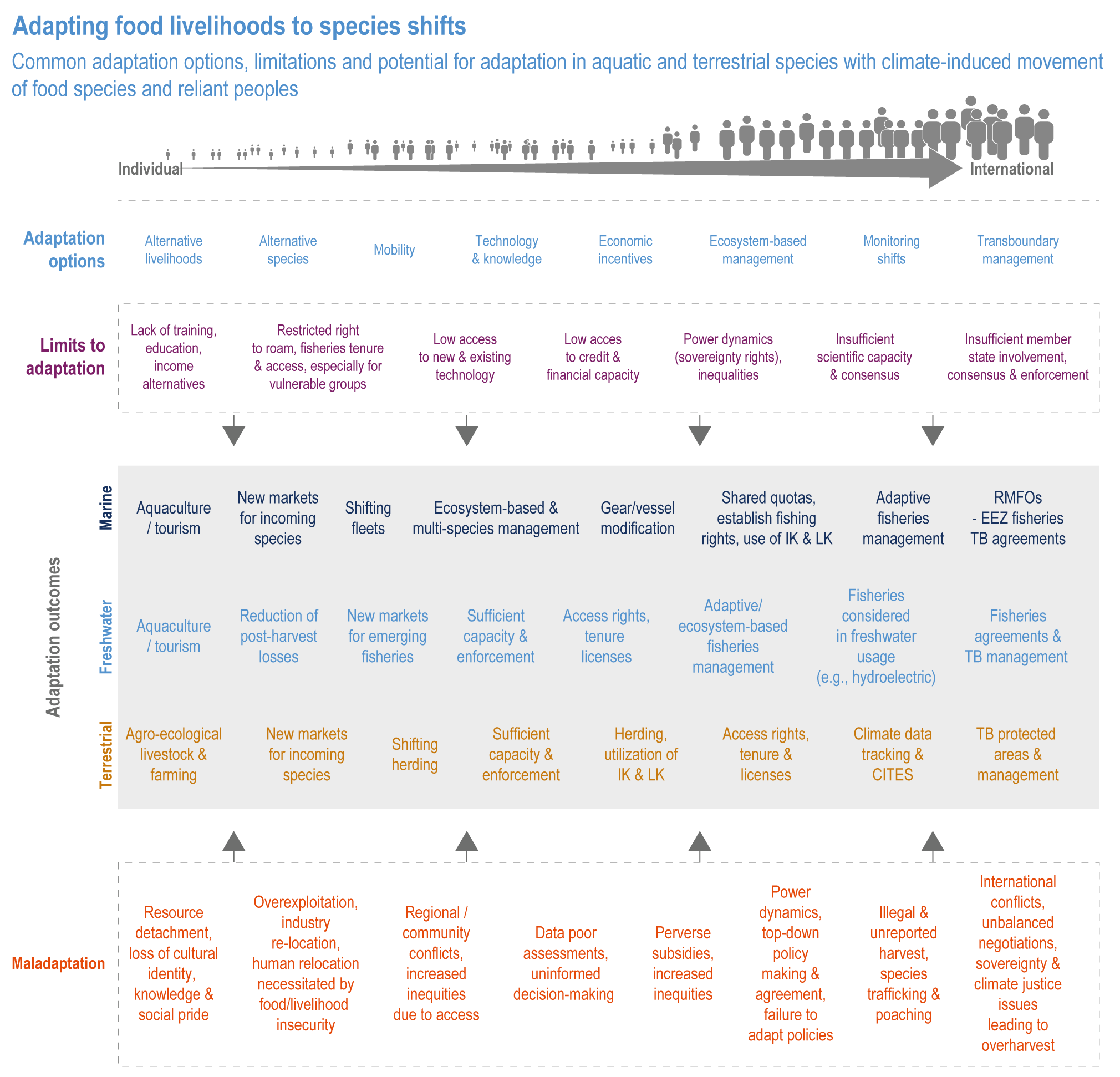
Sustainable resource management in response to distribution shifts of terrestrial and aquatic species under climate change is an effective adaptation option to reduce food and nutritional risk, conflict and loss of livelihood (medium confidence). Adaptive transboundary governance and ecosystem-based management, livelihood diversification, capacity development and improved knowledge-sharing will reduce conflict and promote the fair distribution of sustainably harvested wild products and revenues (medium confidence). Other options include shared quotas and access rights considering trade-offs, shifting livelihoods to follow target species, new markets for emerging species, and technology {Cross Chapter Box MOVING PLATE this chapter, 5.8.4, 5.14.3.4}
Implemented adaptation in crop production will be insufficient to offset the negative effects of climate change (high confidence). Currently available management options have the potential to compensate global crop production losses due to climate change up to ~2°C warming, but the negative impacts even with adaptation will grow substantially from the mid-century under high temperature change scenarios (high confidence). Regionally, the negative effects will prevail sooner where current temperatures are already higher as in lower latitudes (high confidence). {5.2.2, 5.4.3, 5.4.4, 5.8.4, 5.9.4, 5.14.2.4}
Supportive public policies will enhance effectiveness and/or feasibility of adaptation in ecosystem provisioning services (medium confidence). Policies that support system transitions include shifting subsidies, removing perverse incentives, regulation and certification, green public procurement, investment in sustainable value chains, support for capacity-building, access to insurance premiums, payments for ecosystem services, and social protection, among others (medium confidence). {5.4.4.3; 5.4.4.4; 5.10.4.4; 5.12.6; 5.13.4; 5.14.1.3; 5.14.2.4; Box 5.13, Cross-Working Group Box BIOECONOMY in Chapter 2}
Harnessing youth innovation and vision alongside other SDGs such as gender equity, Indigenous knowledge, local knowledge, and urban and rural livelihoods, will support effective climate change adaptation to ensure resilient economies in food systems (high confidence). Adaptation strategies that address power inequities lead to co-benefits in equity outcomes and resilience for vulnerable groups (medium confidence). Indigenous knowledge and local knowledge facilitate adaptation strategies for ecosystem provisioning, especially when combined with scientific knowledge using participatory and community-based approaches (high confidence). {5.4.4.3, Table 5.6, 5.6.3, 5.8.4, 5.9.2, 5.9.4.1, 5.9.5, 5.10.2.2, 5.12.7, 5.12.8, 5.13.4, 5.13.5, 5.14.1.1, 5.14.1.2, 5.14.1.4,5.14.2.1, Box 5.13, 5.14.2.2 }
Policy decisions related to climate change adaptation and mitigation that ignore or worsen risks of adverse effects for different groups and ecosystems increase vulnerability, negatively affect capacity to deal with climate impacts, and impede sustainable development (medium confidence with robust evidence, medium agreement ) . Lacking sufficient stakeholder participation, large-scale land acquisitions have had mostly negative implications for vulnerable groups and climate change adaptation (high confidence). Policy and programme appraisal of adaptation options that consider the risks of adverse effects across different groups at different scales and use inclusive rights-based approaches help avoid maladaptation (medium confidence). Successful forest adaptation involves recognition of land rights and cooperation with Indigenous Peoples and other local communities who depend on forest resources (high confidence). {5.6.3; 5.12.3, 5.13.1; 5.13.2; 5.14.2.1}
Financial barriers limit implementation of adaptation options in agriculture, fisheries, aquaculture and forestry, and vastly more public and private investment is required (high confidence) . Public-sector investment in adaptation of agriculture, forestry and fisheries has grown four-fold since 2010, but adaptation costs will be much higher to meet future adaptation needs (medium confidence). Expanding access to financial services and pooling climate risks will enable and incentivise climate change adaptation (medium confidence). {5.14.3, 5.14.5., Cross-Chapter Box FINANCE in Chapter 17}
Climate resilient development pathways offer a way forward to guide climate action in food system transitions, but operationalisation is hampered by limited indicators and analyses (medium confidence). Robust analyses are needed that detail plausible pathways to move towards more resilient, equitable and sustainable food systems in ways that are socially, economically and environmentally acceptable through time (high confidence). Appropriate monitoring and rapid feedback to food system actors will be critical to the success of many current and future adaptation actions (high confidence). {5.14.4}
5.1 Introduction
5.1.1 Scope of the Chapter
This chapter assesses the scientific literature produced after the Intergovernmental Panel on Climate Change (IPCC) 5th Assessment Report (AR5) dealing with past, current and future climate change effects on managed ecosystems that provide provisioning and cultural services. It spans low- and high-intensity production systems for food, feed, fibre and other ecosystem products.
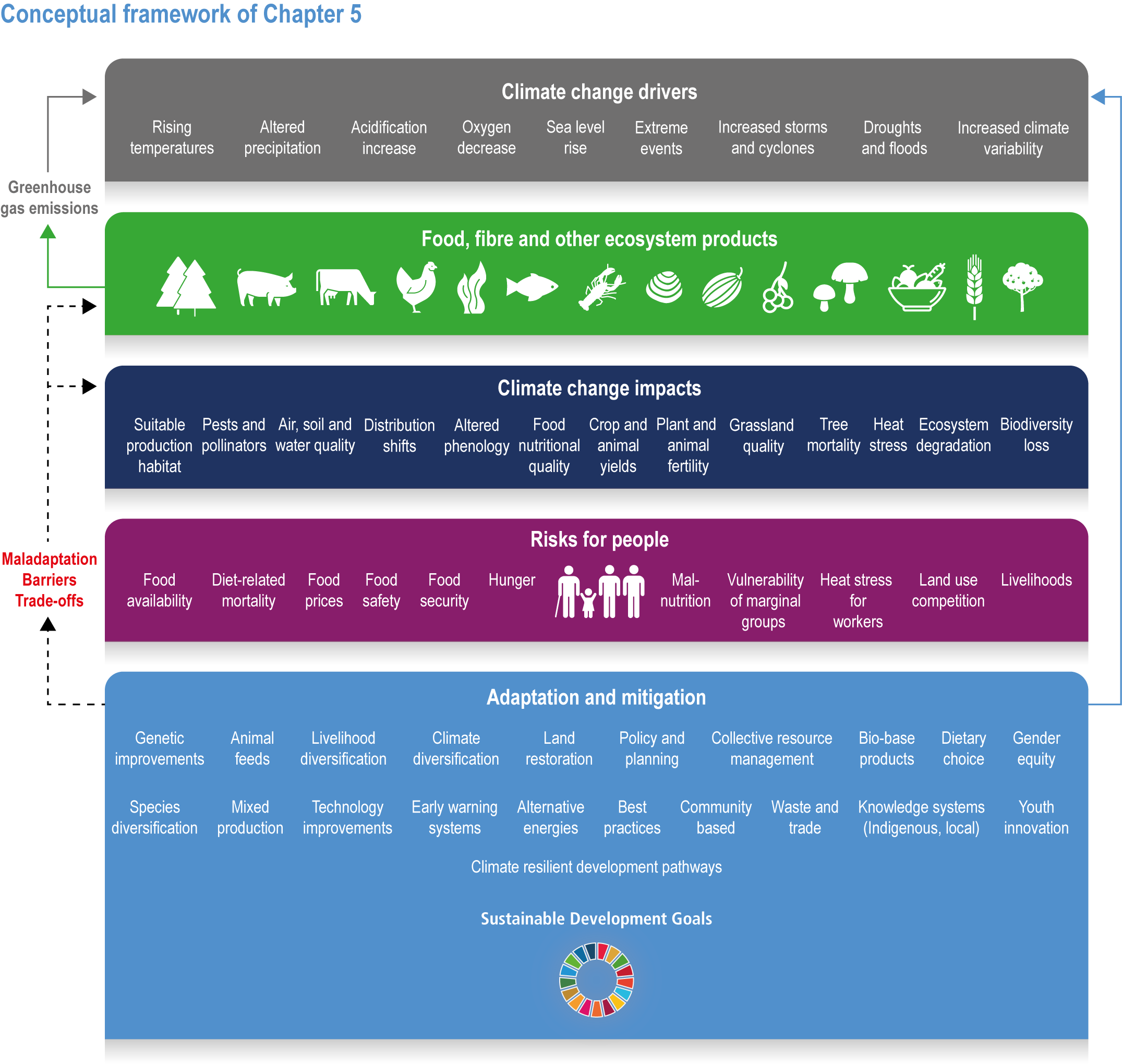
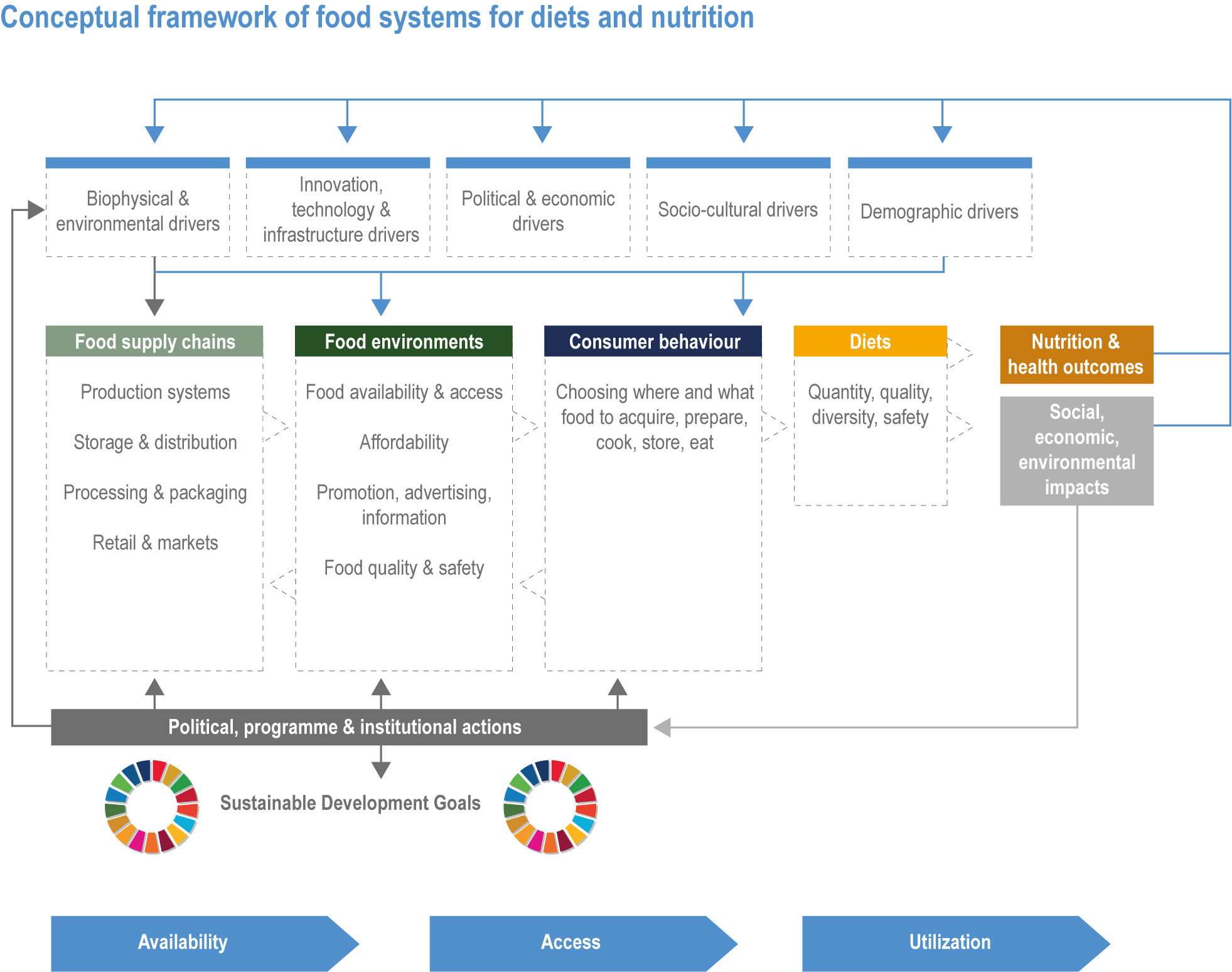
Climate change has already had global impacts, including in high-income countries. Special emphasis is placed on the assessment of vulnerabilities of particular groups that are context- and location-specific, such as Indigenous Peoples and other minorities, women and small-scale food producers. The report builds on the IPCC AR5 and recent Special Reports. This chapter combines food systems, fibre, wood and other products from ecosystems previously detailed in separate chapters of AR5, with an increased focus on ecosystem services, including the long-term sustainability of the global food system (Figure 5.1). The chapter focuses on key climate risks, implementation and outcomes of adaptation solutions for different groups as well as limits to adaptation.
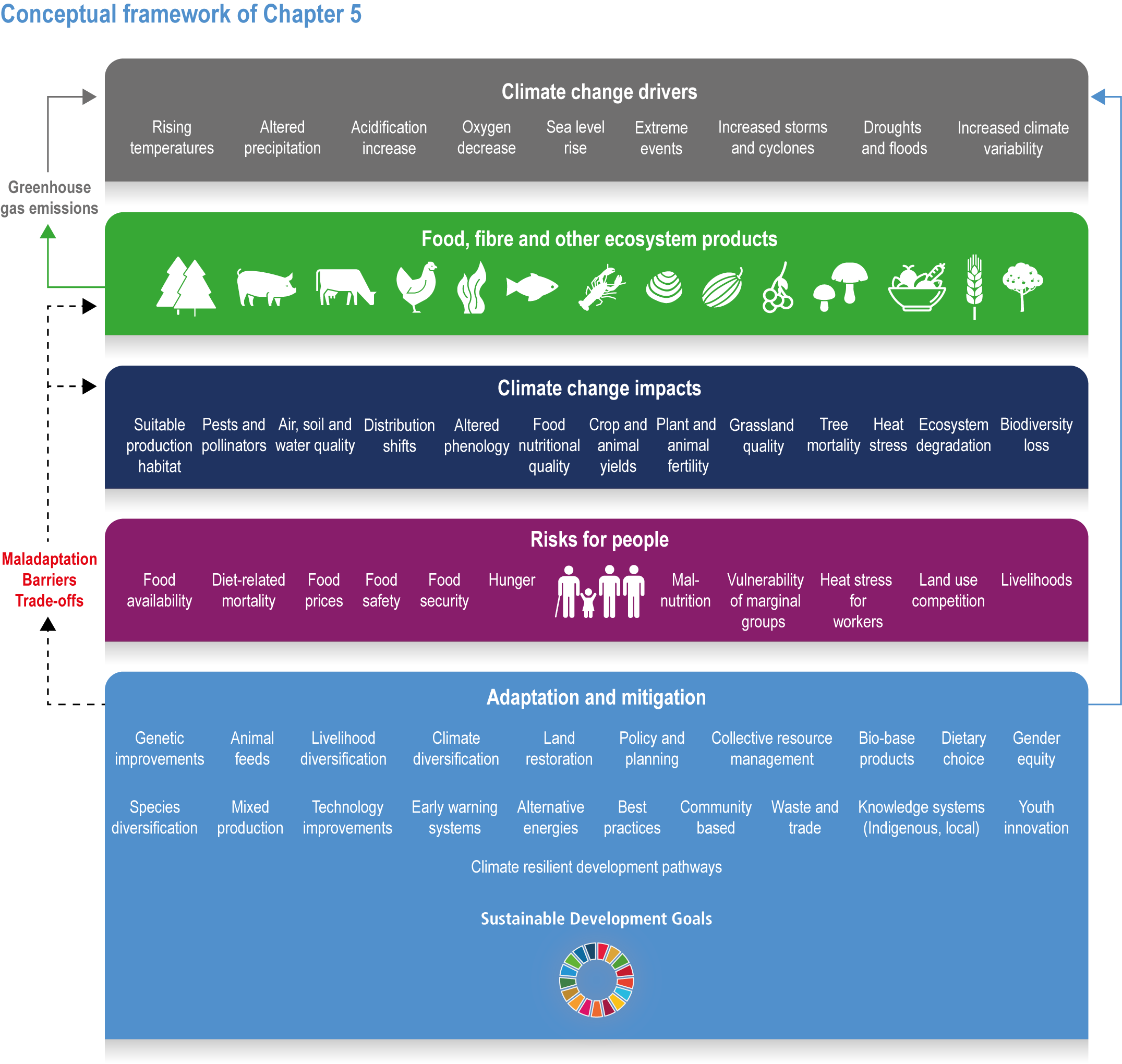
Figure 5.1 | Conceptual framework of Chapter 5.
5.1.2 Starting Point: AR5 and Recent IPCC Special Reports
AR5 Chapter 7 (Porter et al., 2014) reported with high confidence that food production systems were being negatively impacted by climate change, including both terrestrial and aquatic food species (Porter et al., 2014). Increased temperatures will have large negative impacts on the food production system under 2°C warming by late 20th century, with temperatures exceeding 4°C posing even greater risk to global food security (Porter et al., 2014). Adaptation options are needed to reduce the risk from climate change, but there was limited information of their effectiveness.
The 1.5°C Special Report concluded that climate-related risks to food security will rise under 1.5°C and will increase further under 2°C or higher. Above 1.5°C, currently available adaptation options will be much less effective and site-specific limits to adaptation will be reached for vulnerable regions and sectors. There was high confidence that limiting warming to 1.5°C will result in smaller net reductions in yields of major crops affecting food availability and nutrition, and that rising temperatures will adversely affect livestock via changes in feed quality, fertility, production, spread of diseases and water availability.
The IPCC Special Report on Climate Change and Land (SRCCL) expanded beyond the 1.5°C report to provide more in-depth information on climate change interactions with food security, desertification and degradation. There was high confidence that climate risks, both for slow changes and extreme events, are interlinked with ecosystem services, health and food security, often cascading and potentially reinforcing effects. Climate change already affects all dimensions of food security, namely availability, access, utilisation and stability, by disrupting food production, quality, storage, transport and retail. These effects exacerbate competition for land and water resources, leading to increased deforestation, biodiversity reduction and loss of wetlands. With high certainty, limiting global warming would lower future risks related to land, such as water scarcity, fire, vegetation shifts, degradation, desertification and food insecurity and malnutrition, particularly for those most vulnerable today: small-scale food producers in low-income countries, Indigenous communities, women, and the urban poor. SRCCL assessed a range of adaptation pathways to increase food resilience.
The IPCC Special Report on the Ocean and Cryosphere in a Changing Climate (SROCC) identified climate change impacts of warming, deoxygenation and acidification of the ocean and reductions in snow, sea ice and glaciers as having major negative impacts on fisheries and crops watered from mountain runoff and agriculture. These impacts affect food provisioning of food and directly threaten livelihoods and food security of vulnerable coastal communities and glacier-fed river basins. Climate change impacts on fisheries will be particularly high in tropical regions, where reductions in catch are expected to be among the largest globally, leading to negative economic and social effects for fishing communities and with implications for the supply of fish and shellfish (high confidence). While specific impacts will depend on the level of global warming and mitigative action to improve fisheries and aquaculture management, some current management practices and extraction levels may not be viable in the future.
5.1.3 Chapter Framework
This chapter is taking a food systems approach similar to the food security chapter in SRCCL (Mbow et al., 2019), with close attention to food system linkages, interactions and impacts on ecosystem services and biodiversity (Steffen et al., 2015; Raworth, 2017; Gerten et al., 2020). Food security is defined as “a situation that exists when all people, at all times, have physical, social and economic access to sufficient, safe and nutritious food that meets their dietary needs and food preferences for an active and healthy life” (FAO, 2020). Food insecurity is often experienced as chronic hunger reported in the annual UN Food Security and Nutrition in the World (FAO, 2020), when a person is unable to consume enough food over an extended period. The chapter gives special attention to climate change impacts on acute food insecurity, which can occur at any time with a severity that threatens lives, livelihoods or both, regardless of the causes, context or duration, as a result of shocks risking determinants of food security and nutrition, and used to assess the need for humanitarian action (IPC Global Partners, 2019).
Climate change directly affects food systems, and the impacts on terrestrial or aquatic food production will become increasingly negative, although regionally some changes may be beneficial in the near future (Porter et al., 2014). Current food system trajectories are leading to biodiversity loss and land and aquatic ecosystem degradation without delivering food security, nutrition, and sustainable and healthy livelihoods to many (Steffen et al., 2015). Addressing climate change in isolation ignores these interconnections, which is why the chapter considers integrated adaptation solutions to allow humanity to thrive in the long term. At the same time, social foundations of equality, justice and political participation are crucial in order to move towards a safe operating space for humanity (Raworth, 2017). The SDGs provide the most comprehensive set of metrics of humanity’s progress in achieving equitable and thriving socio-ecological systems. Therefore, while the focus of this chapter is climate change impacts, vulnerability and adaptation of food systems, feed, fibre and other ecosystem products, other environmental and social challenges are considered concomitantly.
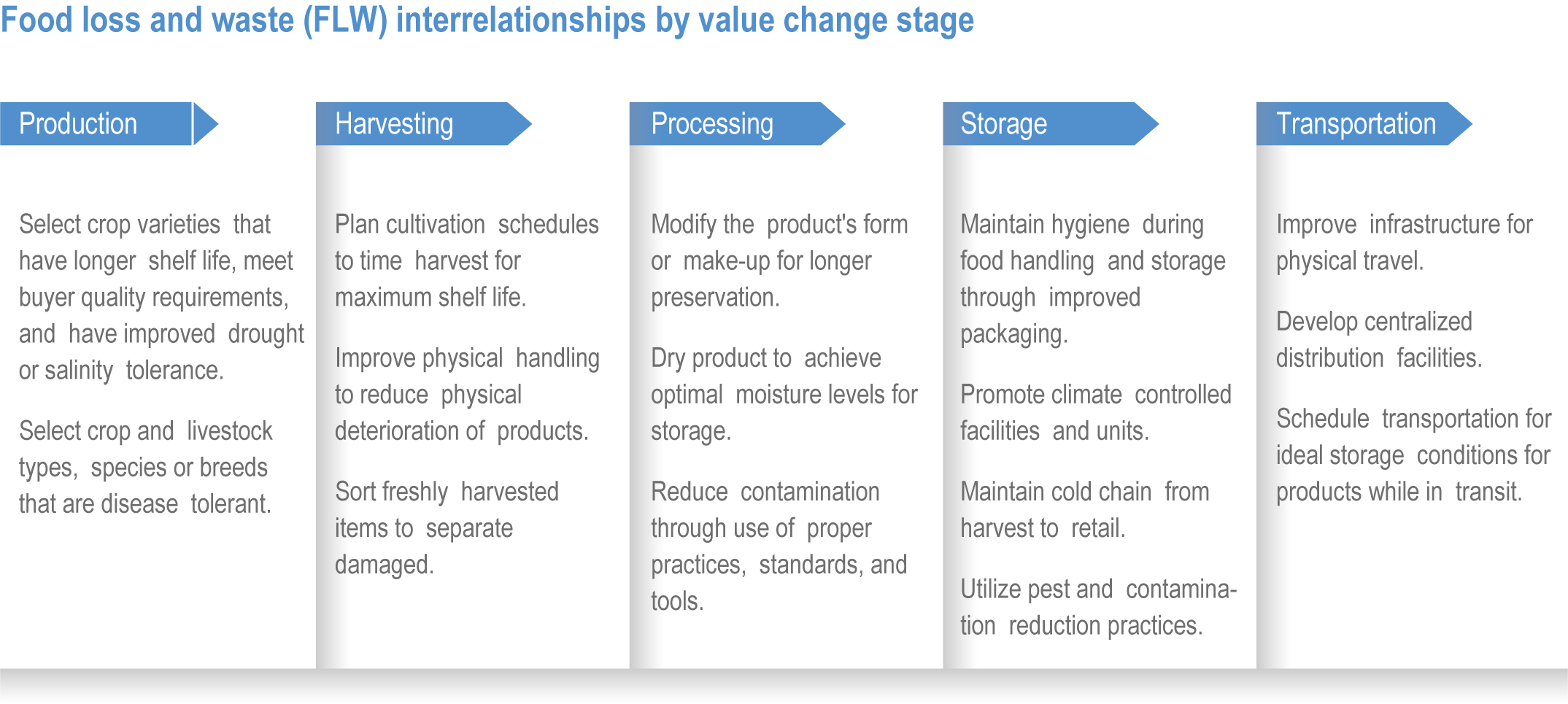
Food system and natural systems interact via political, economic, social, cultural and demographic factors in complex ways, leading to food security and sustainability outcomes. The food system has a supply (production) and demand (consumption) side, connected via processing, trade and retail, with loss and waste streams all along the food chain. Natural ecosystems provide multiple services (regulating, supporting, provisioning, cultural) to the food system. Food security and nutrition strongly depend on the driving forces connecting food and natural systems while at the same time positively or negatively influencing them. Climate change frequently exacerbates the effects of other drivers of change, further limiting the environment within which humanity can safely operate and thrive. The chapter assesses how climate change affects the four pillars of food security and nutrition and how these effects can be mediated by various factors, including our adaptation responses, social equity, underlying ecosystem services and governance (Figure 5.1). Adaptation solutions are a major emphasis of this chapter, including many ecosystem-based adaptation options (Table 5.1), which fall under the broader umbrella of nature-based solutions (Seddon et al., 2020).
Ecosystem-based adaptation, defined as the ‘use of ecosystem management activities to increase the resilience and reduce the vulnerability of people and ecosystems to climate change’ (Campbell et al., 2009), has at its core the recognition that there are unexploited synergies in agricultural systems that can increase productivity and resilience. These can result from increasing biodiversity, adding organic matter to soils, integrating livestock and aquatic species, including aquaculture, into farming practices, broadening landscape practices to exploit crop–forestry synergies, supporting beneficial insect populations and altering pest management practices that have unintended negative consequences. In addition, the chapter considers socioeconomic strategies to build resilience in the food system, strengthening local and regional economies, building on Indigenous and local knowledge, and addressing social inequity, through inclusive, participatory and democratic governance of food systems (HLPE, 2019; Wezel et al., 2020).
Table 5.1 | Adaptation strategies assessment in food, fibre and other ecosystem provisioning services.
Adaptation strategies/options | Systems | Benefits | Constraints or enablers | Confidence | Relevant sections |
| Crops |
| Secure tenure arrangements are often critical for delivering successful ecosystem-based adaptation | High | (5.4.4.5, 5.6.3, 5.12.3, Cross-Chapter Box NATURAL in Chapter 2, 5.14.3.6, 5.14.3.11; Cross-Chapter Box HEALTH in Chapter 7) |
| Crops, livestock, aquaculture, mixed, agroforestry systems |
| Policies and technologies that support diversification at landscape and farm levels: programmes that reward farmers for diversification practices, reduced incentives for intensified monocultures, extension support and market infrastructure for diverse crops, and productivity research on a greater variety of crops with support for post-harvest processing and regional markets | High | (5.4.4.4, 5.14.3.1, 5.14.3.6) |
| Crops–livestock mixed system particularly in the tropics and subtropics |
| Gender inequalities can act as a risk multiplier | Medium | (5.5.4; 5.10.4) |
| Crops, forestry, fisheries |
| Indigenous knowledge and local knowledge can facilitate adaptation when combined with scientific knowledge and utilised in management regimes | High | (5.4.4.5, 5.6.3, 5.14.3) |
| Forestry |
| Partnerships between key stakeholders such as researchers, forest managers, and Indigenous and local forest dependent communities will facilitate sustainable forest management | Medium | (5.6.3) |
| Fisheries |
| Medium | (5.14.3.4; Cross-Chapter Box MOVING PLATE this chapter) | |
| Aquaculture |
| Governance that recognises unexploited biological and socioeconomic food system synergies and equity would lead to positive adaptation strategy development and implementation, but options may be limited for those most at risk due to technological cost and low financial access | High | (5.14.3.5) |
| Freshwater fisheries and aquaculture systems |
| Changing precipitation patterns will increase competition for limited freshwater supplies | Medium | (5.8.4, 5.9.4.) |
| Mixed system |
| Uncertainties exist concerning the scalability of integrated systems; their uptake faces particular barriers around risk, land tenure, social inclusion, information and management skill, and the nature and timing of benefit flows | High | (5.10.4) |
| Post-harvest |
| The extent to which adaptation activities beyond harvest are cost-effective, and the limits to such adaptation, are location-specific and largely unknown | Medium | (5.11.4) |
| Production and post-harvest |
| Differentiated responses based on food security level and climate risk can be effective | Medium | (5.12.4) |
| Production and post-harvest |
| Focus on meaningful participation in governance, design and implementation of adaptation strategies of those groups who are vulnerable, including gender. Can be conflicts and trade-offs, such as between addressing land rights or traditional fishing grounds | Medium | (5.12.4) |
| Production |
| For some high- and medium-income countries, evidence suggests that climate services have been underutilised. In low-income countries, use of climate services can increase yields and incomes and promote changes in farmers’ practices, but low confidence that climate services are delivering on their potential, whether they are being accessed by the vulnerable, and how these services are contributing to food security and nutrition | Medium | (5.14.1) |
5.2 Observed Impacts and Key Risks
5.2.1 Detection and Attribution of Observed Impacts
Detection and attribution of climate change impacts on the food system remain challenging because many non-climate drivers are involved (Porter et al., 2014) but have been improved by recently developed climate model outputs tailored for impact attribution (Iizumi et al., 2018; Moore, 2020; Ortiz-Bobea et al., 2021).
Climate change has caused regionally different, but mostly negative, impacts on crop yields and quality and marketability of products (high confidence) (see Section 5.4.1 for observed impacts). There is medium evidence and high agreement that the effects of human-induced climate warming since the pre-industrial era has had significantly negative effects on global crop production, acting as a drag on the growth of agricultural production (Iizumi et al., 2018; Moore, 2020; Ortiz-Bobea et al., 2021). One global study using an empirical model estimated the negative effect of anthropogenic warming trends from 1961 to 2017 to be on average 5.3% for three staple crops (5.9% for maize, 4.9% for wheat and 4.2% for rice) (Moore, 2020). Another study using a process-based crop model found a yield loss of 4.1% (0.5–8.4%) for maize and 4.5% (0.5–8.4%) for soybean between 1981 and 2010 relative to the non-warming condition, even with CO2 fertilisation effects (Iizumi et al., 2018). Human-induced warming trends since 1961 have also slowed down the growth of agricultural total factor productivity by 21% (Ortiz-Bobea et al., 2021). Regionally, heat and rainfall extremes intensified by human-induced warming in West Africa have reduced millet and sorghum yields by 10–20%, and 5–15%, respectively (Sultan et al., 2019).
Methane emissions significantly impact crop yields by increasing temperatures as a greenhouse gas (GHG) and surface ozone concentrations as a precursor (medium confidence) (Shindell, 2016; Van Dingenen, 2018; Shindell et al., 2019). Shindell (2016) estimated a net yield loss of 9.5±3.0% for four major crops due to anthropogenic emissions (1850–2010), after incorporation of the positive effect of CO2 (6.5±1.0%) and the negative effects of warming (10.9±3.2%) and tropospheric ozone elevation (5.0±1.5%). Although these estimates were not linked with historical yield changes, more than half of the estimated yield loss is attributable to increasing temperature and ozone concentrations from methane emissions, suggesting the importance of methane mitigation in alleviating yield losses (medium confidence) (Section 5.4.1.4).
Climate change is already affecting livestock production (high confidence) (Section 5.5.1). The effects include direct impacts of heat stress on mortality and productivity, and indirect impacts have been observed on grassland quality, shifts in species distribution and range changes in livestock diseases (Sections 5.5.1.1–5.5.1.3). Quantitative assessment of observed impacts is still limited.
In aquatic systems, more evidence has accumulated since AR5 on warming-induced shifts (mainly poleward) of species (high confidence) (Section 5.8.1, Cross-Chapter Box MOVING PLATE this chapter), causing significant challenges for resource allocation between different countries and fishing fleets. Quantitative assessments of climate change impacts on production are still limited, but Free et al. (2019) estimated a 4.1% global loss of the maximum sustainable yield of several marine fish populations from 1930 to 2010 due to climate change. The effects of climate change on aquaculture are apparent but diverse, depending on the types and species of aquaculture (high confidence) (Section 5.9.1). Temperature increases, acidification, salt intrusion, oxygen deficiency, floods and droughts have negatively impacted production via reduced growing suitability, mortalities or damages to infrastructure (Section 5.9.1).
The impacts of climate change on food provisioning have cascading effects on key elements of food security, such as food prices, household income, food safety and nutrition of vulnerable groups (Peri, 2017; Ubilava, 2018; 5.11, 5.12). Climate extreme events are frequently causing acute food insecurity (Section 5.12.3, FSIN, 2021). There is growing evidence that human-induced climate warming has amplified climate extreme events (Seneviratne et al., 2021), but detection and attribution of food insecurity to anthropogenic climate change is still limited by a lack of long-term data and complexity of food systems (Phalkey et al., 2015; Cooper et al., 2019). A recent event attribution study by Funk (2018) demonstrated that anthropogenic enhancement of the 2015/2016 El Niño increased drought-induced crop production losses in Southern Africa. Human-induced warming also exacerbated the 2007 drought in southern Africa, causing food shortages, price spikes and acute food insecurity in Lesotho (Verschuur et al., 2021).
5.2.2 Key Risks
Key risks in this chapter are grouped into those related to food security, food safety and dietary health, livelihoods of people in related sectors and ecosystem services (Table 16.9). Determining when a risk is considered severe is challenging to quantify because of the complexity of the food system, uncertainty about the effects and ethical challenges.
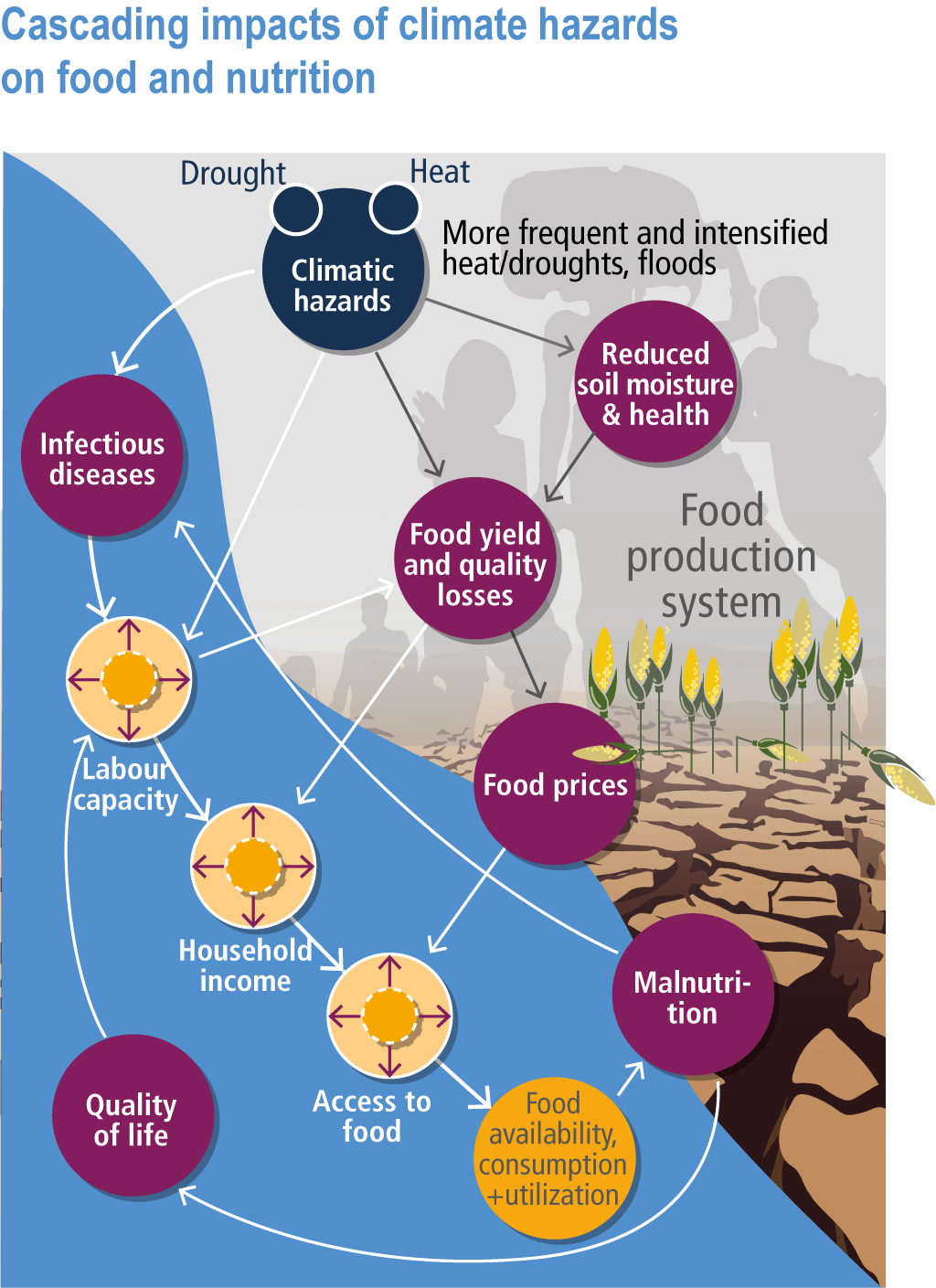
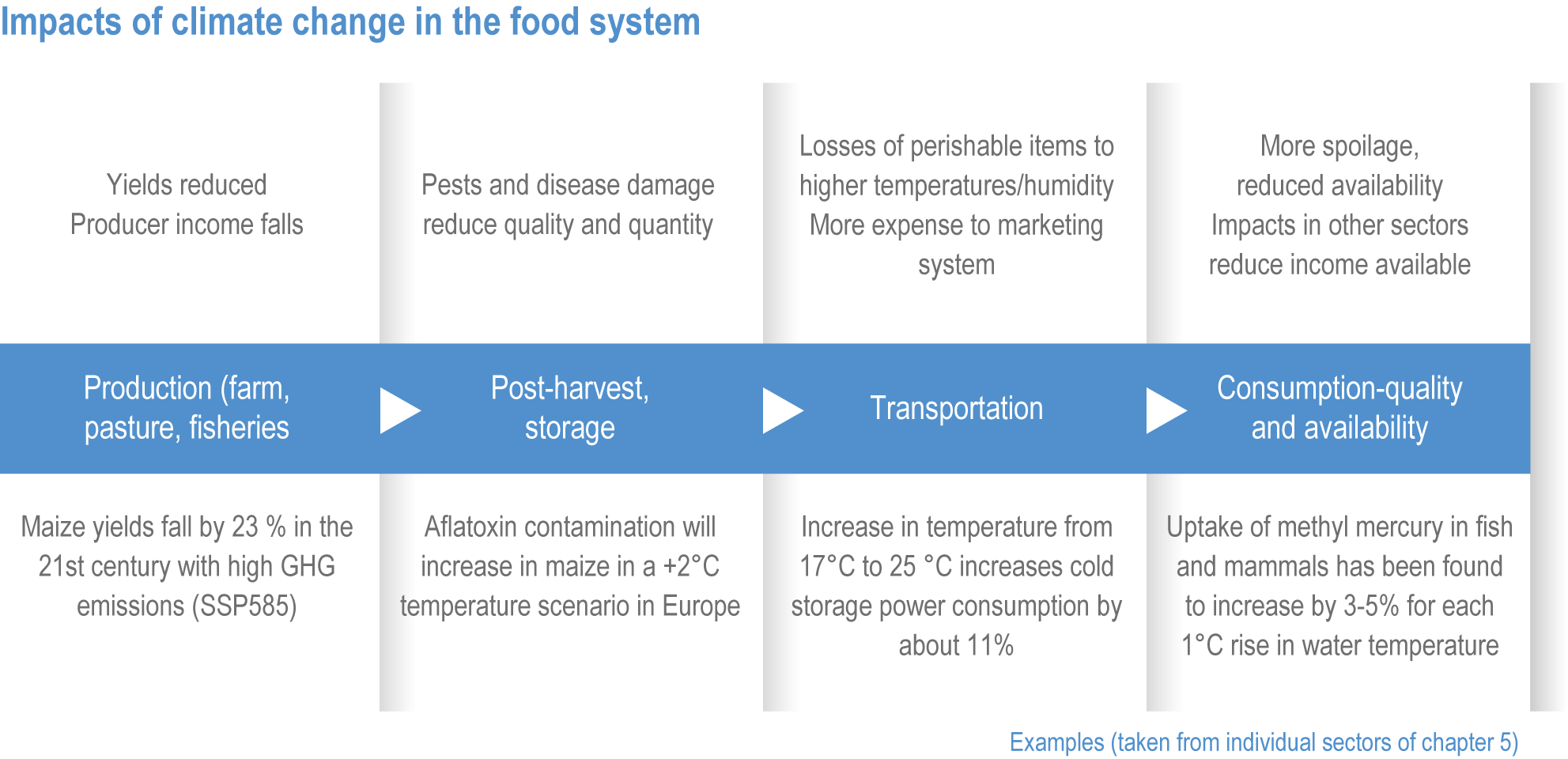
Current levels of food insecurity are already high in some parts of the world, and often exacerbated by short-term food shortages and price spikes caused by weather extremes partly linked to climate change (Sections 5.2.1, 5.12.3, 16.5.2). Climate change will increase malnourished populations through direct impacts on food production and have cascading impacts on food prices and household incomes, all of which will reduce access to safe and nutritious food (high confidence) (Figure 5.2, 5.12).
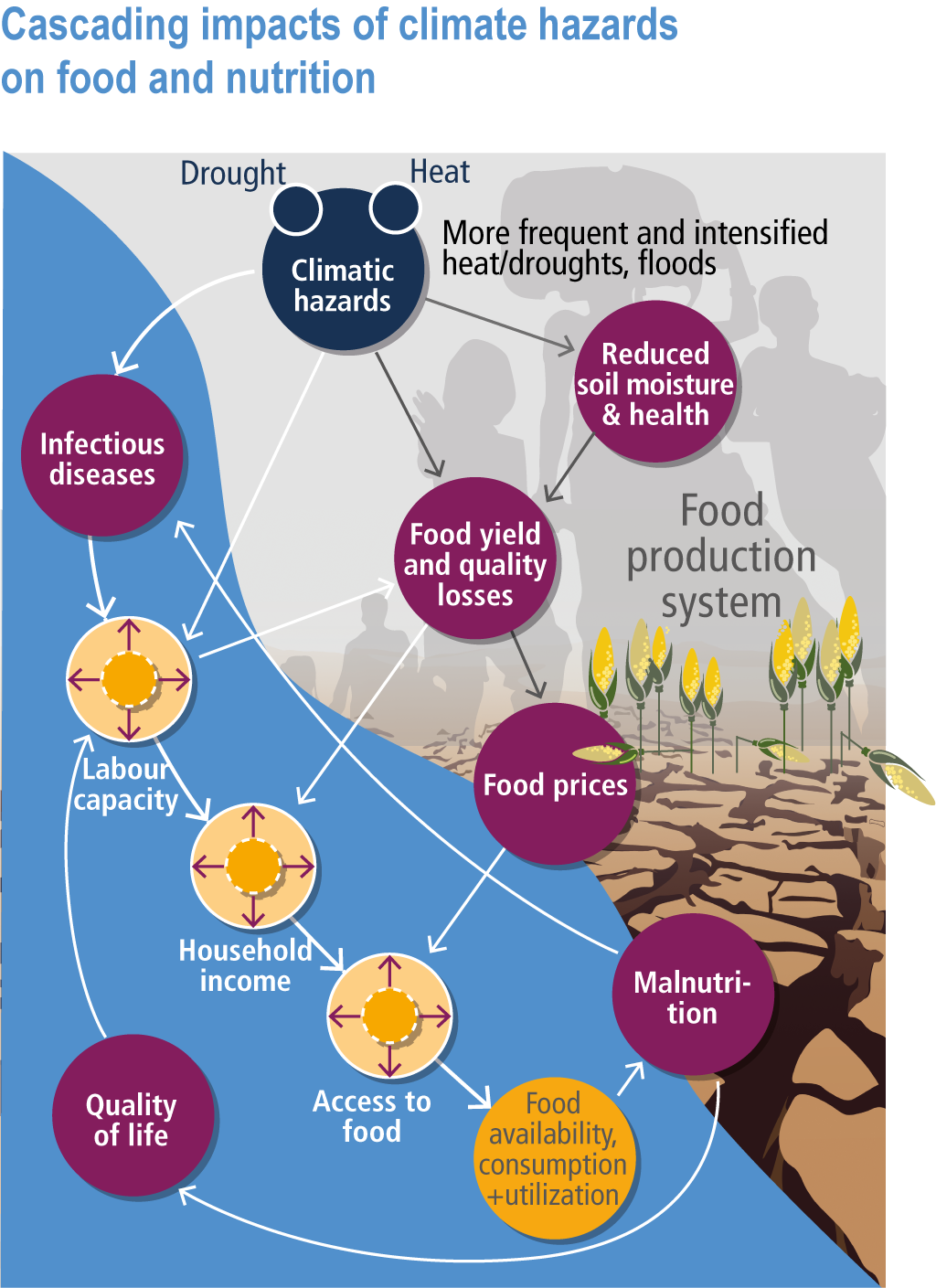
Figure 5.2 | Cascading impacts of climate hazards on food and nutrition. The factors involved the impacts on crop production and prices (black arrows) and interaction among food-health interaction (white arrows). Adapted and revised from (Phalkey et al., 2015).
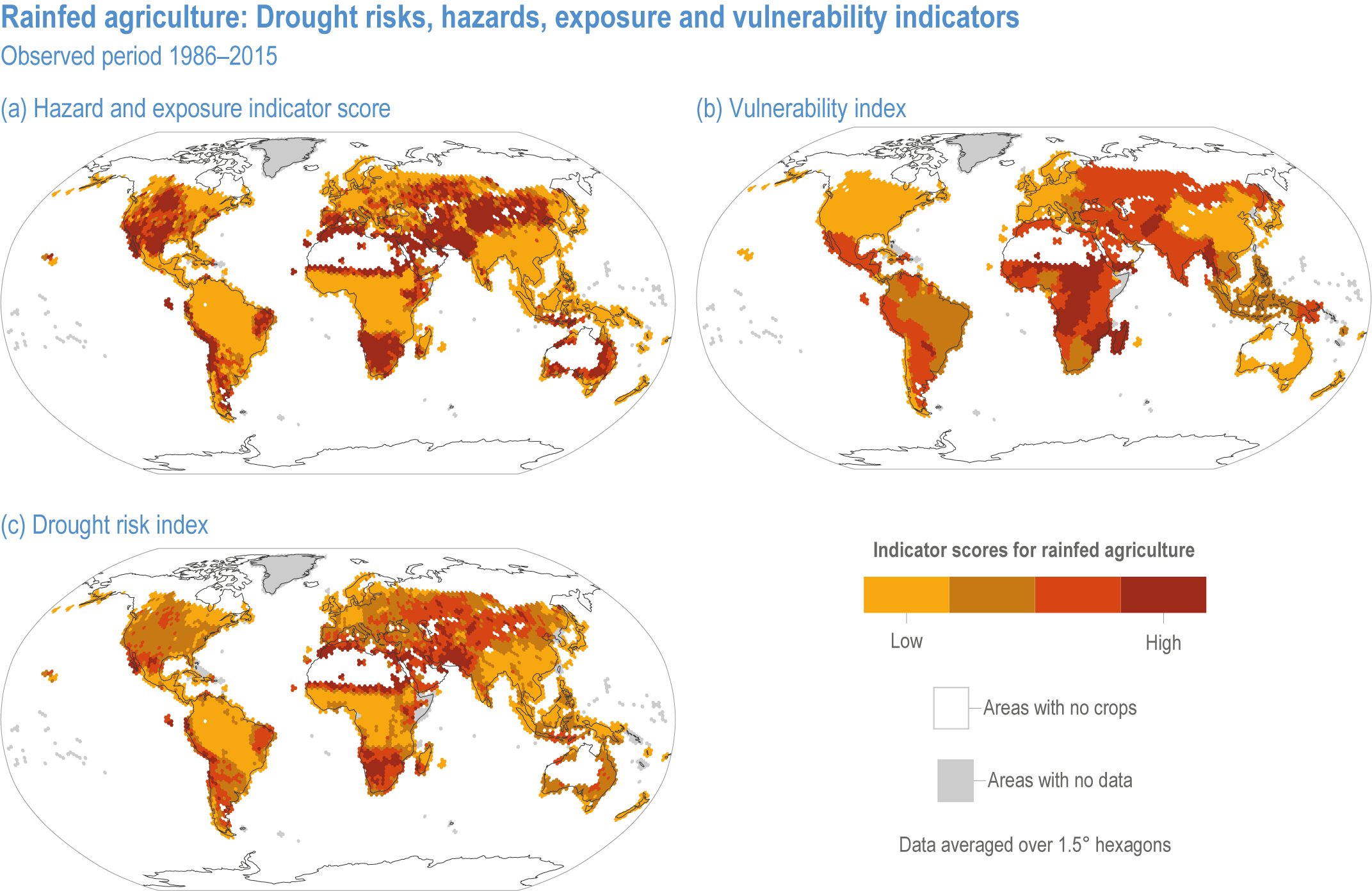
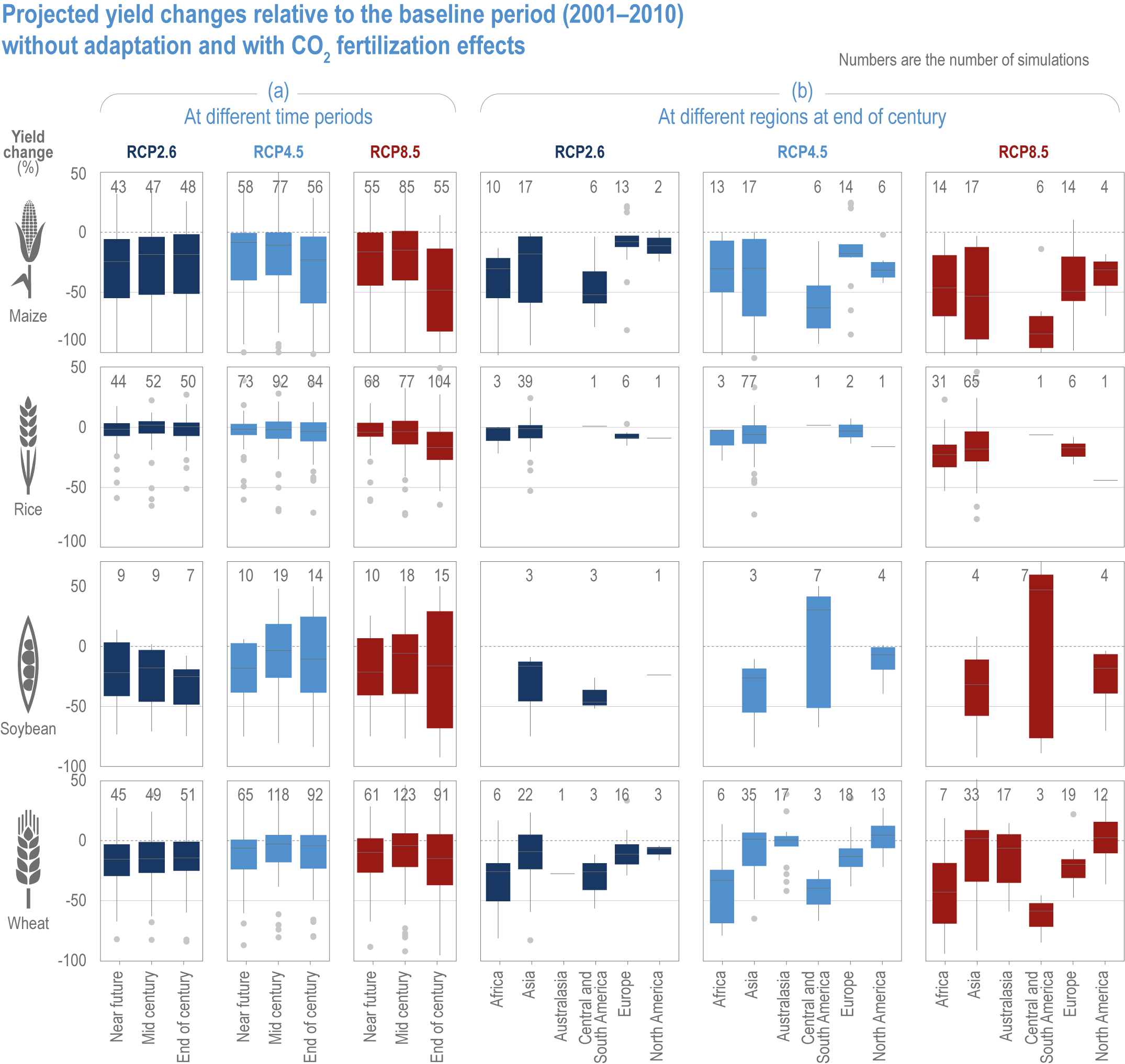
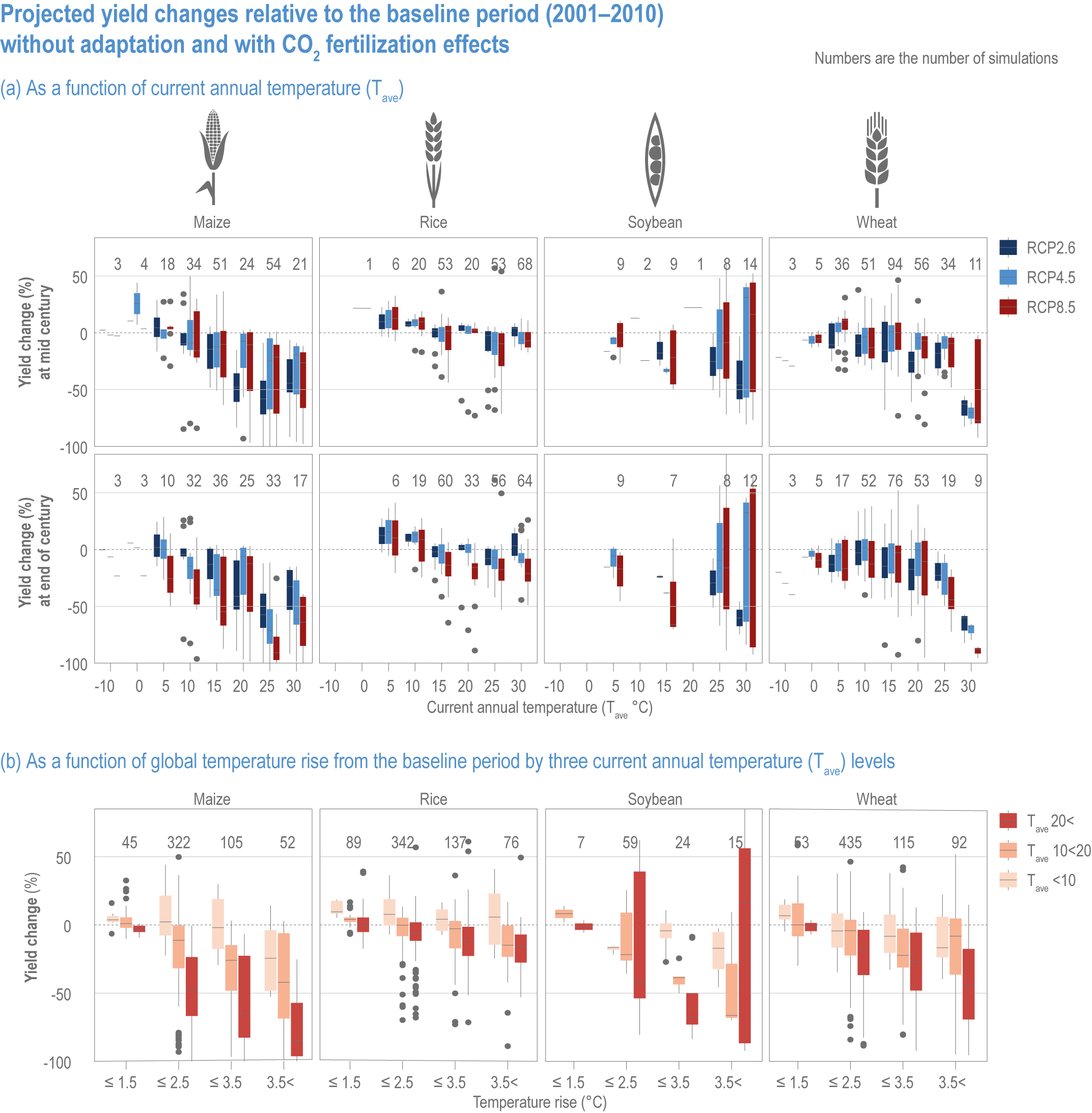
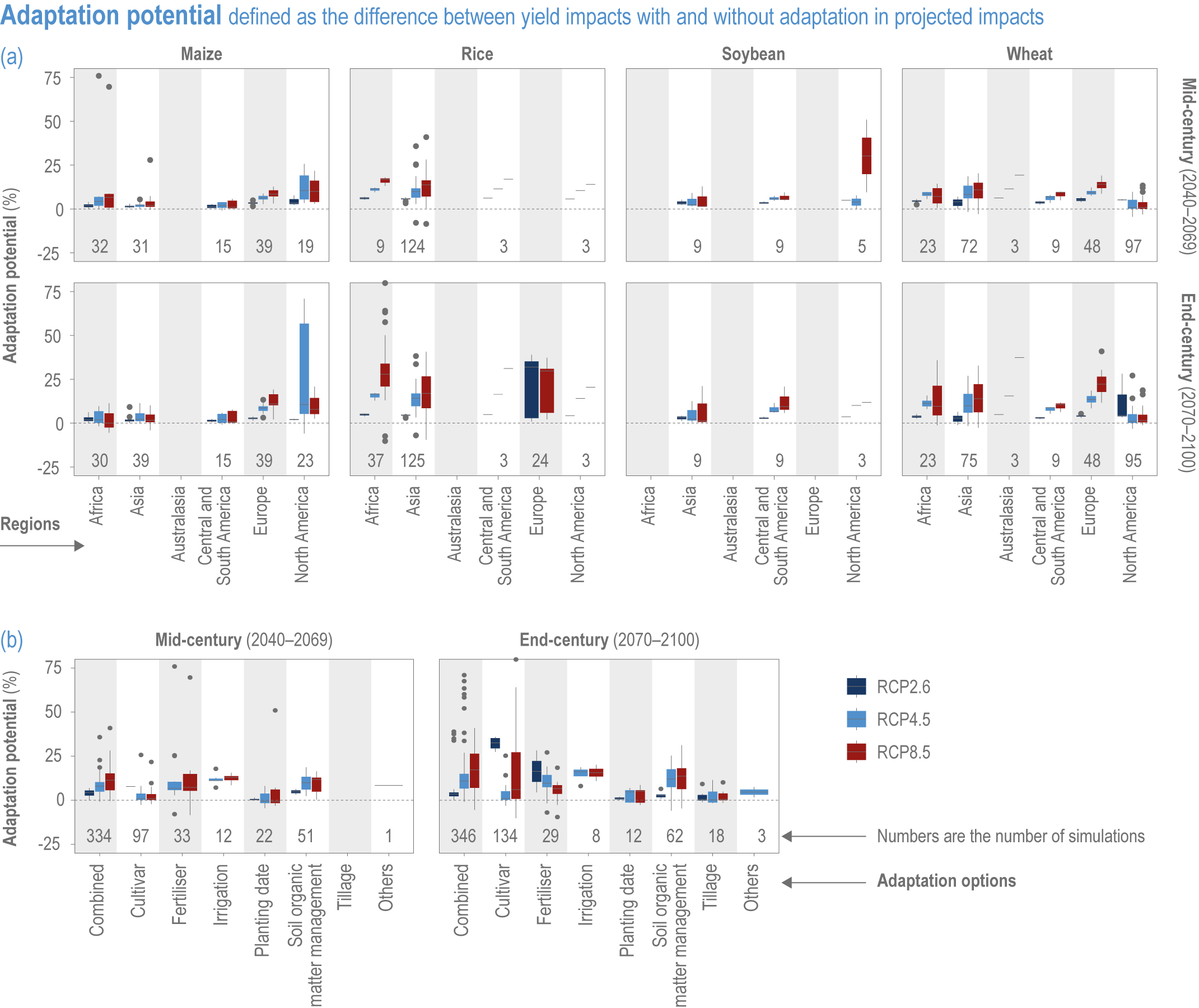
Extreme climate events will become more frequent and force some of the current food production areas beyond the safe climatic space for production (high confidence) (Sections 5.4.3, 5.5.2). Globally, 10% of the currently suitable area for major crops and livestock is projected to be climatically unsuitable in mid-century and 31–34% by the end of the century under SSP5-8.5 (Kummu et al., 2021). Adverse effects of climate change on food production will become more severe when global temperatures rise by more than 2°C (Sections 5.4.4.1, 5.12.4.1). One study estimated that the heat stress from projected 3°C warming above baseline (1986–2005) would reduce labour capacity by 30–50% in Sub-Saharan Africa and Southeast Asia, leading to a 5% increase in crop prices because of higher labour cost and production losses, thereby undermining food availability, access and livelihood (de Lima et al., 2021). Thiault et al. (2019) projected that, by 2100, climate change under RCP8.5 could have negative impacts on both agriculture and marine fisheries productivity in countries where 90% of the world population live. A global analysis of shellfish aquaculture estimated that habitat suitability will decline beyond 2060 globally, but much sooner in some Asian countries (Stewart-Sinclair et al., 2020; 5.9.1). These negative effects in the second half of the century will be much less under RCP2.6.
Climate change impacts will increase the number of people at risk of hunger, in 2050 ranging from 8 million people under SSP1 to 80 million people under SSP3 scenarios (RCP6.0), compared with a world with no climate change (Mbow et al., 2019). Estimates also vary depending on the adaptation and mitigation assumptions (Hasegawa et al., 2018; Janssens et al., 2020). Geographically, nearly 80% of the population at risk of hunger are projected to reside in Africa and Asia (Nelson et al., 2018). Projections of risk of hunger beyond 2050 are limited, but it will grow from the mid-century towards the end of the century, with more people at risk under RCP8.5 compared with RCP4.5 (Richardson et al., 2018). Regional disparity is projected to increase, particularly under a high-emission scenario.
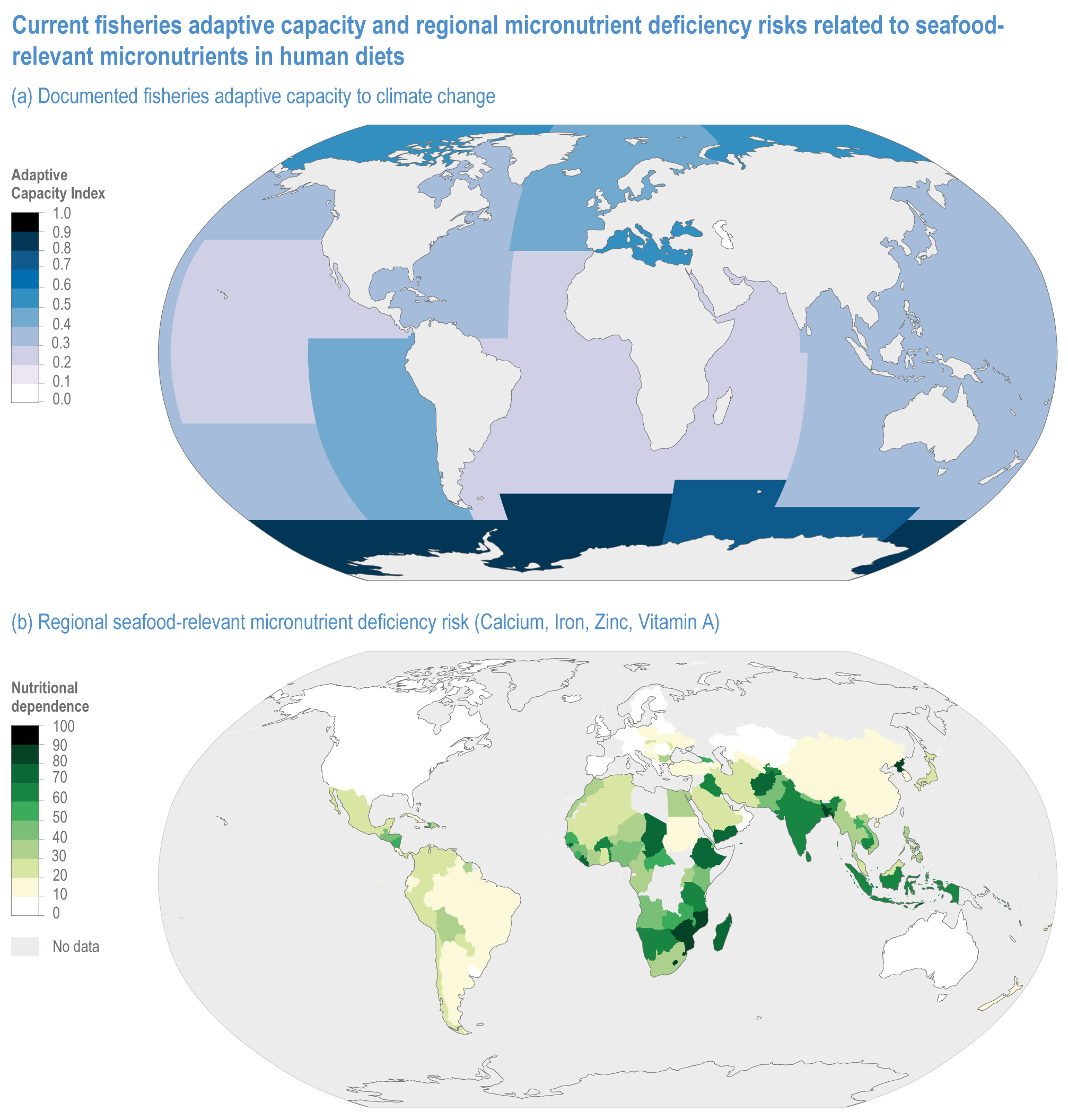
Climate change will increase the costs and management challenges of providing safe food. The safety challenges arise from contamination caused by increased prevalence of pathogens, HAB and toxic inorganic bioaccumulation (high confidence) (Sections 5.8, 5.9, 5.11, 5.12). Micronutrient deficiency is prevalent across many regions and will continue to be a problem at least during the first half of the century (Nelson et al., 2018), with significant implications for human health (Section 5.12.4).
Food security and healthy balanced diets will also be undermined by reduced livelihoods and health of people in agriculture and food-related sectors (Sections 5.12.3, 5.12.4), diminished ecosystem services provided by pollinators, the soil biome (Section 5.4.3) and water systems, and climate-mitigation related policies that solely focus on reducing GHG emissions without considering their potential to increase competition with food production for scarce land and water (Section 5.13.3).
5.3 Methodologies and Associated Uncertainties
Chapter text draws on previous IPCC reports, other reports (i.e., High Level Panel of Experts (HLPE), Food and Agriculture Organization (FAO), and Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES)), and literature published since 2014. This section highlights key trends in research topics and methods since AR5.
5.3.1 Methodologies for Assessing Impacts and Risks
Since AR5, there are more examples of observed impacts from past climate change in cropping systems (Section 5.4.1), pastoral systems (Section 5.5.1), forests (Section 5.6.1), fisheries (Section 5.8.1) and mixed farming systems (Section 5.10.1). These assessments of observed impacts make use of historical data on climate, production area and yield to attribute the role of climate in driving changes in suitability, production, yield, food quality or total factor productivity (Ortiz-Bobea et al., 2021). Observations across the global food systems have been analysed (Cottrell et al., 2019), with the advantage that unexpected impacts due to changes in seasonality and biotic interactions can be detected. Quantitative analysis is only possible in places with adequate historical data; in many cases, studies rely on qualitative assessments, often drawing on farmers’ perceptions of climate impacts.
Projecting future climate impacts relies on modelling that combines climate data with data from experimental studies testing how species respond to each climate factor. In cropping and forest systems, a network of experimental studies with plants exposed to elevated CO2 concentrations, ozone and elevated temperature provides data on the fundamental responses to climate and atmospheric conditions (i.e., free-air carbon dioxide enrichment (FACE) and temperature free-air controlled enhancement (T-FACE) systems). FACE results have been combined and assessed more extensively since AR5 (Bishop et al., 2014; Haworth et al., 2016; Kimball, 2016; Ainsworth and Long, 2021). Field-based FACE studies have several advantages over more enclosed testing chambers, although results from more controlled experiments and coordination between different methods continue to give new insights into crop responses to climate change and variability (Drag et al., 2020; Ainsworth and Long, 2021; Sun et al., 2021). Experimental results have limitations and can be difficult to scale up (Porter et al., 2014; Haworth et al., 2016), but generally the conclusions follow known plant responses (Lemonnier and Ainsworth, 2018). As highlighted in AR5, there is a scarcity of FACE infrastructure in the tropics and subtropics (Leakey et al., 2012; Lemonnier and Ainsworth, 2018; Toreti et al., 2020). One area that has been investigated further is the negative impact of elevated CO2 on crop nutritional value, which has important implications for human nutrition (Scheelbeek et al., 2018; Smith and Myers, 2018; Toreti et al., 2020; Ainsworth and Long, 2021). Increasingly, experimental studies seek to examine the interaction between climatic factors such as temperature, drought and ozone, or the responses of understudied food systems, crop species, cultivars and management interventions (Kimball, 2016; Ainsworth and Long, 2021). The use of experimental data to improve projections has also expanded in other systems. There has been an increased focus on the impact of warming on livestock health and productivity (5.5.3). Aquatic system studies have incorporated projected impacts on physiology, distribution, phenology and productivity (5.8.3).
Modelling approaches differ widely and serve different purposes (Table 5.2; Porter et al., 2014; Jones, 2017a). The use of process-based and statistical modelling alongside remote sensing and other spatial data has grown. Projections increasingly draw on a combination of modelling approaches and coordinated efforts for model intercomparisons and ensemble techniques, using standardised emission scenarios (RCPs). For major crops, models of global yield impacts from CO2 concentration, air temperature and precipitation have been refined and compared (Challinor et al., 2014; Iizumi et al., 2017; Ruane et al., 2017; Zhao et al., 2017; Rojas et al., 2019). Despite advances since AR5, modelling is still constrained by limited data from field experiments (Ruane et al., 2017). Increasingly, studies attempt to incorporate effects of elevated CO2, ozone and climate extremes (Barlow et al., 2015; Schauberger et al., 2019a; Vogel et al., 2019), as well as attempts to incorporate more complex interactions with soil and crop management (Basso et al., 2018; Smith et al., 2020b). However, only a few models consider crop protein content and other quality factors (Nuttall et al., 2017; Asseng et al., 2019). Some models take account of the impacts of climate on the timing of key biological events (phenology) in the target species; however, incorporating biotic interactions with pests, pathogens and pollinators remains a challenge (Table 5.2; Sections 5.4.1, 5.4.3).
Table 5.2 | A comparison of modelling approaches and their application in climate change impact projections. Model types are categorised by: food system, with labels representing the food systems from this chapter where each model type is used ({CROP}, {TREE}, {LIVES}, {FISH}, {MIX}, {FOOD}); scale over which each model type is usually applied local [()], regional [( )], global [( )], or a combination of these); and sensitivity to climate change where the colour intensity indicates the ability of each model type to incorporate each of the listed factors. After Van Wijk et al. (2014), Kanter et al. (2018) and Thornton (2018). Integrated assessment models are discussed in the main text.
Description | Applications for each food-system | Scale | Sensitivity to climate change factors and responses | ||||||
Climate | CO2 | Biotic | Adaptation | System responses | |||||
Empirical | Agroclimatic indices | Use simple equations to link agricultural performance to key climate factors, such as drought or heat stress, or summarise agricultural requirements using multiple environmental descriptors. | Comparing regions; matching crops to regions; early warning systems: e.g Agro-ecological zones, Ecocrop, Palmer Drought Severity Index {CROP}. | (( )) | |||||
Statistical models | Use quantitative associations between agricultural performance and climate, based on past observations. Can include projections for biotic factors such as pest and disease. | Productivity and production area projections; annual climate variability; attribution: e.g. Traditional: regression, statistical emulators {CROP}{TREE}{LIVES}{FISH}; e.g. Spatial suitability models /niche models: MaxEnt, CLIMEX, Ecocrop {CROP}{TREE}{FISH}. | (( )) | ||||||
Process-based (dynamic simulation models) | Vegetation focussed | Use combinations of land-surface energy and soil water balance models to simulate the growth of crop species along with natural vegetation, typically using plant and crop functional types. | Productivity projections; interactions with non-climate variables (e.g. CO2): e.g. PEGASUS, Agro-IBIS, DayCent, LPJmL, LPJ-GUESS, ORCHIDEE{CROP}{TREE}. | ( ) | |||||
Species focussed | Use mechanistic models based on the known responses of species to key environmental descriptors over time. Typically based on detailed information for a particular species within a region, but also applied to mixed systems such as agroforestry and globally. | Productivity projections; matching tree species to locations; species interactions; interactions with non-climate variable s (e.g.CO2); adaptation projections: e.g. point-based versions: APSIM, AquaCrop, DayCent, DSSAT, EPIC, Infocrop, SARRA-H, STICS{CROP}IBIS{TREE}LIVSIM, RUMINANT{LIVES}Fish-MIP{FISH}Yield-SAFE, WaNuLCAS, Hi-sAFe {MIX}; e.g. global gridded version: pDSSAT, pAPSIM, GEPIC, GLAM, MCWLA, PEGASUS, SARRA-O{CROP}. | (( )) | ||||||
Integrated Models | Optimization methods | Mathematical representations of systems with regard to key indicators, constraints, and objectives. Allows prioritisation of different climate change response options using the defined indicators. | Adaptation projections; food security projections; livelihood projections; trade-offs; live cycle assessment: e.g. Global Timber Model {TREE}CSAP toolkit, FarmDESIGN{CROP}{MIX}{FOOD} | () | |||||
Economic (Econometric, Economic surplus) | Used to integrate the broad impacts of climate change with other economic drivers, to quantify the economic costs and assess the value of adaptation/mitigation interventions. | Adaptation projections; food security projections; livelihood projections: e.g. GFPM{TREE}FUND 3.8, DICE 2010, IMPACT{FOOD} | ( ) | ||||||
Household and village models | Use detailed site-specific data to generate rules that describe the current behaviour of stakeholders such as households or villages. Can be integrated with other model approaches to consider climate response and adaptation interventions. | Adaptation projections (case specific); behavioural responses; trade-offs; participatory monitoring: e.g. DECUMA, PALM, MPMAS, MIDAS, TOA-MD{LIVES}{MIX}{FOOD} | (()) | ||||||
In addition to productivity projections, research also draws on climate suitability estimates (Table 5.2). These compare the known climate suitability of species and habitats with projected climate conditions across different locations. Such projections are useful especially for incorporating movement of pests and pathogens but cannot be applied in isolation if non-climate constraints are not considered. As different research groups use different assumptions and data inputs, more coordination is needed if suitability projections are to be compared globally (SM5.3).
Increasingly, projections look across different disciplines and across multiple components of the food system, including livestock, fisheries and mixed farming systems (Campbell et al., 2016; Mbow et al., 2019). Major timber species have been modelled, with projected impacts on productivity, duration of rotation and distribution (i.e., climate suitability) (Albert et al., 2018). Livestock systems are influenced by plant productivity projections via their feedstock, for example, rangeland cattle impacted by changes in net primary production (NPP) (Boone et al., 2018). Direct climate impacts on animals are also projected, using indices based on direct observations (Section 5.5.3). Since AR5, Fisheries and Marine Ecosystem Model Intercomparison Project (Fish-MIP) has allowed for global intercomparisons and ensemble projections of marine fisheries, and projections capturing interactions from multiple food systems (e.g., Inter-Sectoral Impact Model Intercomparison Project (ISI-MIP); Sections 5.8, 5.10).
Global simulations have uncovered important differences between regions (Deryng et al., 2016; Blanchard et al., 2017). Efforts to coordinate and combine regional and global modelling studies allow for greater insight into regional differences in climate change impacts, such as the Coordinated Global and Regional Assessments (CGRA) performed by the Agricultural Model Intercomparison and Improvement Project (AgMIP) (Blanchard et al., 2017; Müller et al., 2017; Rosenzweig et al., 2018; Ruane et al., 2018; Lotze et al., 2019). Increasingly, multi-model intercomparisons are used to evaluate global gridded crop models’ performance and sensitivity to temperature, water, nitrogen and CO2 within AgMIP, with the focus mostly on major annual crops (Valdivia et al., 2015; Ruane et al., 2017; Müller et al., 2021a). Differences in model type, structures and input data can result in large variation in projections, particularly for the response of crops to elevated CO2 and temperature (5.4.3.1); methods for quantifying and minimising this uncertainty have been developed, but improvement is still needed ( Asseng et al., 2015; Li et al., 2015; Zhao et al., 2017; Folberth et al., 2019; Tao et al., 2020; Müller et al., 2021a; Ruane et al., 2021). The use of multi-model intercomparisons has widened the range of uncertainties but has increased the robustness of impact assessments (Asseng et al., 2013; Challinor et al., 2014; Zhao et al., 2017). Model outputs are strongly influenced by decisions over which factors to include; for example, including drought impacts can result in positive yield projections switching to neutral or negative values (Gray et al., 2016; Jin et al., 2018). Models are also limited in their ability to incorporate socioeconomic drivers and extreme events (Porter et al., 2014; Campbell et al., 2016; Ruane et al., 2017; Jagermeyr and Frieler, 2018; Webber et al., 2018; Schewe et al., 2019).
For long-term projections and integrated assessments, a large component of uncertainty remains in the ability to represent socioeconomic responses to climate change and the degree to which these will mitigate or exacerbate climatic changes (Valdivia et al., 2015; Prestele et al., 2016; Arneth et al., 2019). This includes the potential adaptation responses of food producers. Models that incorporate alternative socioeconomic responses offer one solution (e.g., AgMIP) (Nelson et al., 2014; Von Lampe et al., 2014; Wiebe et al., 2015; Rosenzweig et al., 2018; van Zeist et al., 2020). Another approach is the use of solution-oriented scenarios to compare the effectiveness of adaptation options (Le Mouël and Forslund, 2017; Arneth et al., 2019), or to quantify the time period in which adaptation responses will become essential (Challinor et al., 2016; Rojas et al., 2019). Others point to the necessity of managing food systems within the context of uncertainty (Campbell et al., 2016).
5.3.2 Methodologies for Assessing Vulnerabilities and Adaptation
Methods for monitoring vulnerability and adaptation are under-researched but have increased since AR5. Increasingly, projections move from individual crops to assessing risks across the food systems and the relative vulnerability of different systems (Campbell et al., 2016; Gil et al., 2017; Lipper et al., 2017; Richardson et al., 2018). Adaptation options can be considered as parameters in integrated models, such as those used in ISI-MIP, while others use systematic assessments of case studies, such as the application of agent-based household models to assessments of adaptation in livestock systems (Section 5.5.4). Quantitative studies are less common than qualitative assessments, and there is a need to combine modelling and qualitative approaches more effectively (Beveridge et al., 2018a; Vermeulen et al., 2018).
The food system is dynamic, with changes in management practices driven by many factors, including climate adaptation (Iizumi, 2019; Iizumi et al., 2021a). Adaptation potential, such as expected advances in crop breeding, are often not explicitly accounted for in modelling studies, but more recent studies do quantify the potential for adaptation (Iizumi et al., 2017; Tao et al., 2017; Aggarwal et al., 2019; Minoli et al., 2019). To account for this complexity, case studies rely on data derived from the perception and practices of stakeholders who are engaged in adaptation (usually autonomous adaptation) (Hussain et al., 2016; Lipper et al., 2017; Ankrah, 2018; Sousa-Silva et al., 2018). Case studies use a range of different indicators to monitor climate response options, making quantitative comparisons more difficult (Gil et al., 2017; Vermeulen et al., 2018). However, systematic comparisons have provided valuable insights (Descheemaeker et al., 2018; Shaffril et al., 2018; Aggarwal et al., 2019; Bene et al., 2019); for example, the sustainable livelihood framework has been applied widely to diverse aquatic systems (Bueno and Soto, 2017; Barange and Cochrane, 2018) and the Livelihood Vulnerability Index is well used across systems (Section 5.1 4). Coordinated efforts such as the AgMIP also provide systematic assessments (Blanchard et al., 2017; Lipper et al., 2017; Antle et al., 2018). Nonetheless, the full effectiveness of different adaptation options is difficult to assess given that many impacts have not yet occurred (due to the cumulative nature of impacts and the inertia in the climate system) (Stocker et al., 2013; Zickfeld et al., 2013).
Transformation of the food system that addresses all dimensions of ecosystem services is discussed in this chapter, including risk management and the communication of uncertainties (Section 5.1 4). The focus is on flexible approaches to risk and uncertainty, assessing trends, drivers and trade-offs under different future scenarios (Campbell et al., 2016).
5.4 Crop-Based Systems
Crops such as cereals, vegetables, fruit, roots, tubers, oilseeds and sugar account for about 80% of the dietary energy supply (FAO, 2019 f). Crops are a significant source of food and income for about 600 million farms in the world, 90% of which are family farms (Lowder et al., 2019). Previous assessment reports focused on yields of staple crops such as maize, wheat and rice, but studies are emerging on climate change impacts on other crops.
5.4.1 Observed Impacts
5.4.1.1 Observed impacts on major crops
AR5 Chapter 7 (Porter et al., 2014) stated with confidence that warmer temperatures have benefited agriculture in the high latitudes, and more evidence has been published to support this statement. Typical examples include pole-ward expansion of growing areas and reduction of cold stress in East Asia and North America (Table SM5.1).
Recent warming trends have generally shortened the life cycle of major crops (high confidence) (Zhang et al., 2014; Shen and Liu, 2015; Ahmed et al., 2018; Liu et al., 2018c; Tan et al., 2021). Some studies, however, observed prolonged crop growth duration despite the warming trends (Mueller et al., 2015; Tao et al., 2016; Butler et al., 2018; Zhu et al., 2018b) because of shifts in planting dates and/or adoption of longer-duration cultivars in mid-to-high latitudes. Conversely, in mid-to-low latitudes in Asia, a review study found that farmers favoured early maturing cultivars to reduce risks of damages due to drought, flood and/or heat (Shaffril et al., 2018), suggesting that region-specific adaptations are already occurring in different parts of the world (high confidence).
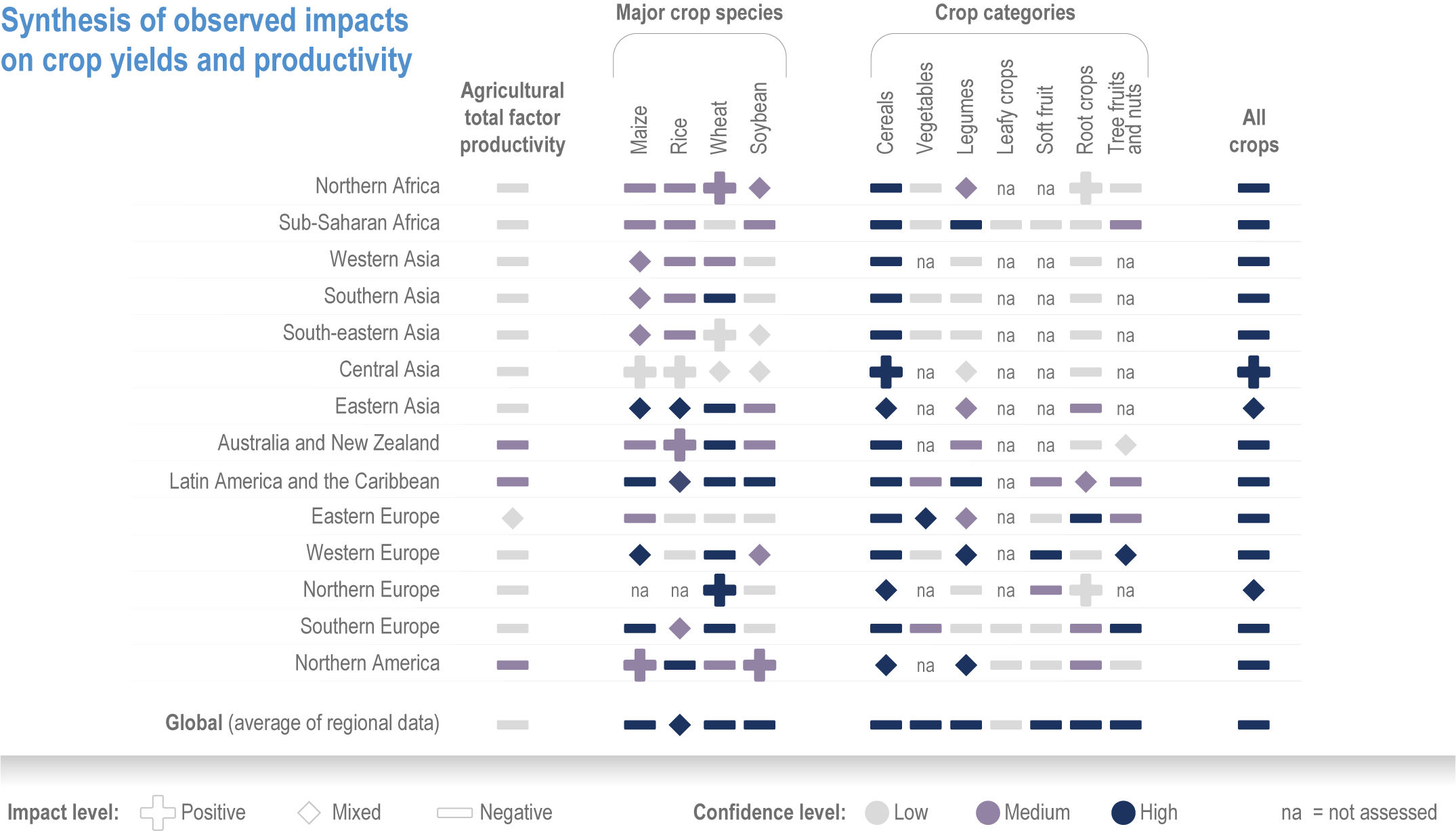
Global yields of major crops per unit land area have increased 2.5- to 3-fold since 1960. Plant breeding, fertilisation, irrigation and integrated pest management have been the major drivers, but many studies have found significant impacts from recent climate trends on crop yield (high confidence) (Figure 5.3; see Section 5.2.1 for the change attributable to anthropogenic climate change).
Climate impacts for the past 20–50 years differ by crops and regions. Positive effects have been identified for rice and wheat in Eastern Asia, and for wheat in Northern Europe. The effects are mostly negative in Sub-Saharan Africa, South America and Caribbean, Southern Asia, and Western and Southern Europe. Climate factors that affected long-term yield trends also differ between regions. For example, in Western Africa, 1°C warming above preindustrial climate has increased heat and rainfall extremes, and reduced yields by 10–20% for millet and 5–15% for sorghum (Sultan et al., 2019). In Australia, declined rainfall and increased temperatures reduced yield potential of wheat by 27%, accounting for the low yield growth between 1990 and 2015 (Hochman et al., 2017). In Southern Europe, climate warming has negatively impacted yields of almost all major crops, leading to recent yield stagnation (Moore and Lobell, 2015; Agnolucci and De Lipsis, 2020; Brás et al., 2021).
Ortiz-Bobea et al. (2021) analysed agricultural total factor productivity (TFP), defined as the ratio of all agricultural outputs to all agricultural inputs, and found that, while TFP has increased between 1961 and 2015, the climate change trends reduced global TFP growth by a cumulative 21% over a 55-year period relative to TFP growth under counterfactual non-climate change conditions. Greater effects (30–33%) were observed in Africa, Latin America and the Caribbean (Figure 5.3).

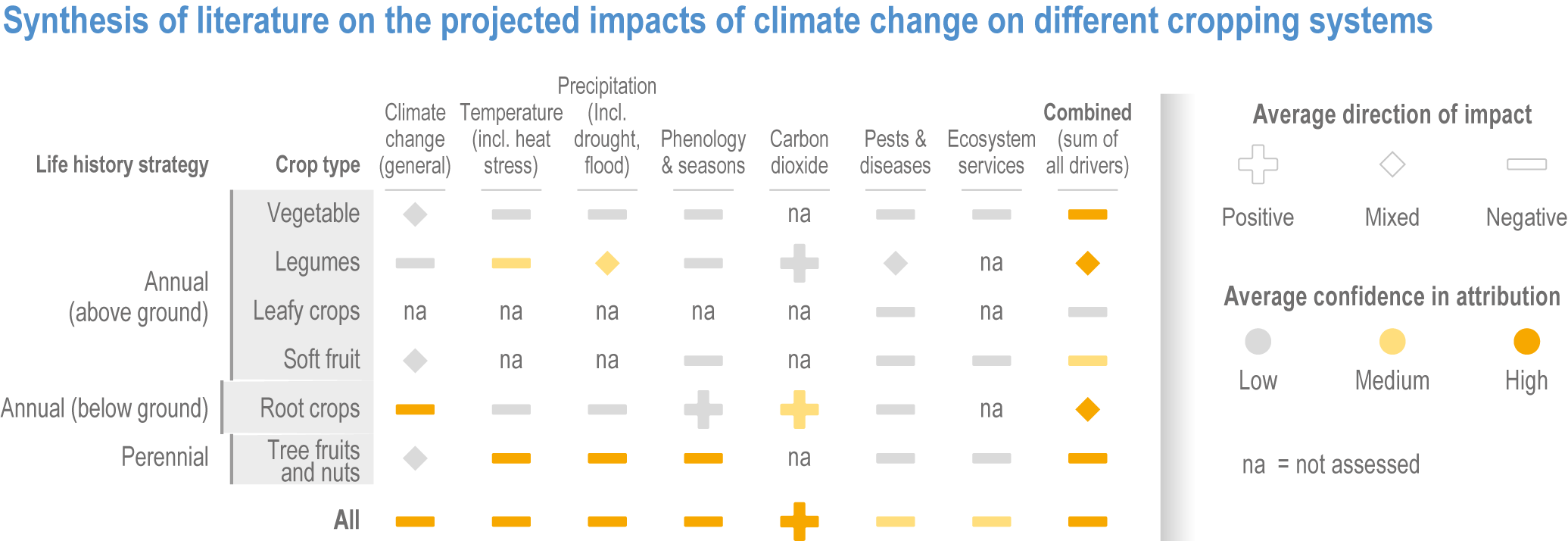
Figure 5.3 | Synthesis of literature on observed impacts of climate change on productivity by crop type and region. The figure draws on >150 articles categorized by: agriculture total factor productivity including literature estimating all agricultural outputs in a region; major crop species including literature assessing yield changes in the four major crops; crop categories including productivity changes (yield, quality and other perceived changes) in a range of crops with different growth habits. The assessment uses literature published since AR5, although the timespan often extends prior to 2014. The direction of the effect and the confidence are based on the reported impacts and attribution, and on the number of articles. See SM5.1 and SM5.2 for details.
Climate variability is a major source of variation in crop production (Ray et al., 2015; Iizumi and Ramankutty, 2016; Frieler et al., 2017; Cottrell et al., 2019)(Table SM5.1). Weather signals in yield variability are generally stronger in productive regions than in the less productive regions (Frieler et al., 2017), where other yield constraints exist such as pests, diseases and poor soil fertility (Mills et al., 2018; 5.2.2). Nevertheless, yield variability in less productive regions has severe impacts on local food availability and livelihood (high confidence) (FAO, 2021).
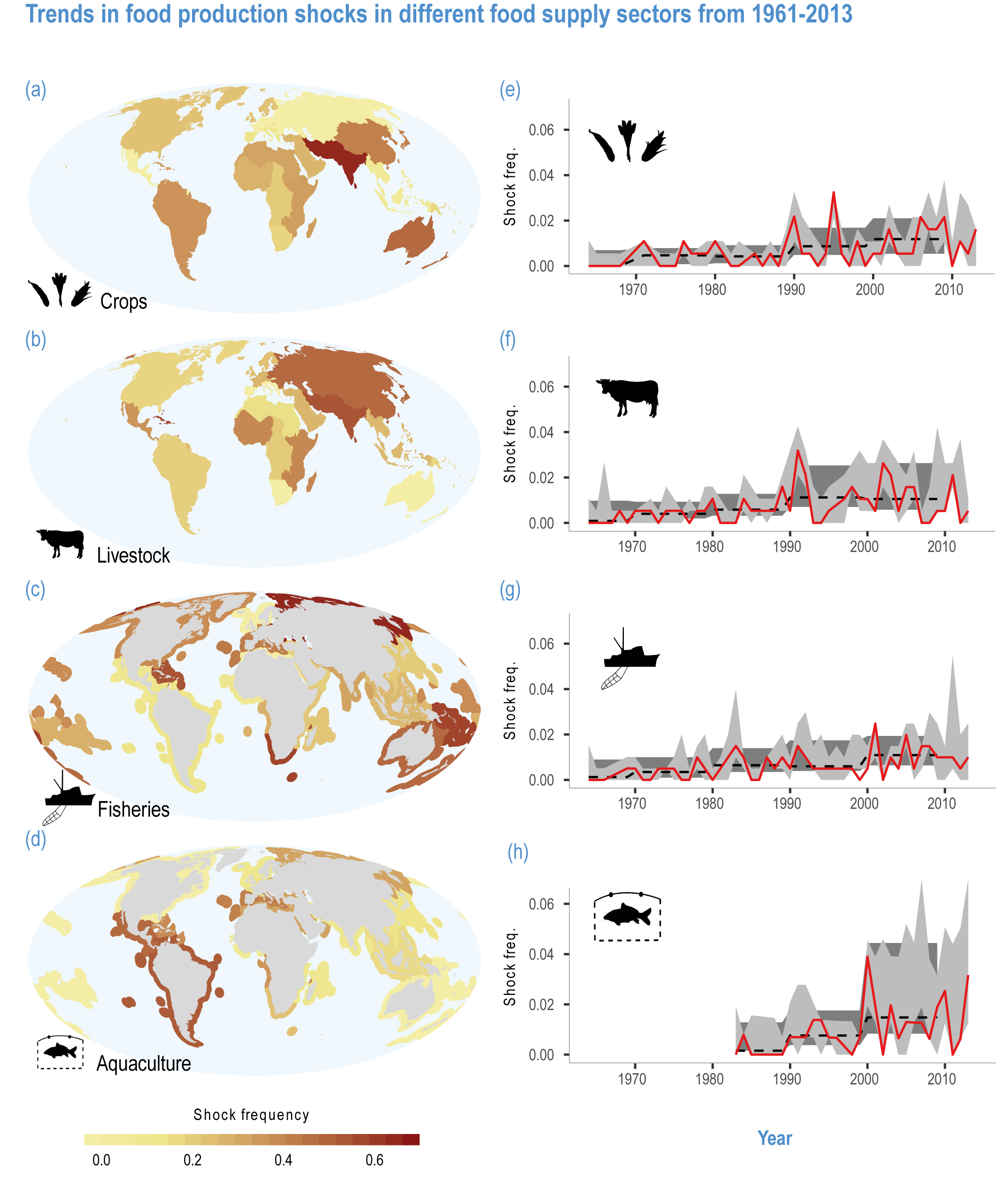
Climate-related hazards that cause crop losses are increasing (medium evidence, high agreement ) (Cottrell et al., 2019; Mbow et al., 2019; Brás et al., 2021; FAO, 2021; Ranasinghe et al., 2021). Drought-related yield losses have occurred in about 75% of the global harvested area (Kim et al., 2019b) and increased in recent years (Lesk et al., 2016). Heatwaves have reduced yields of wheat (Zampieri et al., 2017) and rice (Liu et al., 2019b). The combined effects of heat and drought decreased global average yields of maize, soybeans and wheat by 11.6%, 12.4% and 9.2%, respectively (Matiu et al., 2017). In Europe, crop losses due to drought and heat have tripled over the last five decades (Brás et al., 2021), pointing to the importance of assessing multiple stresses. Globally, floods also increased in the past 50 years, causing direct damages to crops and indirectly reduced yields by delaying planting, which cost 4.5 billion USD in the 2010 flood in Pakistan and 572 million USD in the 2015 flood in Myanmar (FAO, 2021).
5.4.1.2 Observed impacts on other crops (vegetables, fruit, nut and fibre)
The impact of climate change on these diverse crop types is under-researched and uncertain (Manners and van Etten, 2018; Alae-Carew et al., 2020); there are reports of positive impacts in some cases, but overall the observed impacts are negative across all crop categories (Figure 5.3).
Above-ground annual crops consumed as vegetables, fruits or salad are essential for food security and nutrition (5.12). In temperate regions, climate change can result in higher yields (Potopová et al., 2017; Bisbis et al., 2018), while in subtropical/tropical regions, negative impacts from heat and drought take precedence (Scheelbeek et al., 2018). Different species have different sensitivities to heat and drought (Prasad et al., 2017; Scheelbeek et al., 2018) and to combinations of stresses (Zandalinas et al., 2018). Above-ground vegetables are especially vulnerable to heat and drought stress during pollination and fruit set, resulting in negitive impacts on yield (Daryanto et al., 2017; Sita et al., 2017; Brás et al., 2021) and harvest quality (Mattos et al., 2014; Bisbis et al., 2018). Growers have already seen negative impacts from the expansion of pest and disease agents due to warming (Section 5.4.1.3; Figure 5.3).
Below-ground vegetables include starchy roots and tubers that form a regular diet in many parts of the tropics and subtropics. Warming and climate variability has altered the rate of tuber development, with yield impacts varying by location, including yield increases in some cases (Shimoda et al., 2018; Ray et al., 2019). These crops are considered stress tolerant but are more sensitive to drought than cereals (Daryanto et al., 2017). Impacts on water supply are critical as root crops are water-demanding for long periods, and highly sensitive to drought and heat events during tuber initiation (Dua et al., 2013; Potopová et al., 2017; Brás et al., 2021).
Among perennial tree crops, only grapevine, olive, almond, apple, coffee and cocoa have received significant research attention. Concerns about climate impacts on harvest quality are widespread (Figure 5.3) (Barnuud et al., 2014; Bonada et al., 2015). In higher-latitude regions, the primary concern is the effect of temperature variability on harvest stability, pests and diseases and phenology (including fulfilment of winter chill requirements and risks due to early emergence in spring), (El Yaacoubi et al., 2014; Ramírez and Kallarackal, 2015; Santos et al., 2017; Gitea et al., 2019). In lower-latitude regions, information is limited, but studies are focused on increased tree mortality and yield loss due to drought, heat and impacts from variability in the timing of the wet and dry seasons (Glenn et al., 2013; Ramírez and Kallarackal, 2015); see Box 5.7). In fruit trees, warming and climate variability have already affected fruit quality, such as acidity and texture in apples, or skin colour in grape berries (Sugiura et al., 2013; Sugiura et al., 2018). The reliability and stability of harvests has been impacted by climate variability, changes in the distribution of pests and pathogens (Seidel, 2014; Bois et al., 2017), and the mismatch of important phenological events (such as bud emergence and flowering) (Guo and Shen, 2015; Legave et al., 2015; Ito et al., 2018; Vitasse et al., 2018). Perennial crops are particularly vulnerable to these impacts as they are exposed throughout the year, with little potential for growers to adjust planting date or location. Negative impacts via disruption to phenology and pest dynamics are best studied in grapevine (see Box 5.2).
Among the fibre crops, cotton is particularly well studied. As cotton is heat tolerant and yield increases with extra plant growth, positive effects of increasing temperature are expected, but observed impacts have been mixed due to negative impacts on phenology and plant water status (Traore et al., 2013; Chen et al., 2015a; Cho and McCarl, 2017). Negative impacts of climate change due to proliferation of the pest cotton bollworm are widely reported (Ouyang et al., 2014; Huang and Hao, 2020).
The impacts of climate change on water availability (rainfall and irrigation supply) are an emerging issue. Increased occurrence of drought combined with limited access to irrigation water is already a key constraint; for example, Californian almonds are predicted to increase their potential geographical range under climate warming (Parker, 2018), yet a trend of increasing drought has already resulted in trees being removed due to lack of access to irrigation water (Keppen and Dutcher, 2015; Kerr et al., 2018; Reisman, 2019).
5.4.1.3 Observed impacts on pests, diseases and weeds
AR5 and SRCCL (IPCC, 2019) indicated that more frequent outbreaks and area expansion of pests and diseases are serious concerns under climate change but are under-researched because of the difficulties in assessing multi-species interactions (Porter et al., 2014; Mbow et al., 2019). High-quality historical and current observational data to detect changes in pests and diseases attributable to recent trends in climate are still limited.
Bebber (2013) found significant poleward expansions of many important groups of crop pests and pathogens since 1960, with an average shift of 2.7 km yr −1. Different pest species populations respond differently to ongoing climate change, with some shifting, contracting or expanding their current distribution range and others persisting or disappearing in their current range (high confidence). These asymmetric distribution changes can create novel species combinations or decouple existing ones (Pecl et al., 2017; Hobbs et al., 2018), but their consequences on future crop production and food security are hard to predict. Multi-species climate change experiments are rare (Bonebrake et al., 2018), but one study shows that under future climates different pest assemblages of interacting species may alter levels of damage to crops compared with that by only one species (Crespo-Perez et al., 2015). Some studies highlight the importance of location-specific species interactions for more realistic projections of pest distribution, performance and damage to crops, which in turn would allow more effective prevention and pest control strategies (Wilson et al., 2015; Carrasco et al., 2018).
Weeds are recognised as a primary constraint on crop production (Oerke, 2006), rangelands (DiTomaso et al., 2017) and forests (Webster et al., 2006). Climate change could favour the growth and development of weeds over crops with negative consequences for desired plants in managed systems (medium evidence, high agreement ) (Peters et al., 2014; Ziska and McConnell, 2016). First, changes in temperature and precipitation alter the range, composition and competitiveness of native and invasive weeds (Bradley et al., 2010). Second, rising concentrations of CO2 enhance growth of C3 species (~85% of plant species, including many weeds) (Ogren and Chollet, 1982; Ziska, 2003), and increase plant water use efficiency with potentially strong effects on invasive plant species establishment (Smith et al., 2000; Belote et al., 2004; Blumenthal et al., 2013).
Some invasive species within unmanaged areas will expand further, proliferate and be more competitive under climate change as they may benefit from increased resource ability (e.g., additional CO2, enhanced precipitation) (Bradley et al., 2010; Kathiresan and Gualbert, 2016; Merow et al., 2017; Ramesh et al., 2017; Waryszak et al., 2018), which will make chemical weed control more problematic (medium evidence, high agreement ) (Waryszak et al., 2018; Ziska, 2020). The range of other invasive weeds may become static, or even decline (Bradley et al., 2016; Buckley and Csergo, 2017). A recent meta-analysis also supports that invasive plants respond more favourably to elevated CO2 concentrations and elevated temperatures than native plants (Korres et al., 2016; Liu et al., 2017). Movement of invasive species into low-fertility areas, however, could provide resource opportunities, especially if agriculture in those areas is limited (Randriambanona et al., 2019).
Rising CO2 concentrations and climate change could reduce herbicide efficacy (medium evidence, high agreement ). These reductions may be associated with physical environmental changes (precipitation, wind speed) that influence herbicide coverage (Ziska, 2016) as well as direct effects of CO2 on plant biochemistry and herbicide resistance (Refatti et al., 2019). Increasing CO2 levels and altered temperature and precipitation are therefore projected to affect all aspects of weed biology (Peters et al., 2014; Ziska and McConnell, 2016), including establishment (Bradley et al., 2016), competition (Fernando et al., 2019), distribution, (Castellanos-Frías et al., 2016) and management (Waryszak et al., 2018).
A warmer climate increases the need for pesticides (Shakhramanyan et al., 2013; Ziska, 2014; Delcour et al., 2015; Zhang et al., 2018). Increases in temperature and CO2 concentration may reduce pesticide efficiency by altering its metabolism, or accelerating detoxification (Matzrafi et al., 2016; Matzrafi, 2019). Intense rainfall also reduces persistence (Delcour et al., 2015). Invasive pests and pathogens impose an additional cost for the society (Bradshaw et al., 2016). Rapid and large-scale dispersal of pests is already a major threat to food security, as exemplified by the recent outbreak of desert locusts (see Box 5.8), indicating the importance of international cooperation. Taken together, the need for control of pests, disease and weeds will increase under climate change (medium evidence, high agreement ). The use of toxic agricultural chemicals also has human health and environmental risks (Whitmee et al., 2015; IPBES, 2019). Surveillance for monitoring pest distribution and damages, climate-relevant pest risk analysis, and climate-smart strategies for controlling pests with minimal impacts on human and environmental health are important tools in the face of climate change (IPPC Secretariat, 2021).
5.4.1.4 Observed impacts of ozone on crops
Tropospheric (i.e., the lowest 6–10 km of the atmosphere) ozone exacerbates negative impacts of climate change (high confidence) (Mattos et al., 2014; Chuwah et al., 2015; McGrath et al., 2015; Bisbis et al., 2018; Mills et al., 2018; Scheelbeek et al., 2018). Ozone is an air pollutant and short-lived GHG that affects air quality and global climate. It is a strong oxidant that reduces physiological functions, yield and quality of crops and animals. Surface ozone concentration has increased substantially since the late 19th century (Cooper et al., 2014; Forster et al., 2021; Gulev et al., 2021; Szopa et al., 2021) and in some locations and times reaches levels that harm plants, animals and human (high confidence) (Fleming et al., 2018).
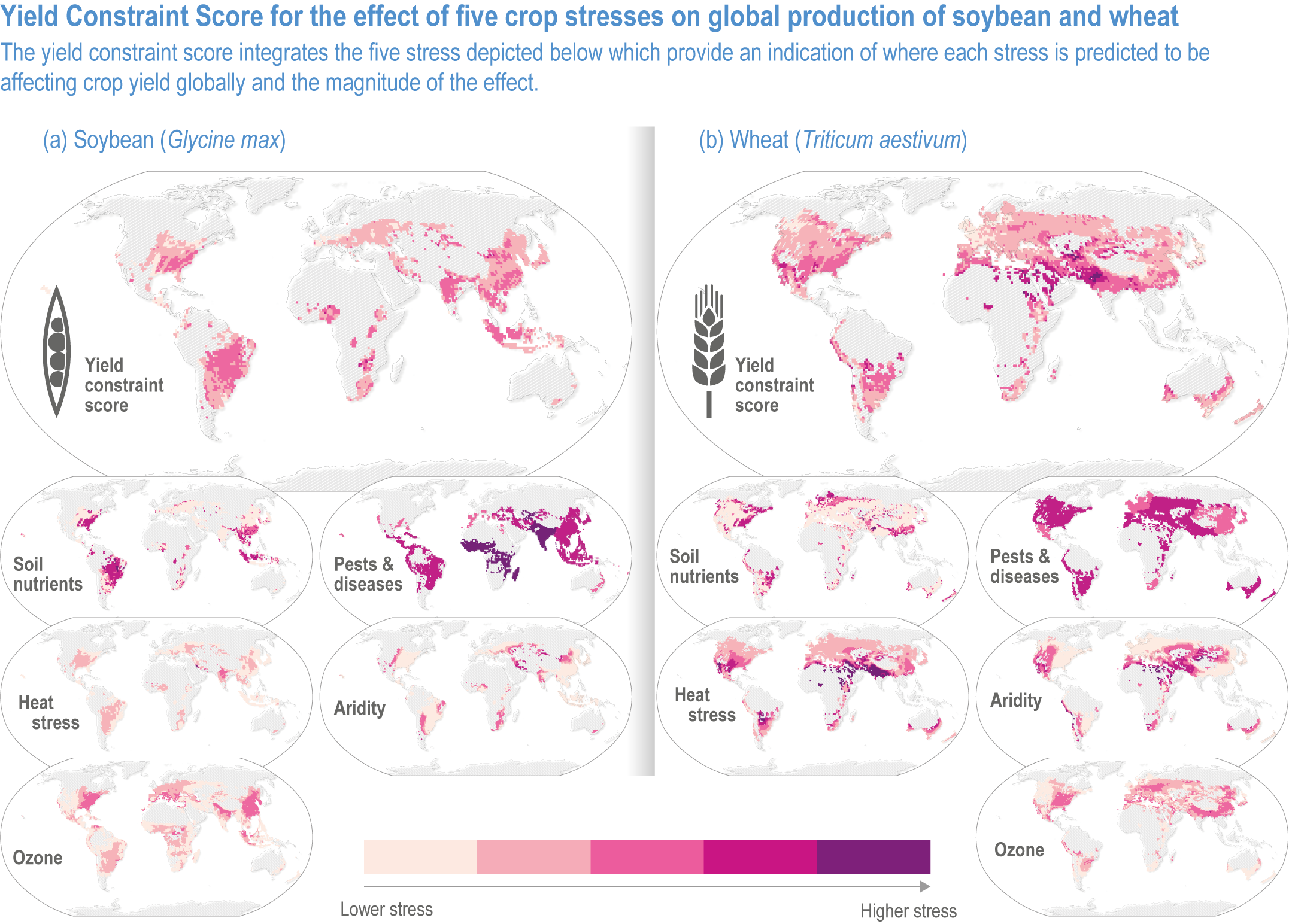
Mills (2018) estimated global distributions of current yield losses of major crops due to ozone, pest and diseases, heat, and aridity (Figure 5.4). Ozone-induced yield losses in 2010–2012 averaged 12.4%, 7.1%, 4.4% and 6.1% for soybean, wheat, rice and maize, respectively. Spatial variation in yield losses is similar among different stresses; areas with a large loss due to ozone are also at high risk of yield losses due to pest and diseases and heat. Many vegetable crops are also susceptible to ozone, which will adversely impact quality and quantity (Mattos et al., 2014; Bisbis et al., 2018; Scheelbeek et al., 2018).
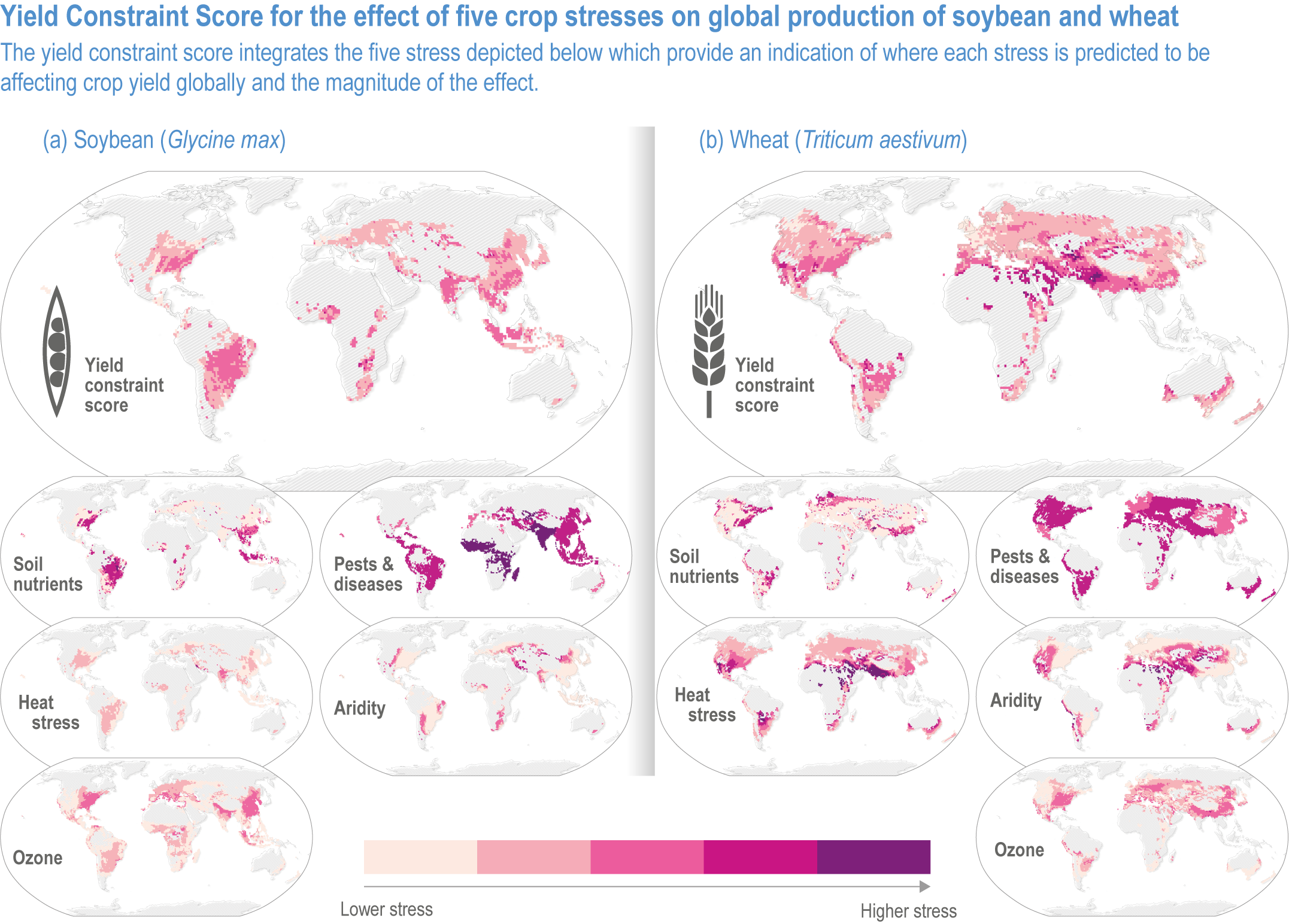
Figure 5.4 | The global effects of five biotic and abiotic stresses on soybean and wheat. All data are presented for the 1 × 1° (latitude and longitude) grid squares where the mean production of soybean or wheat was >500 tonnes (0.0005 Tg). The effect of each stress on yield is presented as a Yield Constraint Score (YCS) on a scale of 1–5, where 5 is the highest level of stress from ozone, pests and diseases, heat stress and aridity (Mills et al., 2018). Data are available at Sharps et al. (2020). See Annex I: Global to Regional Atlas for all four crops.
The estimated yield loss does not account for interactions with other climatic factors. Temperatures enhance not only ozone production but also ozone uptake by plants, exacerbating yield and quality damage. Burney (2014) estimated current yield losses due to the combined effects of ozone and heat in India at 36% for wheat and 20% for rice. Schauberger et al. (2019a) found global yield losses, ranging from 2% to 10% for soybean and 0% to 39% for wheat with a model that accounts for temperature, water and CO2 concentration on ozone uptake.
Box 5.1: Evidence for Simultaneous Crop Failures Due to Climate Change
Simultaneous yield losses across major producing regions can be a threat to food security but had not been quantified by the time of AR5. Large-scale sea surface temperature (SST) oscillations greatly influence global yield of major crops (high confidence) (Anderson et al., 2019b; Najafi et al., 2019; Ubilava and Abdolrahimi, 2019; Heino et al., 2020; Iizumi et al., 2021b) and food prices (Ubilava, 2018). Some studies showed that crop yields in different regions covaried with SST oscillations, suggesting occurrences of tele-connected yield failures (crop losses caused by related factors in distant regions; Table Box 5.1.1) (medium confidence). Evidence of synchronised crop failures increasing with ongoing climate change is still limited.
Table Box 5.1.1 | A summary of peer-review papers detecting synchronised yield losses.
Regions/ commodities | Period studied | Observed impacts | Climate driver | Evidence for multiple breadbasket failures | Evidence for increasing risks due to multiple breadbasket failures | Reference |
Global breadbaskets for maize, rice, sorghum and soybean | 1961–2013 | Not only yields of each crop covaried in many countries, but also those of different crops, maize in particular, covaried with other crops. | SST anomalies, atmospheric and oceanic in- dices, air temperature anomalies and Palmer Drought Severity Index | High | NA | Najafi et al. (2019) |
Global breadbaskets for wheat, soybean and maize | 1980–2010 | Climate modes (El Niño-Southern Oscillation (ENSO), the Indian Ocean Dipole (IOD), tropical Atlantic variability (TAV) and the North Atlantic Oscillation (NAO)) account for 18%, 7% and 6% of global maize, wheat and soybean production variability, respectively. ENSO events sometimes offset yield reductions in some places by increases in other places (e.g., soybean yields in the USA and southeast South America). Since 1961, ENSO in 1983 was the only climate mode that showed global synchronous crop failures. | Climate modes | Medium (1983) | NA | |
Global breadbaskets for wheat, soybean and maize | Climate modes induce yield variability in major breadbaskets, e.g., ENSO affects about half of maize and wheat areas. IOD and ENSO influence wheat in Australia. ENSO affects soybean in northern South America. | Climate modes | Medium | NA | ||
67 maize producing countries | 1961–2017 | SST anomalies from the 1980–2010 base period in the Niño3.4 region, a rectangular area bounded by 120°W–170°W and 5°S–5° is used as a driver. Maize yields are tele-connected among the southeastern tier of Sub-Saharan Africa, as well as Central America, South Asia and Australia. A 1° increase in SST reduced maize yield by up to 20% in these countries. | Climate modes (SST), precipitation | Medium | NA | |
Global breadbasket (the USA, Argentina, Europe, Russia/Ukraine, China, India, Australia, Indonesia and Brazil) | 1967–2012 | Likelihood of simultaneous climate risks increased from 1967–1990 to 1991–2012 in the global breadbasket (lower 25th yield deviation percentile events at province level) for wheat, soybean and maize, but not rice. Likelihood of simultaneous climate risks increased from 1967–1990 to 1991–2012 in China (lower 25th yield deviation percentile events at province level). | Unspecified | Medium | Medium | |
Global | 1961–2008 | Synchronous yield losses among major breadbaskets within each commodity, such as maize and soybean, decreased between 1961 and 2008. In contrast, synchronous yield variation between crops has increased. Under a scenario of synchronisation of all four crops, the global maximum production losses for rice, wheat, soybean and maize are estimated to reach between −17% and −34%. | Unspecified | Medium | Medium |
5.4.2 Assessing Vulnerabilities within Production Systems
Since AR5, vulnerability assessment has become a pivotal component of risk analysis associated with climate hazards, climate change and climate variability (UNDRR, 2019). Vulnerability assessment can be sectoral or regional but involves social and ecological indicators. This section presents examples of vulnerability assessment to climatic hazards and social vulnerabilities.
5.4.2.1 Vulnerability to climatic hazards
Drought is a major risk component in cropping systems globally, with substantial economic loss (Kim et al., 2019b), livelihood impacts (Shiferaw et al., 2014; Miyan, 2015) and ultimately health risks such as malnutrition (Phalkey et al., 2015; Cooper et al., 2019). Vulnerability to drought can be estimated with a range of indicators (Hagenlocher et al., 2019). Meza (2020) showed that drought risks could be exacerbated or moderated by regional differences in vulnerability (Figure 5.5). For instance, high-level risks observed in southern Africa, western Asia and central Asia result from high vulnerability (low coping capacity), whereas risk levels are relatively low despite the high exposure by relatively high adaptive capacity to drought in other regions.
Figure 5.5 | Hazard and exposure indicator score (a), vulnerability index (b) and drought risk index (c), for rainfed agricultural systems between 1986 and 2015. Drought hazard indicator is defined as the ratio of actual crop evapotranspiration to potential crop evapotranspiration, calculated for 24 crops. Vulnerability index is the country-scale weighted average of a total of 64 indicators including social and ecological susceptibility indicators, and coping capacity. Risk index is calculated by multiplying hazard/exposure indicator score and vulnerability index (Meza et al., 2020).
Regional-scale assessment also highlights the importance of adaptive capacity. For instance, rice and maize production in Viet Nam Mekong Delta has high exposure to multiple climate hazards such as flooding, sea level rise, salinity intrusion and drought (Parker et al., 2019). Risks can be moderated by a relatively high adaptive capacity because of infrastructure, resources and high education levels (Parker et al., 2019). Another regional study demonstrated that erratic rains and high temperatures in southern and southeastern Africa increased the vulnerability of agricultural soils, thereby exacerbating impacts of prolonged and frequent droughts (Sonwa et al., 2017a; See also Box 5.4).
Farm-scale assessment exemplifies context-sensitive vulnerability to climate hazards. Studies of coffee growers in Central America demonstrated that key vulnerability indicators varied greatly between regions and between farms, ranging from a lack of labour, postharvest infrastructure, conservation practices and transport that limits access to market, technical and financial assistance (Baca et al., 2014; Bouroncle et al., 2017). These region- and scale-specific vulnerability indicators assist in identifying ways to enhance resilience to climate hazards (high confidence).
5.4.2.2 Inequities in cropping systems—other crops and regional disparities
While those working with major crops have benefited from the release of new cultivars, those growing other crops are typically reliant on a heritage cultivars or landraces. While Indigenous knowledge and local smallholder knowledge and practices play an important role in supporting agrobiodiversity which provides genetic diversity resistant to climate-related stresses, a global and national focus in international research, subsidies and support for a few crop species has contributed to an overall decline in agrobiodiversity (FAO, 2019e; Song et al., 2019) Similarly, there is a lack of agronomic innovation and research to service ‘minor’ crops (Moriondo et al., 2015; Manners and van Etten, 2018). Even some high-value commodities grown outside high-income countries suffer from imbalances in the focus of available credit, research and innovation (Section 5.4.4.3; Glover, 2014; Fischer, 2016; Farrell et al., 2018). There is a possibility that a lack of adaptive capacity and policy support will drive these growers to move away from these diverse crops, further reducing the resilience of food systems by increasing risk of crop loss from pests, disease and drought and potential loss of Indigenous or local knowledge (Section 5.13.5, Table Box 5.1.1). In the Andean Altiplano of Bolivia, for example, Indigenous farmers have traditionally managed a diverse set of native crops which are drought and frost-tolerant, using cultural practices of seed selection and exchange, but have faced an increase in pests and diseases and a decline of traditional crops due to climate-change-related stresses, out-migration and intensification drivers (Meldrum et al., 2018).
5.4.2.3 Gender and other social inequities
Social inequities such as gender, ethnicity and income level, which vary by time and place and may overlap, can compound vulnerability to climate change for producers within cropping systems (high confidence) (Table 5.3, Arora-Jonsson, 2011; Djoudi et al., 2013; Carr and Thompson, 2014; Mbow et al., 2019; Rao et al., 2019a; Nyantakyi-Frimpong, 2020a). Rather than binary and static categories (i.e., men versus women), social vulnerabilities are dynamic and intersect; to understand vulnerability, the specific socio-cultural identities and political and environmental context need to be studied in relation to climate stress (Thompson-Hall et al., 2016; Rao et al., 2019a; Nyantakyi-Frimpong, 2020a).
Table 5.3 | Examples of social inequities in cropping systems that compound climate change vulnerability.
Social inequity | How social inequity increases vulnerability to climate change in cropping systems |
Gender inequity can create and worsen social vulnerability to climate change impacts within cropping systems (high confidence) (Carr and Thompson, 2014; Sugden et al., 2014; Nyantakyi-Frimpong and Bezner-Kerr, 2015; Rao et al., 2019a; Ebhuoma et al., 2020; Nyantakyi-Frimpong, 2020a; see Cross-Chapter Box GENDER in Chapter 18). |
|
Globally, smallholder food producers are more vulnerable than large-scale producers to climate change impacts (high confidence). |
|
Farmworkers are another social group with heightened vulnerability to climate change (medium confidence). |
|
5.4.3 Projected Impacts
5.4.3.1 Advances in the characterisation of the effects of elevated atmospheric CO2
Elevated CO2 concentrations stimulate photosynthesis rates and biomass accumulation of C3 crops, and enhance crop water use efficiency of various crop species, including C4 crops (high confidence) (Kimball, 2016; Toreti et al., 2020). Perennial crops and root crops may have a greater capacity for enhanced biomass under elevated CO2 concentrations, although this does not always result in higher yields (Glenn et al., 2013; Kimball, 2016).
Recent FACE studies found that the effects of elevated CO2 are greater under water-limited conditions (medium confidence) (Manderscheid et al., 2014; Fitzgerald et al., 2016; Kimball, 2016), which was generally reproduced by crop models (Deryng et al., 2016). However, drought sometimes negates the CO2 effects (Jin et al., 2018).
There are significant interactions between CO2, temperature, cultivars, nitrogen and phosphorous nutrients (Kimball, 2016; Toreti et al., 2020): positive effects of rising CO2 on yield are significantly reduced by higher temperatures for soybean, wheat and rice (medium confidence) (Ruiz-Vera et al., 2013; Cai et al., 2016; Gray et al., 2016; Hasegawa et al., 2016; Obermeier et al., 2016; Purcell et al., 2018; Wang et al., 2018). In above-ground vegetables, elevated CO2 can in some cases reduce the impact of other climate stressors, while in others the negative impacts of other abiotic factors negate the potential benefit of elevated CO2 (Bourgault et al., 2017; Bourgault et al., 2018; Parvin et al., 2018; Parvin et al., 2019). Significant variation exists among cultivars in yield response to elevated CO2, which is positively correlated with yield potential in rice and soybean, suggesting the potential to develop cultivars for enhanced productivity under future elevated [CO2] (Ainsworth and Long, 2021).
Elevated CO2 reduces some important nutrients such as protein, iron, zinc and some grains, fruit or vegetables to varying degrees depending on crop species and cultivars (high confidence) (Mattos et al., 2014; Myers et al., 2014; Dong et al., 2018; Scheelbeek et al., 2018; Zhu et al., 2018a; Jin et al., 2019; Ujiie et al., 2019). This is of particular relevance for fruit and vegetable crops given their importance in human nutrition (high confidence) (see Section 5.12.4 for potential impacts on nutrition; Nelson et al., 2018; Springmann et al., 2018). Recent experimental studies (Section 5.3.2), however, show some complex and counteracting interactions between CO2 and temperature in wheat, soybean and rice; heat stress negates the adverse effect of elevated CO2 on some nutrient elements (Macabuhay et al., 2018; Kohler et al., 2019; Wang et al., 2019b). The CO2 by temperature interaction for grain quality needs to be better understood quantitatively to predict food nutritional security in the future.
5.4.3.2 Projected impacts on major crop production
AR5 Chapter 7 estimated global crop yield reduction due to climate change to be about 1% per decade (Porter et al., 2014), similar to the previous assessment reports (Porter et al., 2019). Additional research confirms that climate change will disproportionately affect crop yields among regions, with more negative than positive effects being expected in most areas, especially in currently warm regions, including Africa and Central and South America (high confidence).
A systematic literature search between 2014 and 2020 resulted in about 100 peer-reviewed papers that simulated crop yields of four major crops (maize, rice, soybean and wheat) using Coupled Model Intercomparison Project Phase 5 (CMIP5) data (Hasegawa et al., 2021b). Most studies focus on the relative change in crop yields due to climate change but do not consider technological advances. Nevertheless, they provide useful insights into time-, scenario- and warming-degree-dependent impacts of climate change.
The impact of climate change on crop yield without adaptation projected in the 21st century is generally negative even with the CO2 fertilisation effects, with the overall median per-decade effect being −2.3% for maize, −3.3% for soybean, −0.7% for rice and −1.3% for wheat, which is consistent with previous IPCC assessments (Porter et al., 2014). The effects vary greatly within each crop, timeframe and RCP, but show a few common features across crops (Figure 5.6a ). Differences in the projected impacts between RCPs are not pronounced by mid-century. From then onward, the negative effect becomes more pronounced under RCP8.5, notably in maize. Rice yields show less variation across models than other crops presumably because simulations are mostly under irrigated conditions. A part of the uncertainty in the projection is due to regional differences (Figure 5. 6b). Negative impacts on cereals are projected in Africa and Central and South America at the end of the century, which agrees with the previous studies (Aggarwal et al., 2019; Porter et al., 2019).
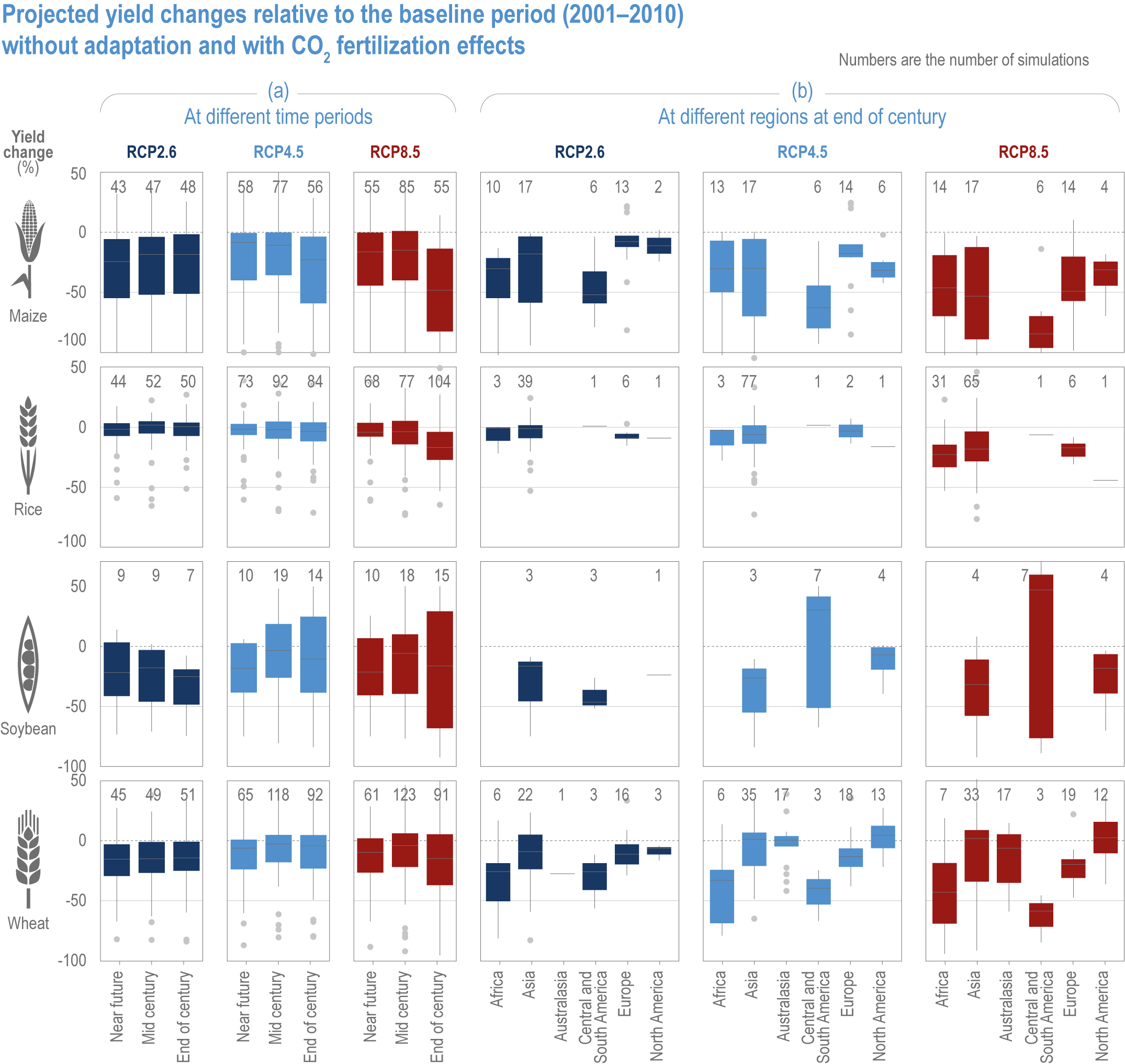
Figure 5.6 | Projected yield changes relative to the baseline period (2001–2010) without adaptation and with CO2fertilisation effects (Hasegawa et al. , 2021b). The box is the interquartile range (IQR), and the middle line in the box represents the median. The upper and lower end of whiskers are median 1.5 × IQR ± median. Open circles are values outside the 1.5 × IQR. (a) At different time periods (near future, NF, baseline to 2039; mid-century, MC, 2040–2069; end-century, EC, 2070–2100) under three RCPs, and (b) at different regions at EC.
The differences due to regions, RCPs and timeframes are related to the current temperature level and degree of warming (Figure 5.7). The projected effects of climate change are positive where current annual mean temperatures (Tave) are below 10°C, but they become negative with Tave above around 15°C. At Tave> 20°C, even a small degree of warming could result in adverse effects. In maize, negative effects are apparent at almost all temperature zones. A new study using the latest climate scenarios (Coupled Model Intercomparison Project Phase 6, CMIP6) and global gridded crop model ensemble projected that climate change impacts on major crop yields appear sooner than previously anticipated, mainly because of warmer climate projections and improved crop model sensitivities (Jägermeyr et al., 2021).
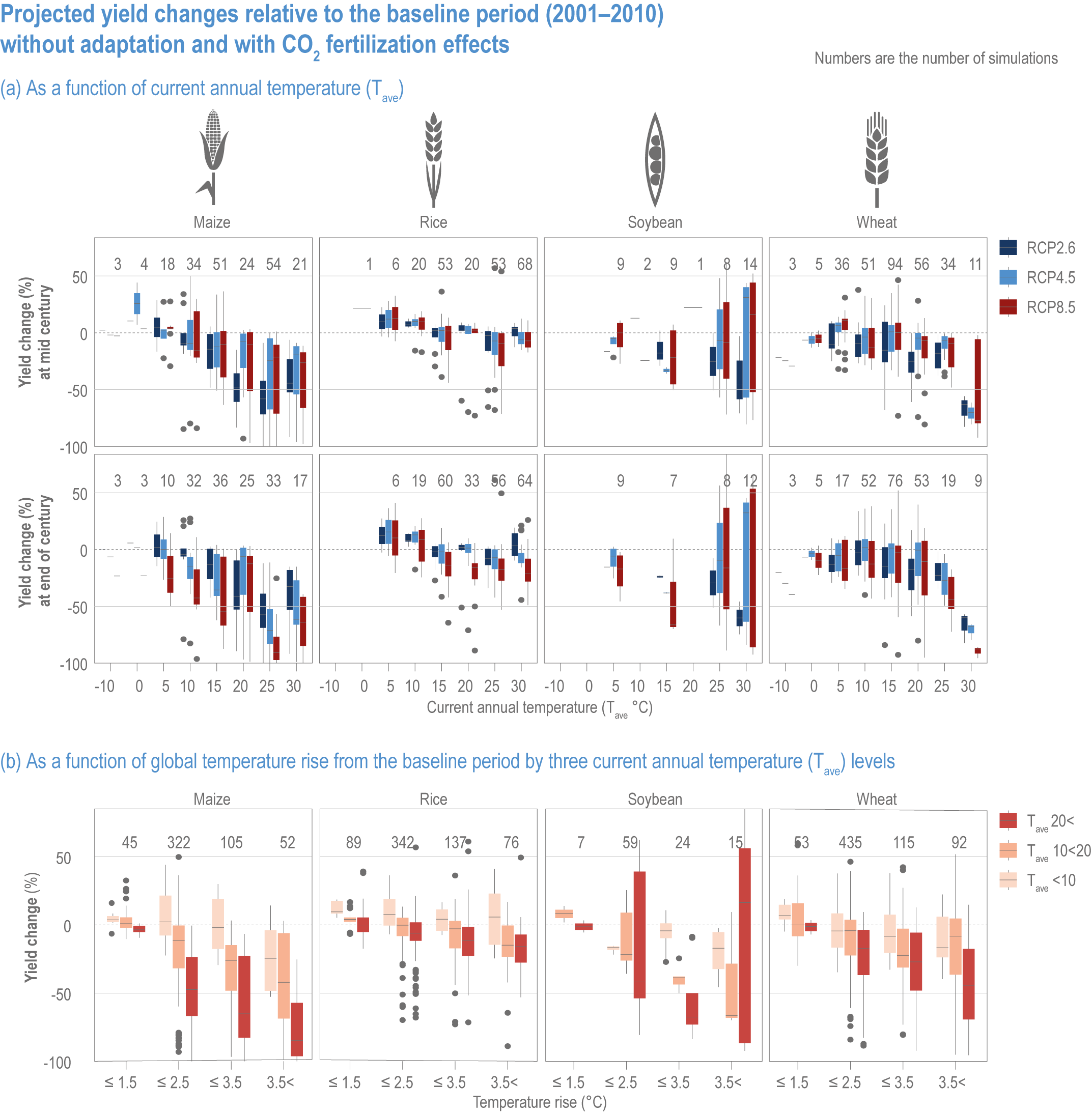
Figure 5.7 | Projected yield changes relative to the baseline period (2001–2010) without adaptation and with CO2fertilisation effects (Hasegawa et al. , 2021b). (a) Mid-century (MC, 2040–2069) and end-century (EC, 2070–2100) projections under three RCP scenarios as a function of current annual temperature (Tave), (b) as a function of global temperature rise from the baseline period by three Tave levels. See Figure. 5.6 for legends.
As noted in Section 5.3.1, most simulations do not fully account for responses to pests, diseases, long-term change in soil, and some climate extremes (Rosenzweig et al., 2014), but studies are emerging to include some of these effects. For example, based on the temperature response of insect pest population and metabolic process, global yield losses of rice, maize and wheat are projected to increase by 10–25% per degree Celsius of warming (Deutsch et al., 2018). Rising temperatures reduce soil carbon and nitrogen, which in turn exacerbate the negative effects of +3°C warming on yield from 9% to 13% in wheat and from 14% to 19% in maize (Basso et al., 2018).
A few studies have examined possible occurrences of tele-connected yield losses (5.4.1.2) using future climate scenarios. Tigchelaar (2018) estimated that, for the top four maize-exporting countries, the probability that simultaneous production losses greater than 10% occur in any given year increases from 0% to 7% under 2°C warming and to 86% under 4°C warming. Gaupp (2019) estimated that risks of simultaneous failure in maize would increase from 6% to 40% at 1.5°C and to 54% at 2°C warming, relative to the historical baseline climate. Large-scale changes in SST are the major factors causing simultaneous variation in climate extremes, which are projected to intensify under global warming (Cai et al., 2014; Perry et al., 2017). Consequently, risks of simultaneous yield losses in major food-producing regions will also increase with global warming levels above 1.5°C (medium confidence). Further examination is needed for the effects of spatial patterns of these extremes on breadbaskets in relation to SST anomalies under more extreme climate scenarios.
Future surface ozone concentration is highly uncertain (Fiore et al., 2012; Turnock et al., 2018); it is projected to increase under RCP8.5 and decrease under other RCPs depending largely on different methane emission trajectories because methane is an important precursor of ozone. Methane, therefore, reduces crop yield both from climate warming and ozone increase (Avnery et al., 2013). Shindell (2016) estimated yield losses of four major crops (to be 25±11% by 2100 under RCP8.5, as a net balance of the positive effect of CO2 (15±2%) and negative effects of warming (35±10%) and ozone (4.0±1.3%), and that 62% of the yield loss was attributable to methane. This points to the importance of reducing methane and other precursors of ozone as an effective adaptation strategy (medium evidence, high agreement ).
5.4.3.3 Projected impacts on other crops
Yield projections for crops other than cereals indicate mostly negative impacts on production due to a range of climate drivers (high confidence), with yield reductions similar to that of cereals expected in tropical, subtropical and semi-arid areas (Mbow et al., 2019). Springmann et al. (2016), compared the projected global food availability for different food groups under the SSP2 2050 scenario and found reductions in availability were similar in cereals, fruit and vegetables, and root and tubers (with legumes and oilseed crops showing a smaller reduction).
Fruit and vegetables have not been subject to extensive or coordinated yield projections (Figure 5.8). Yield projections have been performed for individual crops and locations (Ruane, 2014; Adhikari et al., 2015; Awoye et al., 2017; Ramachandran et al., 2017), but more often crop suitability models have been used (SM5.3). Zhao (2019) introduced a modelling approach that could be used to generate yield projections for a wider range of annual crops. The discussion here also draws on reviews of more restricted experimental studies. Negative impacts of climate change on crop production are expected across many cropping systems (Figure 5.8). Apart from the direct effects of elevated carbon dioxide, most changes are expected to have negative effects on crop production. Changes in temperature and rainfall are most often mentioned as drivers of climate impacts, but expected changes in phenology, pests and diseases are also raising concerns. Scheelbeek et al. (2018) synthesised projections for vegetables and legumes, based on their response to climate factors under experimental conditions; in most cases, the magnitude of the changes is comparable to the RCP8.5 2100 forecasts. Scheelbeek et al. (2018) projected yield changes of: +22.0% (+11.6% to +32.5%) for a 250 ppm increase in CO2 concentration; −34.7% (−44.6% to −24.9%) for a 50% reduction in water availability; −8.9% (−15.6% to −2.2%) for a 25% increase in ozone concentration; −31.5% for a 4°C increase in temperature (in papers with a baseline temperature of >20°C). Overall, impacts are expected to be largely negative in regions where the temperature is currently above 20°C, while some yield gains are expected in cooler regions (provided that water availability and other conditions are maintained). Scheelbeek et al. (2018) did not consider changes in pest and disease pressure, which are projected to increase with warming (see SM5.3).
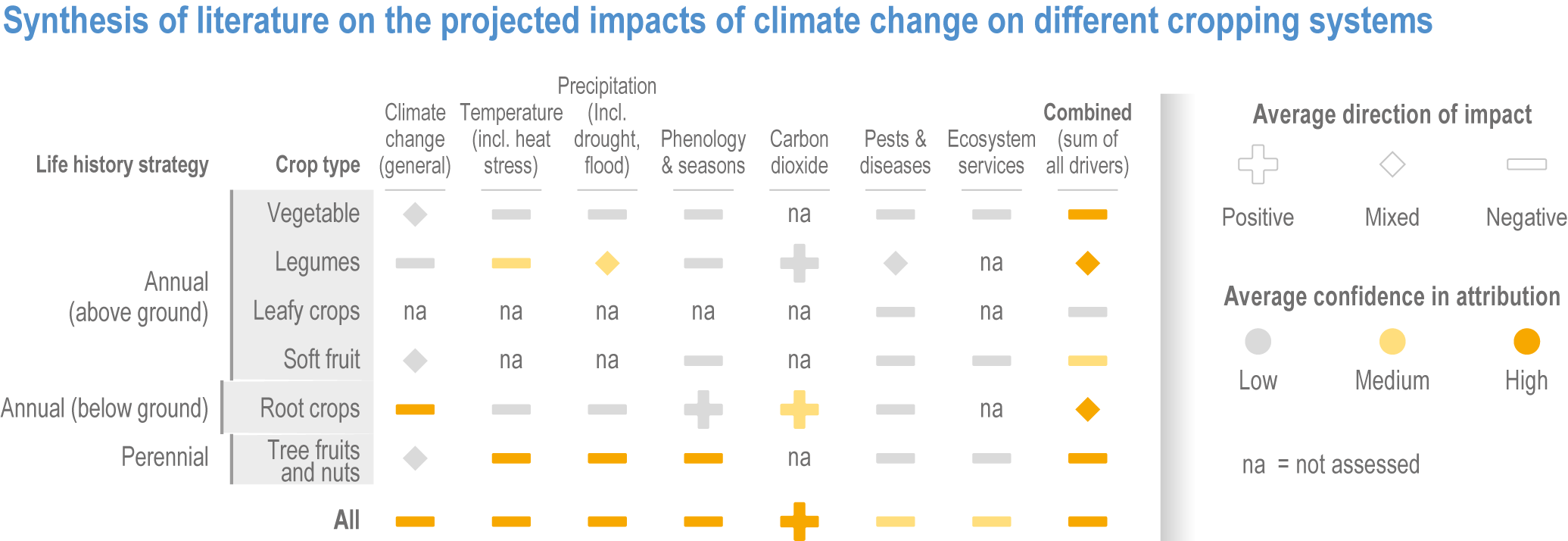
Figure 5.8 | Synthesis of literature on the projected impacts of climate change on different cropping systems. The assessment includes projections of impacts on crop productivity over a range of emission scenarios and time periods. The projected impacts are disaggregated by the different climate and climate-related drivers. Impacts are reported as positive, negative or mixed. The assessment draws on >60 articles published since AR5. The confidence is based on the evidence given in individual articles and on the number of articles. See SM5.2 information for details.
Systematic assessments of climate response for root crops as a group are lacking (Raymundo et al., 2014; Knox et al., 2016; Manners and van Etten, 2018). Climate suitability is projected to increase for tropical root crops (SM5.3), and some studies have found that root crops will be less negatively impacted than cereals, but there is no consensus on this (Brassard and Singh, 2008; Adhikari et al., 2015; Schafleitner, 2016; Manners et al., 2021). For potato, Raymundo et al. (2018) projected global yield reductions of 2–6% by 2055 under different RCPs, but with important differences among regions; tuber dry weight may experience reductions of 50–100% in marginal growing areas such as central Asia, while increases of up to 25% are expected in many high-yielding environments. Projections show yield increases of 6% per 100 ppm elevation in CO2 but declines of 4.6% per degree Celsius and 2% per 10% decrease in rainfall (Fleisher et al., 2017). Jennings et al. (2020) projected an overall increase in global potato production, but only if widespread adoption of adaptation measures is achieved. Although increases in CO2 could produce positive yield responses, the effects of temperature may offset these potential benefits (Dua et al., 2013; Raymundo et al., 2014). Warming offers the potential of longer growing seasons but can also have negative impacts through disrupted phenology and interactions with pests (Figure 5.8, Bebber, 2015; Pulatov et al., 2015).
Global yield modelling is lacking for woody perennial crops. Experimental studies suggest negative impacts on yields due to reduced water supply and increased soil salinity, as well as from warming and ozone (although evidence was limited for these) (Alae-Carew et al., 2020). Increasing CO2 is expected to increase yields, but only where other factors, such as warming, do not become yield-limiting (Alae-Carew et al., 2020). Many local projections include large uncertainty because of a lack of observational data and reliable parametrisation (Moriondo et al., 2015; Mosedale et al., 2016; Kerr et al., 2018; Mayer et al., 2019b). Most perennial crop models have found large negative impacts on yield and suitability, although CO2 fertilisation and phenology are not always considered (Lobell and Field, 2011; Glenn et al., 2013). Perennial crops are often grown in dryland areas where rainfall or irrigation water can be critical (Mrabet et al., 2020). Valverde (2015) found that yield losses in the Mediterranean region were largely driven by reduced rainfall, with maximum estimated yield losses of 5.4% for grape, 14.9% for olive and 27.2% for almond under a relatively hot and dry scenario (by 2041–2070). Moriondo (2015) highlight the need for perennial crop models to incorporate phenology and extreme climate events. Equally challenging is the need to estimate the impact of biotic changes, particularly climate-driven movement of pests and diseases (Ponti et al., 2014; Bosso et al., 2016; Schulze-Sylvester and Reineke, 2019).
For cotton, experimental studies suggest positive impacts from rising CO2 and temperature (Zhang et al., 2017a; Jans et al., 2021), but projections show mixed impacts on yield, including large negative impacts in warmer regions due to heat, drought and the interaction of temperature with phenology (Yang et al., 2014; Williams et al., 2015; Adhikari et al., 2016; Rahman et al., 2018). Climate change is also expected to increase the demand for irrigation water, which will likely limit production (Jans et al., 2021). There are also concerns that fibre quality may deteriorate (e.g., air permeability of compressed cotton fibers) (Luo et al., 2016).
Higher temperatures and altered moisture levels are expected to present a food safety risk, particularly for above-ground harvested vegetables (Figures 5.8; 5.10). Warmer and wetter weather is anticipated to increase fungal and microbial growth on leaves and fruit, while altered flooding regimes increase the risk of crop contamination (Liu et al., 2013; Uyttendaele et al., 2015). This is also true for perennial crops; for example, warming and climate variability can increase fungal contamination of grapes, including that associated with mycotoxins (Battilani, 2016; Paterson, 2018).
5.4.3.4 Observed and projected impacts on cultural ecosystem service
Cultural ecosystem services (CES) are those non-material benefits, such as aesthetic experiences, recreation, spiritual enrichment, social relations, cultural identity, knowledge and other values (Millennium Ecosystem Assessment, 2005), which support physical and mental health and human well-being (Chan et al., 2012; Triguero-Mas et al., 2015). CES in agricultural and wild landscapes include recreational activities, access to wild or cultivated products, and cultural foods, spiritual rituals, heritage and memory dimensions, and aesthetic experiences (Daugstad et al., 2006; Calvet-Mir et al., 2012; Ruoso et al., 2015). Relative to other ecosystem services, CES in agricultural landscapes have been less researched (Merlín-Uribe et al., 2012; Milcu et al., 2013; Bernues et al., 2014; Plieninger et al., 2014; van Berkel and Verburg, 2014; Ruoso et al., 2015; Quintas-Soriano et al., 2016). Agricultural heritage is a key aspect of CES and plays an important role in maintaining agrobiodiversity (Hanaček and Rodríguez-Labajos, 2018).
Climate change is projected to have negative impacts on CES (medium confidence) (Table 5.4). There is limited evidence that climate change has been the main driver affecting CES of agroecosystems confounded by other drivers such as migration and changing farming patterns (Hanaček and Rodríguez-Labajos, 2018; Dhakal and Kattel, 2019). Recent studies observed declines in CES in alpine pastures and floodplains in Europe in part due to climate change impacts (Probstl-Haider et al., 2016; Schirpke et al., 2019). Another study estimated that the scenic beauty enjoyed by those who visit the vineyards in central Chile will decline by 18–28% by 2050 owing to a combination of reduced precipitation, increased temperatures and natural fire cycles (Martinez-Harms et al., 2017). More research is needed, however, particularly on cultural heritage and spiritually significant places and in low-income countries.
Table 5.4 | Projected impacts on CES from climate change.
Region | CES | Climate change scenario | Projected impacts from climate change | References |
Central Chile, South America | Aesthetic experience of scenic beauty in vine-growing region. | RCP2.6 and 8.5. | Increased temperature, reduced precipitation and increased fires will damage scenic beauty of vineyards. Participatory scenario analysis estimated reduction in aesthetic experience from scenic beauty by 18–28% by 2050 for RCP2.6, with greater impacts under RCP8.5. | |
Mountainous regions of Austria | Cultural and aesthetic experiences in alpine pastures and diverse agricultural landscapes. | Temperature +1.5°C from 2008 to 2040 and four precipitation scenarios (high, similar , seasonal shift and low). | Some decline in CES, with trade-offs between diversity and CES and provisioning services depending upon the scenario. | |
Forest and agricultural landscapes in southern Saxony-Anhalt in Germany | Recreation, scenic landscape beauty and spiritual value of agricultural landscapes and forests. | Regional scenarios, do not specify RCPs. | Not anticipated to be significantly changed by climate change under most scenarios, except for intensification scenario, which would lead to a decline in the forest cultural services as they provide important historical and cultural ties. | Gorn et al. (2018) |
Northeast Austria floodplains (grasslands and wetlands) | Tourism, recreation, cultural heritage. | Increased temperature by 2050 and 2100 and seasonal shifts in precipitation. | Increased agricultural intensification due to shifts in climate and decline in CES is predicted, based on farmer interviews. | |
Mount Kenya, Kenya | Tourism, recreation, spiritual and cultural values. | Not specified. | Glacier disappearance may lead to reduced mountain trekking and other tourism and recreational activities. | |
Philippines | Nature-based tourism in agri-tourism. | Not specified. | Risk of typhoon, drought and strong wind, grass fire, heavy rains. Anticipated to increase vulnerability in terms of human health services and energy use in tourism. |
Box 5.2: Case Study: Wine
Wine-growing regions cover 7.4 million ha, with a value of 35 billion USD in 2018 (OIV, 2019). Important regions (Italy, France, Spain, USA, Argentina, Australia, South Africa, Chile, Germany, China, Argentina) are located in areas where mean annual temperature roughly varies between 10°C and 20°C (Schultz and Jones, 2010; Mosedale et al., 2016).
Temperature is the primary determinant for vine development. Recent warming trends have advanced flowering, maturity and harvest (high confidence) (Koufos et al., 2014; Cook and Wolkovich, 2016; Hall et al., 2016; Ruml et al., 2016; van Leeuwen and Destrac-Irvine, 2017; Koufos et al., 2020; Wang et al., 2020b; Wang and Li, 2020), and wine-growing regions have expanded outside the normal temperature bounds of locally grown varieties (limited evidence, high agreement ) (Kryza et al., 2015; Irimia et al., 2018). Milder winters have affected harvest in ice-wine growing regions (Pickering et al., 2015). Higher temperatures have mixed effects depending on site, but generally decrease grape quality (Barnuud et al., 2014; Morales et al., 2014; Sweetman et al., 2014; Kizildeniz et al., 2015; Kizildeniz et al., 2018). Warming increases sugar accumulation and decreases acidity (Leolini et al., 2019). Secondary metabolites are negatively affected (Biasi et al., 2019; Teslić et al., 2019). Developmental phases are projected to proceed faster in response to warming (high confidence) (Fraga et al., 2016a; Fraga et al., 2016b; García de Cortázar-Atauri et al., 2017; Costa et al., 2019; Molitor and Junk, 2019; Sánchez, 2019). However extreme high temperatures may have inhibitory effects on development (Cuccia et al., 2014).
In some cases, irrigation is required, and more frequent droughts are a key concern for yield and fruit quality (Morales et al., 2014; Bonada et al., 2015; Kizildeniz et al., 2015; Salazar-Parra, 2015; Kizildeniz et al., 2018; Funes et al., 2020). Water stress reduces shoot growth and berry size, and increases tannin and anthocyanin content (van Leeuwen and Darriet, 2016). However, controlled water stress produces positive impacts on wine quality, increasing skin phenolic compounds (van Leeuwen and Destrac-Irvine, 2017). The level of stress will depend on soil type, texture and organic matter content (Fraga et al., 2016a; Fraga et al., 2016b; Bonfante, 2017; García de Cortázar-Atauri et al., 2017; Leibar et al., 2017; Costa et al., 2019; Molitor and Junk, 2019; Sánchez, 2019). Increases in water demands with potential negative effects from increased soil salinity are among the most common effects of climate change in irrigated regions (medium evidence, high agreement ) (Mirás-Avalos et al., 2018; Phogat et al., 2018).
Rising CO2 will have mixed effects on vine growth and quality (medium evidence, high agreement ) (Martínez-Lüscher et al., 2016; Edwards et al., 2017; van Leeuwen and Destrac-Irvine, 2017). Rising CO2 concentrations will negatively affect wine quality by reducing anthocyanin concentration and colour intensity (Leibar et al., 2017).
Suitability responses to warming are region-specific. In regions where low temperature is a limiting factor, warming will enable growers to grow a wider range of varieties and obtain better-quality wines (high confidence) (Fuhrer et al., 2014; Mosedale et al., 2015; Mosedale et al., 2016; Meier et al., 2018; Jobin Poirier et al., 2019; Maciejczak and Mikiciuk, 2019). Subtropical and Mediterranean regions will experience major declines in fruit quality for high-quality wines (high confidence) (Resco et al., 2016; Lazoglou et al., 2018; Cardell et al., 2019; Fraga et al., 2019a; Fraga et al., 2019b; Teslić et al., 2019). These changes will also affect wine tourism (Nunes and Loureiro, 2016).
Impacts on suitability may reshape the geographical distribution of wine regions. Viability of the wine-growing regions will depend on the knowledge of local climatic variability (Neethling et al., 2019; Rességuier et al., 2020) and the implementation of adaptation strategies such as use of adapted plant material rootstocks, cultivars and clones, viticultural techniques (e.g., changing trunk height, leaf area to fruit weight ratio, timing of pruning), irrigation, enological interventions to control alcohol and acidity, and policy incentives and support (Callen et al., 2016; Ollat and Leeuwen, 2016; van Leeuwen and Destrac-Irvine, 2017; Merloni et al., 2018; Alikadic et al., 2019; del Pozo et al., 2019; Fraga et al., 2019b; Santillan et al., 2019; Morales-Castilla et al., 2020; Marín et al., 2021).
Box 5.3: Pollinators
Climate change will reduce the effectiveness of pollinator agents as species are lost from certain areas, or the coordination of pollinator activity and flower receptiveness is disrupted in some regions (high confidence) (Potts et al., 2010; Gonzalez-Varo et al., 2013; Polce et al., 2014; Kerr et al., 2015; Potts et al., 2016; Settele et al., 2016; Giannini et al., 2017; Mbow et al., 2019). A modelling study estimates that complete removal of pollinators could reduce global fruit supply by 23%, vegetables by 16%, and nuts and seeds by 22%, leading to significant increases in nutrient-deficient population and malnutrition-related diseases (Smith and Haddad, 2015), highlighting the importance of this ecosystem service for human health.
Bees are an essential agricultural pollinator, widely recognised for their role in the fertilisation of many domesticated plants. The observed widespread decline in native bees and honeybee colony numbers, particularly in the USA and Europe, has been associated with a number of environmental stressors in addition to climate change, such as neonicotinoids and varroa mites, and has raised concerns regarding plant–pollinator networks, the stability of pollination services, global food production and the prevalence of malnutrition (Williams and Osborne, 2009; Potts et al., 2010; Chaplin-Kramer et al., 2014).
Any climatic influence on floral phenology or physiology could, potentially, alter bee biology. At present, there is evidence that climate-change-induced asynchrony in pollen and pollinators can occur (Stemkovski et al., 2020). In addition, the nutritional composition of floral pollen may also affect bees’ health at the global level (low evidence). For example, goldenrod (Solidago spp.), a ubiquitous pollen source for bees just prior to winter, has experienced a ~30% drop in protein since the onset of CO2 emissions from the industrial revolution (Ziska et al., 2016).
Climate extremes could pose risks to pollinators when species tolerance is exceeded, with subsequent reduction in populations and potential extirpation (Nicholson and Egan, 2020; Soroye et al., 2020). The rate of climate change may induce potential mismatches in the timing of flowering and pollinator activity depending on the species (Bartomeus et al., 2011). For instance, Miller-Struttmann (2015) showed that long-tongued bumblebees may be at a disadvantage as warming temperatures are reducing their floral hosts, making generalist bumblebees more successful.
Overall, there is medium confidence that long-term mutualisms may be impacted directly by CO2 increases in terms of nutrition, or by temperature and other climatic shifts that may alter floral emergence relative to pollinator life cycles. Additional research is needed to further our understanding of the biological basis for these effects, and their consequence for pollination services.
Box 5.3
Box 5.4: Soil Health
Soil health, defined as an integrative property that reflects the capacity of soil to respond to land management, continues to support provisioning ecosystem services (Kibblewhite et al., 2008). Climate change will have significant impacts on soil health indicators such as soil organic matter (SOM). For example, precipitation extremes can reduce soil biological functions, and increase surface flooding, waterlogging, soil erosion and susceptibility to salinisation (Herbert et al., 2015; Chen and Mueller, 2018; Akter et al., 2019; Sánchez-Rodríguez et al., 2019).
The most significant threat to soil health is the loss of SOM (FAO and ITPS, 2015). SOM holds a great proportion of the nutrients, and regulates important soil physical, chemical and biological processes, such as cation exchange capacity, pH buffering, soil structure, water-holding capacity and microbial activity (FAO and ITPS, 2015). Soils also hold the largest terrestrial organic carbon stock, three to four times greater than the atmosphere (Stoorvogel et al., 2017). At the global scale, climate and vegetation are the main drivers of soil organic carbon (SOC) storage (Wiesmeier et al., 2019). While organic matter input is the primary driver of SOC stocks (Fujisaki et al., 2018), temperature and soil moisture play a key role in SOC storage at the local scale (Carvalhais et al., 2014; Doetterl et al., 2015). Soil type, land use and management practices also play important roles at the local scale.
Increase in soil temperature will negatively impact SOC, but primarily in higher latitudes (medium confidence) (Carey et al., 2016; Qi et al., 2016; Feng et al., 2017; Gregorich et al., 2017; Hicks Pries et al., 2017; Melillo et al., 2017; Hicks Pries et al., 2018). Experiments have shown that warming can accelerate litter mass loss and soil respiration (Lu et al., 2013) and reduces the soil recalcitrant C pool (Chen et al., 2020). SOC losses may speed up soil structural degradation, changes in soil stoichiometry and function (Hakkenberg et al., 2008; Tamene et al., 2019), with downstream effects on aquatic ecosystems. The rate and extent of SOC losses vary greatly depending on the scale of measurement (local to global), soil properties, climate, land use and management practices (Sanderman et al., 2017; Wiesmeier et al., 2019).
Adoption of practices that build SOC can improve crop resilience to climate-change-related stresses such as agricultural drought. Iizumi and Wagai (2019) found that a relatively small increase in topsoil (0–30 cm) SOC could reduce drought damages to crops over 70% of the global harvested area. The effects of increasing SOC are more positive in drylands owing to more efficient use of rainwater, which can increase drought tolerance (Iizumi and Wagai, 2019). Similarly, Sun et al. (2020) found that, relative to local conventional tillage, conservation agriculture has a win-win outcome of enhanced C sequestration and increased crop yield in arid regions. However, the impact of no-till may be minimal if not supplemented with residue cover and cover crops. As such, this is a highly debated area where some authors argue that no-till has limited effect and the evidence outside drylands is weak. Furthermore, the use of crop residues is constrained by its alternative uses (e.g., fuel, livestock feed, etc.) in much of the developing world. Practices that build up SOC may encourage soil microbial populations, which in turn can increase yield stability under drought conditions (Prudent et al., 2020).
Soil C sequestration is an important strategy to improve crop and livestock production sustainably that could be applied at large scales and at a low cost, if there was adequate institutional support and labour, using agroforestry, conservation agriculture, mixed cropping and targeted application of fertilizer and compost (high confidence) (Paustian et al., 2016; Kongsager, 2018; Nath et al., 2018; Woolf et al., 2018; Corbeels et al., 2019; Kuyah et al., 2019; Corbeels et al., 2020; Muchane et al., 2020; Sun et al., 2020; Nath et al., 2021). For example, a widespread adoption of agroforestry, conservation agriculture, mixed cropping and balanced application of fertilizer and compost by India’s small landholders could increase annual C sequestration by 70–130 Tg CO2 e (Nath et al., 2018; Nath et al., 2021).
5.4.4 Adaptation Options
Adaptation strategies in crop production range from field and farm-level technical options such as crop management and cultivar/crop options to livelihood diversification and income protection such as index-based insurance. This section assesses crop management options for different crop types. Feasibility of adaptation options in various systems is addressed in Section 5.1.4.
5.4.4.1 Adaptation options for major crops
Crop management practices are the most commonly studied adaptation measures (Shaffril et al., 2018; Hansen et al., 2019a; Muchuru and Nhamo, 2019), but quantitative assessments are mostly limited to existing agronomic options such as changes in planting schedules, cultivars and irrigation (Beveridge et al., 2018a; Aggarwal et al., 2019). This section draws on the global data set used in Section 5.4.3.2 (Hasegawa et al., 2021b) to estimate adaptation potential, defined as the difference in simulated yields with and without adaptations. A caveat to the analysis is that the data set includes management options if the literature treats them as adaptation. They include intensification measures such as fertilizer and water management, not allowing for physical and economic feasibility.
The overall adaptation potential of existing farm management practices to reduce yield losses averaged 8% in mid-century and 11% in end-century (Figure 5.9), which is insufficient to offset the negative impacts from climate change, particularly in currently warmer regions (Section 5.4.3.2). Emission scenarios, crop species, regions and adaptation options do not show discernible differences. Combinations of two or more options do not necessarily have greater adaptation potential than a single option, though a fair comparison is difficult in the data set from independent studies. One regional study in West Africa found that currently promising management would no longer be effective under future climate, suggesting the need to evaluate effectiveness under projected climate change.
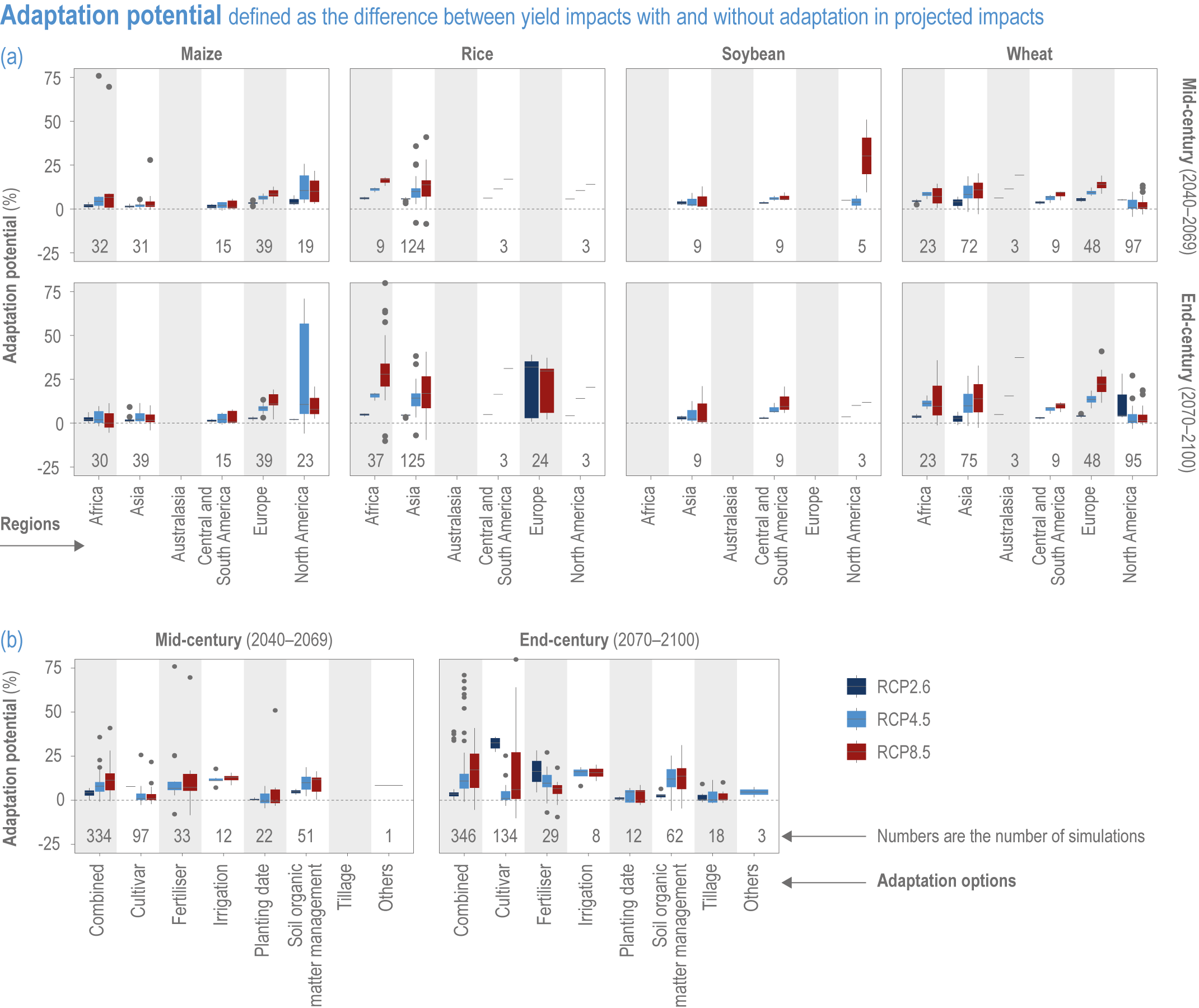
Figure 5.9 | Adaptation potential, defined as the difference between yield impacts with and without adaptation in projected impacts (Hasegawa et al. , 2021b). (a) Projections under three RCP scenarios by regions and (b) by options at mid-century (MC, 2040–2069) and end-century (EC, 2070–2100). n is the number of simulations. See Figure 5.6 for legends.
A global-scale meta-analysis estimated a 3–7% yield loss per degree Celsius increase in temperature (Zhao et al., 2017). Two global-scale studies using multiple global gridded crop models found that growing-season adaptation through cultivar changes offsets global production losses up to 2°C of temperature increase (Minoli et al., 2019; Zabel et al., 2021). While these studies do not account for CO2 fertilisation effects, another global-scale study with the CO2 fertilisation effects (Iizumi et al., 2020) showed that residual damage (climate change impacts after adaptation) would start to increase almost exponentially from 2040 towards the end of the century under RCP8.5. The cost required for adaptation and due to residual damage is projected to rise from USD 63 billion at 1.5°C to USD 80 billion at 2°C and to USD 128 billion at 3°C (Iizumi et al., 2020). All these global studies project that risks and damages are greater in tropical and arid regions, where crops are exposed to heat and drought stresses more often than in temperate regions (Sun et al., 2019; Kummu et al., 2021; SM5.4). There are still large uncertainties in the crop model projections (Müller et al., 2021a), but these multiple lines of evidence suggest that warming beyond +2°C (projected to be reached by mid-century under high-emission scenarios) will substantially increase the cost of adaptation and the residual damage to major crops (high confidence). The residual damage will prevail much sooner in currently warmer regions, where the effect of even a modest temperature increase is greater (Section 5.4.3.2).
Most crop modelling studies on adaptation are still limited to a handful of options for each crop type (Beveridge et al., 2018a). A range of other options are possible not just to reduce yield losses but to diversify risks to livelihoods, which are partially assessed in Sections 5.4.4.4 and 5.14.1. Current modelling approaches are not suited for the assessment of multiple dimensions of adaptation options. New studies are emerging that evaluate multiple options for productivity, sustainability and GHG emission (Xin and Tao, 2019; Smith et al., 2020b), but local- and household-scale assessment, taking account of future climatic variability, needs to be enhanced (Beveridge et al., 2018a).
5.4.4.2 Adaptation options for other crops
Across this diverse group of cropping systems, distinct adaptation options and adaptation limits have emerged (Figure 5.10; Acevedo et al., 2020; Berrang-Ford et al., 2021b). Some crop types have already seen widescale implementation of climate adaptation (e.g., grapevines), while others show little evidence of preparation for climate change (e.g., leafy salad crops). Many adaptation responses are shared with the major crops, but prominent options such as plant breeding are underutilised and there is a lack of evidence for assessing adaptation for many crops (Bisbis et al., 2018; Gunathilaka et al., 2018; Manners and van Etten, 2018). Figure 5.10 assesses several adaptation options based on the perceived importance of each in the literature. Fruit and vegetable crops tend to be more reliant on ecosystem services in the form of pollination, biocontrol and other resources (water, nutrients, microbes, etc.), and ecosystem-based adaptation options are prominent. The range of crops means that there is great potential for crop switching, but cultural and economic barriers will make such options difficult to implement, with barriers to entry for production and marketing (Waha et al., 2013; Magrini et al., 2016; Kongsager, 2017; Rhiney et al., 2018). Perennial crops are exposed to a wide range of climate factors throughout the year and have significant barriers to implementing some of the common adaptation options, such as relocation or replacing tree species/cultivar; agronomic interventions on-farm are well used in high-value tree crops and provide some climate resilience, but longer-term options will be needed (Glenn et al., 2013; Mosedale et al., 2016; Gunathilaka et al., 2018; Sugiura, 2019).
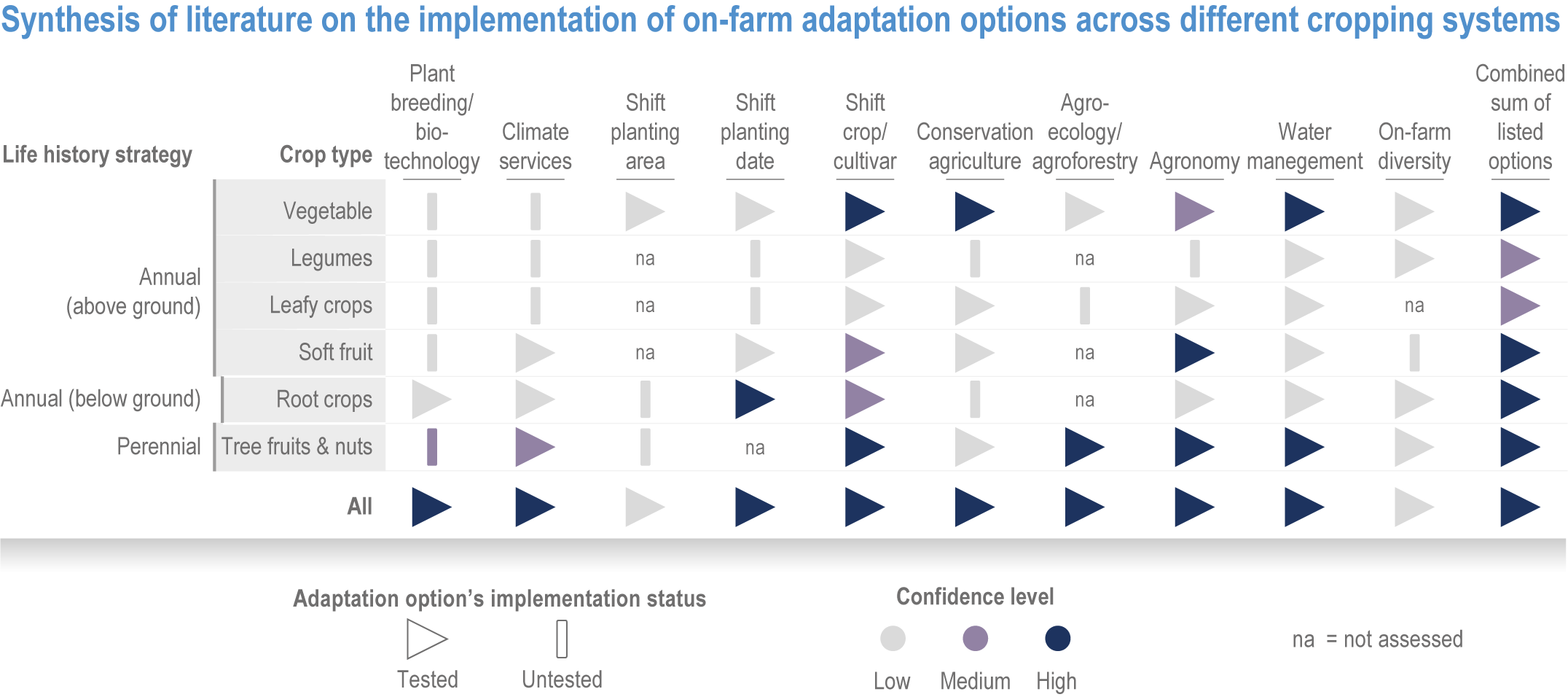
Figure 5.10 | Synthesis of literature on the implementation of on-farm adaptation options across different cropping systems. Adaptation options that have been implemented by growers are considered ‘tested’, while those that have not are considered ‘untested’. Untested options are those that appear in studies as suggestions by stakeholder or experts but were not implemented within the study. The assessment draws on >200 articles published since AR5. The confidence is based on the evidence given in individual articles and on the number of articles. See SM5.2 for details.
Many fruit and vegetable crops are water demanding, and adaptation responses relating to water management and access to irrigation water are crucial. Rainwater storage and deficit irrigation techniques are frequently mentioned as adaptation options and can minimise the burden on off-farm water supplies (Bisbis et al., 2018; Acevedo et al., 2020).
5.4.4.3 Cultivar improvements
As stated in AR5, cultivar improvements are one effective countermeasure against climate change (Porter et al., 2014; Challinor et al., 2016; Atlin et al., 2017). Plant breeding biotechnology for climate change adaptation draws upon modern biotechnology and conventional breeding, with the latter often assisted by genomics and molecular markers. Plant breeding biotechnology will contribute to adaptation for large-scale producers (high confidence). However, in addition to inconsistencies in meeting farmer expectations, a variety of socioeconomic and political variables strongly influence, and limit, uptake of climate-resilient crops (Acevedo et al., 2020; Rhoné et al., 2020).
Genome sequencing significantly increases the rate and accuracy for identifying genes of agronomic traits that are relevant to climate change, including adaptation to stress from pests and disease, temperature and water extremes (high confidence) (Brozynska et al., 2016; Scheben et al., 2016; Voss-Fels and Snowdon, 2016). Access to this information where it is needed and in practical timeframes, as well as the expertise to use it, will limit the sharing of benefits by the most vulnerable groups and countries (high agreement , limited evidence) (Heinemann et al., 2018).
Genetic improvements for climate change adaptation using modern biotechnology have not reliably translated into the field (Hu and Xiong, 2014; Nuccio et al., 2018; Napier et al., 2019), but good progress has been made by conventional breeding. Desirable traits that adapt plants to environmental stress are inherited as a complex of genes, each of which makes a small contribution to the trait (Negin and Moshelion, 2017). Adaptation by conventional breeding requires making rapid incremental changes in the best germplasm to keep pace with the environment (Millet et al., 2016; Atlin et al., 2017; Cobb et al., 2019). Further improvements would be difficult without in situ and ex situ conservation of plant genetic resources to maintain critical germplasm for breeding (Dempewolf et al., 2014; Castañeda-Álvarez et al., 2016).
Despite the advances in sequencing, phenotyping remains a significant bottleneck (Ghanem et al., 2015; Negin and Moshelion, 2017; Araus and Kefauver, 2018); the emergence of high-throughput phenotyping platforms may reduce this bottleneck in future. Emerging modern biotechnology such as gene/genome editing may in the future increase the ability to better translate genetic improvements into the field (medium agreement , limited evidence) (Puchta, 2017; Yamamoto et al., 2018; Friedrichs et al., 2019; Kawall, 2019; Zhang et al., 2019b).
Other breeding approaches assisted by genomics have been making steady gains in introducing traits that adapt crops to climate change (high confidence). DNA sequence information is used to identify markers of desirable traits that can be enriched in breeding programmes, as well as to quantify the genetic variability in species (Gepts, 2014; Brozynska et al., 2016; Voss-Fels and Snowdon, 2016). However, breeding for smallholder farmers and the stresses caused by climate change are unlikely to be addressed by the private sector and will require more public investment and adjusting to the local social-ecological system (Glover, 2014; Heinemann et al., 2014; Acevedo et al., 2020). Modern biotechnology has not demonstrated the scale neutrality needed to serve smallholder-dominated agroecosystems, due to a combination of the kinds of traits and restrictions that come from the predominant intellectual property rights instruments used in their commercialisation, as well as the focus on a small number of major crop species (medium confidence) (Fischer, 2016; Montenegro de Wit et al., 2020).
Globally, there is a notable lack of programmes aimed specifically at breeding for climate resilience in fruits and vegetables, although there have been calls to begin this process (Kole et al., 2015). Breeding for climate resilience in vegetables has great potential given the range of crop species available. Tolerance to abiotic stress is reasonably advanced in pulses (Araújo et al., 2015; Varshney et al., 2018), but examples of translation to commercial cultivars are still limited (Varshney et al., 2018; Varshney et al., 2019). The infrastructure for germplasm collection, maintenance, testing and breeding lags behind that of major crops (partly because of the large number of species involved) (Keatinge et al., 2016; Atlin et al., 2017).
Participatory plant breeding (PPB) facilitates interaction between Indigenous and local knowledge systems and scientific research and can be an effective adaptation strategy in generating varieties well adapted to the socio-ecological context and climate hazards (high confidence) (Table 5.5, Westengen and Brysting, 2014; Humphries et al., 2015; Anderson et al., 2016; Migliorini et al., 2016; Leitão et al., 2019; Ceccarelli and Grando, 2020; Singh et al., 2020).
Table 5.5 | PPB as cultivar improvement adaptation method.
Region | Crop(s) used for breeding | Results |
West Africa | Sorghum and pearl millet |
|
South America (Andes) | Potato |
|
Asia (southwest China) | Maize |
|
5.4.4.4 Integrated approach to enhance agroecosystem resilience
Diversifying agricultural systems is an adaptation strategy that can strengthen resilience to climate change, with socioeconomic and environmental co-benefits, but trade-offs and benefits vary by socio-ecological context (high confidence) (Table 5.6, M’Kaibi et al., 2015; Bellon et al., 2016; Jones, 2017b; Schulte et al., 2017; Jarecki et al., 2018; Jones et al., 2018; Luna-Gonzalez and Sorensen, 2018; Sibhatu and Qaim, 2018; Renard and Tilman, 2019; Rosa-Schleich et al., 2019; Bozzola and Smale, 2020; Mulwa and Visser, 2020). Crop diversification alongside livestock, fish and other species can be applied at various scales in a range of systems, from rainfed or irrigated to urban and home gardens in multiple spatial and temporal arrangements such as mixed planting, intercrops, crop rotation, diversified management of field margins, agroforestry (Section 5.10.1.3) and integrated crop livestock systems (Section 5.10.1.1, Isbell et al., 2017; Kremen and Merenlender, 2018; Dainese et al., 2019; Rosa-Schleich et al., 2019; Hussain et al., 2020; Renwick et al., 2020; Tamburini et al., 2020; Snapp et al., 2021; see Section 5.1 4 and Cross-Chapter Box NATURAL in Chapter 2).
Table 5.6 | Agroecosystem diversification practices, climate change adaptation mechanisms, trade-offs, co-benefits and constraints to implementation.
Agroecosystem diversification practice and mechanism for climate change adaptation | Benefits, trade-offs and constraints to implementation with examples |
Crop diversification
|
|
Legume diversification can be effective for both mitigation and adaptation, by reducing use of nitrogen derived from fossil fuels, and meat consumption, and providing ecosystem services through nutrient cycling, increasing soil biological activity and erosion control (Snapp et al., 2019). |
|
Organic amendments, no/low tillage or crop residue retention may increase diversity in soil biological organisms, which might be important in building resilience to multiple stresses such as drought and pest pressure (Furze et al., 2017; Blundell et al., 2020; de Vries et al., 2020; Stefan et al., 2021; Yang et al., 2021). |
|
Livestock integration. Inclusion of legumes and other forage into crop rotation allows mixed crop and livestock operations to mitigate farm-level risk and ecosystem buffering. |
|
Traditional and locally adapted mixed cropping and agroforestry practices which include leguminous trees can improve soil fertility and microclimate (Sida et al., 2018; Amadu et al., 2020). | Benefits: resilience to extreme events such as hurricanes can be promoted by supporting ecosystem functions to mitigate impacts and accelerate recovery (high agreement , medium evidence) (Altieri et al., 2015; Simelton et al., 2015; Sida et al., 2018; Perfecto et al., 2019).
|
Diversification improves regulating and supporting ecosystem services such as pest control, soil fertility and health, pollination, nutrient cycling, water regulation and buffering of temperature extremes (high confidence) (Barral et al., 2015; Prieto et al., 2015; Tiemann et al., 2015; Schulte et al., 2017; Beillouin et al., 2019a; Dainese et al., 2019; Kuyah et al., 2019; Tamburini et al., 2020), which can in turn mediate yield stability and reduced risk of crop loss according to socio-ecological contexts and time since adoption (high confidence) (Prieto et al., 2015; Roesch-McNally et al., 2018; Sida et al., 2018; Williams et al., 2018; Birthal and Hazrana, 2019; Degani et al., 2019; Amadu et al., 2020; Bowles et al., 2020; Li et al., 2020; Sanford et al., 2021).
Agroecosystem diversification often has variable impacts depending on crop combination, agro-ecological zone and soil types, and rigorous assessments of adaptive gains with traditional and locally diversified systems and potential trade-offs still need to be conducted across socio-ecological contexts. The quantitative upstanding will assist in enhancing multiple benefits of diversification tailored for each condition (Table 5.6). Progress is also needed via breeding and/or agronomy to adapt underutilised as well as major food crops to diversified agroecosystems and optimise management of nutrients, pest and disease pressure and other socio-ecological constraints (Araújo et al., 2015; Foyer et al., 2016; Adams et al., 2018; Pang et al., 2018).
Managing for diversity and flexibility at multiple scales is central to developing adaptive capacity. Policies to support diversification include shifting subsidies towards diversified systems, public procurement for diverse foods for schools and other public institutions, investment in shorter value chains, lower insurance premiums and payments for ecosystem services that include diversification (Sorensen et al., 2015; Guerra et al., 2017; Nehring et al., 2017; Valencia et al., 2019). Integrated landscape approaches involving multiple stakeholders (Reed et al., 2016) including urban governments can support diversification at a regional scale through public and private sector investment in extension services, regional supply chains, agritourism and other incentives for diversified landscapes (Milder et al., 2014; Münke et al., 2015; Sorensen et al., 2015; Pérez-Marin et al., 2017; Caron et al., 2018; 5.14.1.5).
5.5 Livestock-Based Systems
Livestock systems may be classified as industrial (monogastric, ruminant), grassland-based in which crop-based agriculture is absent or minimal (pastoralism, agro-pastoralism), mixed rainfed combining mostly rainfed cropping with livestock, and mixed irrigated systems with a significant proportion of irrigated cropping interspersed with livestock. Livestock systems are located widely across all regions of the world, and animal-sourced food provides humans with 39% of their protein and 18% of their calorie intake (FAO, 2019 f). Some 400 million people depend on livestock for a substantial part of their livelihood (Robinson et al., 2011).
5.5.1 Observed Impacts
Climate change affects livestock productivity and production in many ways (Porter et al., 2014; Rojas-Downing et al., 2017). Evidence is accumulating that rising temperatures are increasing heat stress in domestic species and affecting productivity (high confidence) (Das et al., 2016b; Godde et al., 2021).
5.5.1.1 Pastoral systems
Many grassland-based livestock systems are vulnerable to climate change and increases in climate variability (high confidence) (Dasgupta et al., 2014; Sloat et al., 2018; Stanimirova et al., 2019). Decadal vegetation changes from warming and drying trends have been detected in North American grasslands, with implications for species composition, rangeland quality and economic viability of grazing livestock (Rondeau et al., 2018; Reeves et al., 2020). Feed quality in South Asian grasslands has been negatively affected, reducing food security (Rasul et al., 2019). Increased grassland degradation has been observed in parts of Inner Mongolia (Nandintsetseg et al., 2021). Changing seasonality, increasing frequency of drought and rising temperatures are affecting pastoral systems globally (high confidence). These and other drivers are reducing herd mobility, decreasing productivity, increasing incidence of vector borne diseases and parasites, and reducing access to water and feed (high agreement , medium evidence) (López-i-Gelats et al., 2016; Vidal-González and Nahhass, 2018; de Leeuw et al., 2020).
5.5.1.2 Livestock distribution and climate variability
There is limited evidence of observed distributional changes in livestock species due to climate changes. Asian buffalo and yak breeds in China over the past 50 years have shifted distribution partly because of increases in heat stress (Wu, 2015; Wu, 2016). Nepalese cattle numbers have declined, attributed to increases in the number of hot days (Koirala and Shrestha, 2017).
Climate variability has been identified as the primary cause of vegetation cover changes on the Tibetan Plateau since 2000 (Lehnert et al., 2016). Increasing inter-annual variability is a driver of farm extensification in Mediterranean dairy systems (Dono et al., 2016). In Australian rangelands (Godde et al., 2019) and dairy systems (Harrison et al., 2016; Harrison et al., 2017), increasing rainfall variability contributes more to stocking rate and profitability variability than changes in mean rainfall.
5.5.1.3 Diseases and disease vectors
Climate change is affecting the transmission of vector-borne diseases (Hutter et al., 2018; Semenza and Suk, 2018) and parasites (Rinaldi et al., 2015) in high latitudes (high confidence). Different processes link climate change and infectious diseases in domesticated livestock. Some show a positive association between temperature and range expansion of arthropod vectors that spread the bluetongue virus. Others show a contraction, such as tsetse flies that transmit trypanosome parasites of several livestock species. Positive associations have been found between temperature and the spread of pathogens such as anthrax, and droughts and ENSO weather patterns and Rift Valley fever outbreaks in East Africa (Bett et al., 2017). Observed range expansion of economically important tick disease vectors in North America (Sonenshine, 2018) and Africa (Nyangiwe et al., 2018) are presenting new public health threats to humans and livestock.
5.5.2 Assessing Vulnerabilities
5.5.2.1 Rising temperature and heat stress
Most domestic livestock have comfort zones in the range 10–30°C, depending on species and breed (Nardone et al., 2006). At higher temperatures, animals eat 3–5% less per additional degree of temperature, reducing their productivity and fertility. Heat stress suppresses the immune and endocrine system, enhancing susceptibility of the animal to disease (Das et al., 2016b). Recent stagnation in dairy production in West Africa and China may be associated with increased periods of high daily temperatures (low confidence) (Rahimi et al., 2020; Ranjitkar et al., 2020). Increases in the productive capacity of domestic animals can compromise thermal acclimation and plasticity, creating further loss. Escalating demand for livestock products in low-to-middle-income countries (LMICs) may necessitate considerable adaptation in the face of new thermal environments (medium confidence) (Collier and Gebremedhin, 2015; Theusme et al., 2021). Heat effects on productivity have been summarised for pigs (da Fonseca de Oliveira et al., 2019), sheep and goats (Sejian et al., 2018), and cattle (Herbut et al., 2019). The direct effects of higher temperatures on the smaller ruminants (sheep and goats) are relatively muted, compared with large ruminants; goats are better able to cope with multiple stressors than sheep (Sejian et al., 2018). Under SSP5-8.5 to mid-century, land suitability for livestock production will decrease because of increased heat stress prevalence in mid and lower latitudes (high confidence) (Thornton et al., 2021).
5.5.2.2 Livestock water needs
Livestock production may account for 30% of all water (blue, green and grey) used in agriculture (Mekonnen and Hoekstra, 2010) and can negatively affect water quality. Cropland feed production accounts for 38% of crop water consumption (Weindl et al., 2017). High-input livestock systems may consume more water than grazing or mixed systems, though water used per kg beef produced, for example, depends on country, context and system (Noya et al., 2019). In systems where feed production is rainfed, livestock and crop water productivity may be comparable (Haileslassie et al., 2009). Direct water consumption by livestock is <1–2% of global water consumption (Hejazi et al., 2014). Rising temperatures increase animal water needs, potentially affecting access of herders and livestock to drinking water sources (Flörke et al., 2018).
5.5.2.3 Rising temperatures and livestock disease
Climate change will have effects on future distribution, incidence and severity of climate-sensitive infectious diseases of livestock (high confidence) (Bett et al., 2017). In an assessment of climate sensitivity of European human and domestic animal infectious pathogens, 63% were sensitive to rainfall and temperature, and zoonotic pathogens were more climate-sensitive than human- or animal-only pathogens (McIntyre et al., 2017). Over the last 75 years, >220 emerging zoonotic diseases, some associated with domesticated livestock, have been identified, several of which may be affected by climate change, particularly vector-borne diseases (Vaillancourt and Ogden, 2016; see Cross-Chapter Box ILLNESS in Chapter 2). Walsh et al. (2018) identified both temperature and rainfall as influential factors in predicting increasing anthrax outbreaks in northern latitudes. Growing infectious disease burdens in domesticated animals may have wide-ranging impacts on the vulnerability of rural livestock producers in the future, particularly related to human health and projected increases in zoonoses (high confidence) (Bett et al., 2017; Heffernan, 2018; Rushton et al., 2018; Meade et al., 2019).
5.5.2.4 Livestock and socioeconomic vulnerability to climate change
There is limited evidence about the role of livestock in addressing socioeconomic vulnerability. Although agriculture in parts of North America has become more sensitive to climate over the last 50 years, livestock have helped to moderate this effect, being less sensitive to increasing temperatures than some specialised crop systems (Ortiz-Bobea et al., 2018). Increasing frequency and severity of droughts will affect the future economic viability of grassland-based livestock production in the North American Great Plains (Briske et al., 2021). Purchasing more forage and selling more livestock have reduced household vulnerability in semi-arid parts of China over the last 35 years (Bai et al., 2019). A greater focus on sheep production away from cropping has increased the resilience of farming systems in Western Australia in low-rainfall years, although with mixed environmental effects (Ghahramani and Bowran, 2018). More insights are needed as to where and how livestock can affect the vulnerability of farmers and pastoralists.
5.5.2.5 Effects of climate on the health and vulnerability of livestock keepers
Vulnerability to the health impacts of climate change will be shaped by existing burdens of ill health and is expected to be highest in poor and socioeconomically marginalised populations (high agreement , limited evidence) (Labbé et al., 2016). In addition to projected changes in infectious disease burdens, labour capacity in a warming climate is anticipated to decrease further, beyond the >5% drop estimated since 2000 (Watts et al., 2018). Loss of labour capacity may greatly increase the vulnerability of subsistence livestock keepers (high agreement , limited evidence).
5.5.2.6 Gender and other social inequities
Vulnerability to climate change depends on demography and social roles (Mbow et al., 2019). Gender inequities can act as a risk multiplier, with women being more vulnerable than men to climate-change-induced food insecurity and related risks (high confidence) (Cross-Chapter Box GENDER in Chapter 18). Women and men often have differential and unequal control over different productive assets and the benefits they provide, such as income from livestock (Ngigi et al., 2017; Musinguzi et al., 2018). Indigenous livestock keepers can be more vulnerable to climate change, partly due to ongoing processes of land fragmentation (Hobbs et al., 2008), historical land dispossession, discrimination and colonialisation, creating greater levels of poverty and marginalisation (Stephen, 2018). Adaptation actions may also be affected by gender and other social inequities (Balehey et al., 2018; Dressler et al., 2019). Men and women heads of household may access institutional support for adaptation in different ways (Assan et al., 2018). Further research is warranted to evaluate alternative gendered and equity-based approaches that can address differences in adaptive capacity within communities.
5.5.3 Projected Impacts
There is limited evidence on future impact of climate change on livestock production, particularly in LMICs (Rivera-Ferre et al., 2016).
5.5.3.1 Impacts on rangelands, feeds and forages
Uncertainties persist regarding estimates of net primary productivity (NPP) in grazing lands (Fetzel et al., 2017; Chen et al., 2018b), so estimation of climate change impacts on grasslands is challenging. Mean global annual NPP is projected to decline 10 gC m −2 yr −1 in 2050 under RCP8.5, although herbaceous NPP is projected to increase slightly (Boone et al., 2018; see Figure 5.11). Similar estimates were made by Havlik et al. (2014): large increases in projected NPP in higher northern latitudes (21% increase in the USA and Canada) and large declines in western Africa (−46%) and Australia (−17%). The cumulative effects of impacts on forage productivity globally are projected to result in 7–10% declines in livestock numbers by 2050 for warming of ~2°C, representing a loss of livestock assets ranging from USD 10 to 13 billion (Boone et al., 2018). Changes to African grassland productivity will have substantial, negative impacts on the livelihoods of >180 million people.
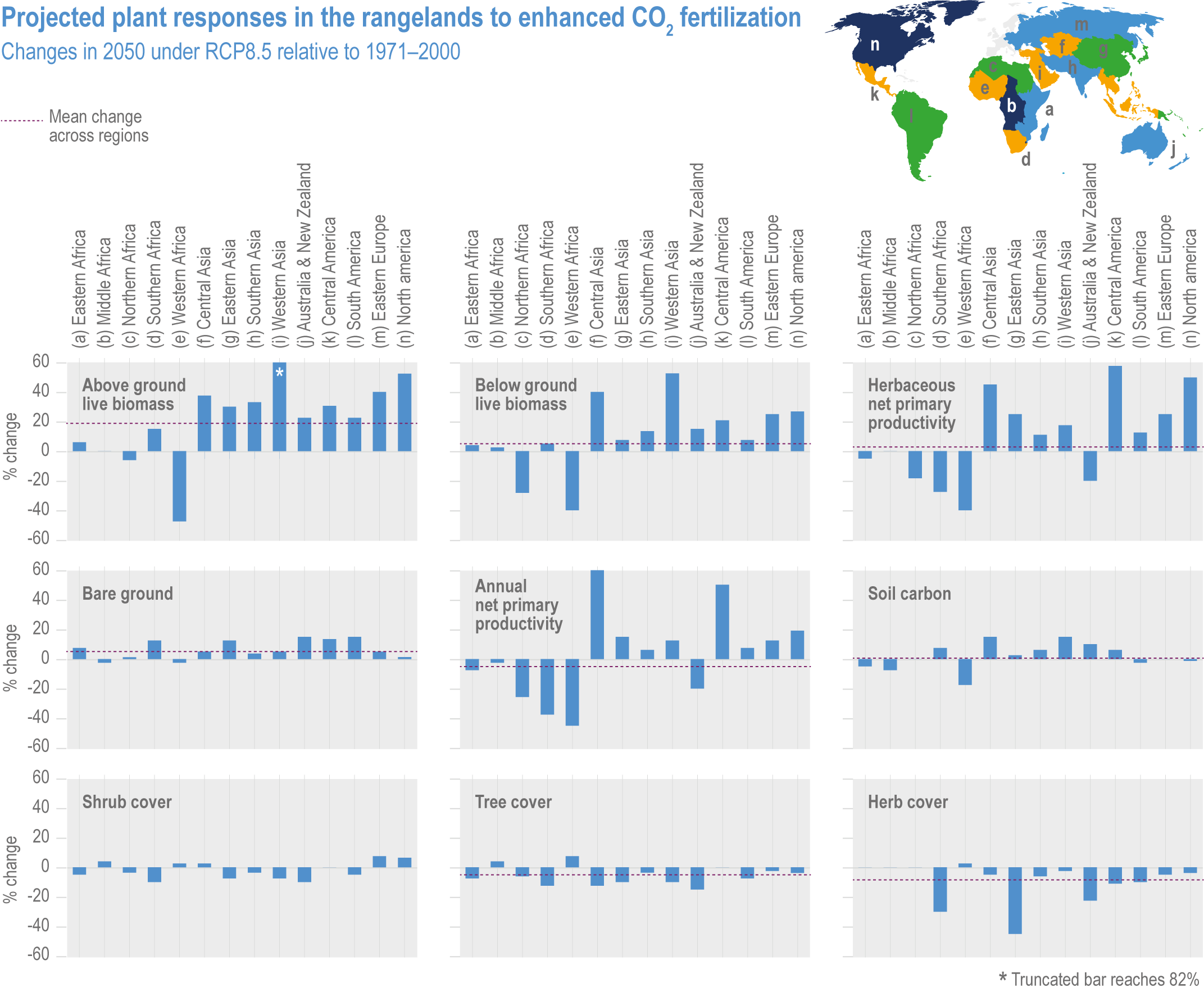
Figure 5.11 | Regional percent changes in land cover and soil carbon from ensemble simulation results in 2050 under emissions scenario RCP8. 5 compared with 1971–2000. Plant responses were enhanced by CO2 fertilisation. The larger chart (lower left) shows mean changes for all rangelands, and all charts are scaled to −60% to +60% change. Shown are annual net primary productivity (ANPP), herbaceous net primary productivity (HNPP), bare ground, herbaceous (herb), shrub, and tree cover, SOC (soil carbon), above-ground live biomass and below-ground live biomass. Regions as defined by the United Nations Statistics Division. The bar for above-ground live biomass in Western Asia (*) is truncated and is 82% (Boone et al., 2018).
Increases in above-ground NPP, and woody cover at the expense of grassland, are projected in some of the tropical and subtropical drylands (Doherty et al., 2010; Ravi et al., 2010; Saki et al., 2018), in Mediterranean wood pastures (Rolo and Moreno, 2019) and in the northern Great Plains of North America (Klemm et al., 2020). Godde et al. (2021) projected that woody encroachment would occur on 51% of global rangeland area by 2050 under RCP8.5. The future makeup of grasslands under climate change is uncertain, given the variation in responses of the component species, though this variation may provide a climate buffer (Jones, 2019) (low confidence). C4 grass species are regarded as less responsive to elevated carbon dioxide than C3 species, though this is not always the case (Reich et al., 2018).
There are other interactions between climate change and grazing effects on grasslands. Li (2018a) reported strong negative responses of NPP and species richness to 4°C warming, a 50% precipitation decrease, and high grazing intensity. Changes in grassland composition will inevitably change their suitability for different grazing animal species, with switches from herbaceous grazers such as cattle to goats and camels to take advantage of increases in shrubland (Kagunyu and Wanjohi, 2014). Rangeland feed quality may also be reduced via invasive species of lower quality than native species (Blumenthal et al., 2016).
Warming and water deficits impair the quality and digestibility of a C4 tropical forage grass, Panicum maximum, because of increases in leaf lignin (Habermann et al., 2019). A metanalysis by Dellar (2018) of climate change impacts on European pasture yield and quality found an increase in above-ground dry weight under increased CO2 concentrations for forbs, legumes, graminoids and shrubs with reductions in N concentrations in all plant functional groups. Temperature increases will increase yields in alpine and northern areas (+82.6%) but reduce N concentrations for shrubs (−13.6%) and forbs (−18.5%).
Increased temperatures and CO2 concentrations may increase herbaceous growth and favour legumes over grasses in mixed pastures (He et al., 2019). These effects may be modified by changes in rainfall patterns, plant competition, perennial growth habits and plant–animal interactions. The cumulative effect of these factors is uncertain. Large, persistent declines in forage quality are projected, irrespective of warming, under elevated CO2 conditions (600 ppm and +1.5°C day/3°C night temperature increases) in North American grasslands (Augustine et al., 2018). Rising CO2 concentrations may result in losses of iron, zinc and protein in plants by up to 8% by 2050 (Smith and Myers, 2018). Little information is available on possible impacts on carbon-based micronutrients, such as vitamins. About 57% of grasses globally are C3 plants and thus susceptible to CO2 effects on their nutritional quality (Osborne et al., 2014). These impacts will result in greater nutritional stress in grazing animals as well as reduced meat and milk production (quality and quantity) (high confidence, medium evidence).
5.5.3.2 Impacts of increased temperature on livestock
Recent research confirms the seriousness of the heat stress issue (medium evidence, high agreement ). Considerable increases are projected during this century in the number of ‘extreme stress’ days per year for cattle, chicken, goat, pig and sheep populations with SSP5-8.5 but many fewer with SSP1-2.6 (Thornton et al., 2021: Figure 5.12; see Cross-Chapter Box MOVING PLATE in this chapter). Resulting impacts on livestock production and productivity may be large, particularly for cattle throughout the tropics and subtropics and for goats in parts of Latin America and much of Africa and Asia. Pigs are projected to be particularly affected in the mid-latitudes of Europe, East Asia and North America. Lallo et al. (2018) estimated that global warming of 1.5°C and 2°C may exceed limits for normal thermo-regulation of livestock animals and result in persistent heat stress for animals in the Caribbean. Breed differences in heat stress resistance in dairy animals are now being quantified (Gantner et al., 2017), as are effects on sow reproductive performance in temperate climates (Wegner et al., 2016). Estimates of losses in milk production due to heat stress in parts of the USA, UK and West Africa to the end of the century range from 1% to 17% (Hristov et al., 2018; Fodor et al., 2018; Wreford and Topp, 2020; Rahimi et al., 2020). Much larger losses in dairy and beef production due to heat stress are projected for many parts of the tropics and subtropics: these could amount to USD 9 billion per year for dairy and USD 31 billion per for beef to end-century under SSP5-8.5, approximately 5% and 14% of the global value of production of these commodities in constant 2005 dollars.
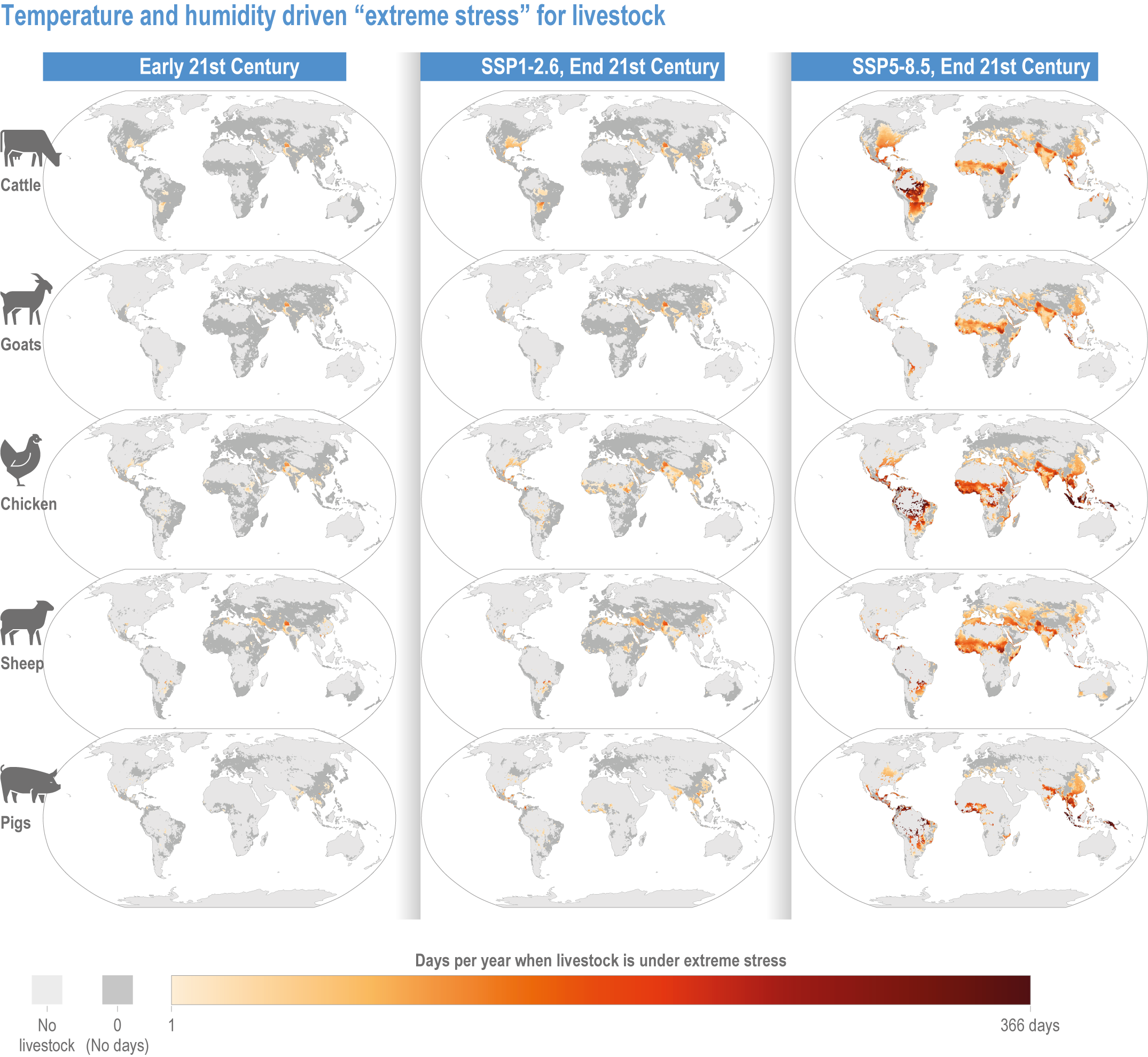
Figure 5.12 | Change in the number of days per year above ‘extreme stress’ values from the early 21st century (1991–2010) to end of century (2081–2100), estimated under SSP1-2. 6 and SSP5-8.5 using the Temperature Humidity Index (THI). Mapped for species current global distribution (Gilbert et al., 2018) (grey areas, no change). (Thornton et al., 2021), Also see Annex I: Global to Regional Atlas.
In many LMICs, poultry contribute significantly to rural livelihoods, including via modest improvements in nutritional outcomes of household children (de Bruyn et al., 2018). Rural poultry are generally assumed to be hardy and well adapted to stressful environments, but little information exists regarding their performance under warmer climates or interactions with other production challenges (Nyoni et al., 2019).
5.5.3.3 Impacts on livestock diseases
The impacts of climate change on livestock diseases remain highly uncertain (medium evidence, high agreement ). Bett et al. (2017) showed positive associations between rising temperature and expansion of the geographical ranges of arthropod vectors such asCulicoides imicola, which transmits the bluetongue virus. A 1-in-20-year bluetongue outbreak at present-day temperatures is projected to increase in frequency to 1-in-5 to 1-in-7 years by the 2050s, under RCP4.5 and RCP8.5, although animal movement restrictions can prevent devastating outbreaks (Jones et al., 2019).
The prevalence and occurrence of some livestock diseases are positively associated with extreme weather events (high confidence). There are high risks of future Rift Valley fever (RVF) outbreaks under both RCP4.5 and RCP8.5 this century in East Africa and beyond (Taylor et al., 2016; Mweya et al., 2017).
Few studies explicitly consider the biotic and abiotic factors that interact additively, multiplicatively or antagonistically to influence host–pathogen dynamics (Cable et al., 2017). Integrative concepts that aim to improve the health of people, animals and the environment such as One Health may offer a framework for enhancing understanding of these complex interactions (Zinsstag et al., 2018). Much remains unknown concerning disease transmission dynamics under a warming climate (Heffernan, 2018), highlighting the need for effective monitoring of livestock disease (Brito et al., 2017; Hristov et al., 2018).
5.5.3.4 Impacts on livestock and water resources
Water resources for livestock may decrease in places because of increased runoff and reduced groundwater resources, as well as decreased groundwater availability in some environments (AR5). Increased temperatures will cause changes in river flow and the amount of water stored in basins, potentially leading to increased water stress in dry areas such as parts of the Volta River Basin (Mul et al., 2015). Toure (2017) estimated decreases in groundwater recharge rates of 49% and of stored groundwater by 24% to the 2030s in the Klela Basin in Mali under both RCP4.5 and RCP8.5, with potentially serious consequences for water availability for livestock and irrigation.
Water intake by livestock is related to species, breed, animal size, age, diet, animal activity, temperature and physiological status of animals (Henry et al., 2018). Direct water use by cattle may increase by 13% for a temperature increase of 2.7°C in a subtropical region (Harle et al., 2007). Changes in water availability may arise because of decreased supply or increased competition from other sectors. Availability changes may be accompanied by shifts in water quality, such as increased levels of microorganisms and algae, that can negatively affect livestock health (Naqvi et al., 2015). In arid lands, projected decreases in water availability will severely compromise reproductive performance and productivity in sheep (Naqvi et al., 2017). In higher-input livestock systems, water costs may increase substantially owing to increased competition for water (Rivera-Ferre et al., 2016).
5.5.3.5 Livestock and climate variability
Information on future climate variability changes on livestock system productivity does not exist yet. Increases in climate variability may increase food insecurity in the future, mediated through increased crop and livestock production variability (Thornton and Herrero, 2014) in LMICs. Rainfall variability increases in pastoral lands have been linked to declining cattle numbers (Megersa et al., 2014). Changes in future climate variability may have large negative impacts on livestock system outcomes (Sloat et al., 2018; Stanimirova et al., 2019); these effects can be larger than those associated with gradual climate change (limited evidence, medium agreement ) (Godde et al., 2019). In grasslands, Chang et al. (2017) (Europe) and Godde et al. (2020) (globally) projected increases in biomass inter-annual variability, the worst effects occurring in rangeland communities that are already vulnerable. Ways in which climate variability impacts have been addressed in the past, such as via herd mobility, may become increasingly unviable in the future (Hobbs et al., 2008).
5.5.3.6 Societal impacts within the production system
Livestock play important social (Kitalyi et al., 2005) and cultural (Gandini and Villa, 2003) roles in many societies. Climate change will negatively affect the provisioning of social benefits in many of the world’s grasslands (medium confidence). Examples include moving to semi-private land ownership models, driven in part by climate change, that are changing social networks and limiting socio-ecological resilience in pastoral systems in East Africa (Kibet et al., 2016; Bruyere et al., 2018) and Asia (Cao et al., 2018a); altering traditional food, resource and medicine sharing mechanisms in West Africa (Boafo et al., 2016); and the limited ability of current livestock systems to satisfy societies’ demand for CES in Northwest Europe (Bengtsson et al., 2019). The societal impacts of climate change on livestock systems may interact with drivers of change and increase herders’ vulnerability via processes of sedentarisation and land fragmentation, both of which may result in decreased animal access to rangelands (Adhikari et al., 2015; Cross-Chapter Box MOVING PLATE this chapter). Stronger linkages are needed between ecosystem service and food security research and policy to address these challenges (Gentle and Thwaites, 2016; Bengtsson et al., 2019).
5.5.4 Adaptation in Livestock-Based Systems
Livestock adaptation options are increasingly being studied with methods such as agent-based household models (Hailegiorgis et al., 2018), household models that disaggregate climate scenarios as well as differentiate farms of varying types and farmer attributes (Descheemaeker et al., 2018), new meso-scale grassland models (Boone et al., 2018) and modelling approaches that capture decision making at the farm level for sample populations (Henderson et al., 2018).
Many grassland-based livestock systems have been highly resilient to past climate risk, providing a sound starting point for current and future climate change adaptation (Hobbs et al., 2008). These adaptations include more effective matching of stocking rates with pasture or other feed production; adjusting herd and watering point management to altered seasonal and spatial patterns of forage production; managing diet quality, which also helps reduce enteric fermentation in ruminants and thus GHG emissions (using diet supplements, legumes, choice of introduced pasture species and pasture fertility management); more effective use of silage, rotational grazing or other forms of pasture spelling; fire management to control woody thickening; using better-adapted livestock breeds and species; restoration of degraded pastureland; migratory pastoralist activities; and a wide range of biosecurity activities to monitor and manage the spread of pests, weeds and diseases (Herrero et al., 2015; Godde et al., 2020). Combining adaptations can result in increases in benefits in terms of production and livelihoods over and above those attainable from single adaptations (high confidence) (Bonaudo et al., 2014; Thornton and Herrero, 2015; ul Haq et al., 2021).
The adaptations that livestock keepers have been undertaking in Asia (Hussain et al., 2016; Li et al., 2017) and Africa (Belay et al., 2017; Ouédraogo et al., 2017) are largely driven by their perceptions of climate change. Keeping two or more species of livestock simultaneously on the same farm can confer economic and sustainability benefits to European farmers (Martin et al., 2020). Some livestock producers are changing and diversifying management practices, improving access to water sources, increasing uptake of off-farm activities, trading short-term profits for longer-term resilience benefits and migrating out of the area (Hussain et al., 2016; Berhe et al., 2017; Merrey et al., 2018; Thornton et al., 2018 ; Espeland et al., 2020). Others are adopting more climate-resilient livestock species such as camels (Watson et al., 2016a), using climate forecasts at differing time scales, and benefitting from innovative livestock insurance schemes, though challenges remain in their use at scale (Dayamba et al., 2018; Hansen et al., 2019a; Johnson et al., 2019).
In West Africa, cattle and small ruminant producers and traders are changing strategies in response to emerging market opportunities as well as to multiple challenges including climate change (Gautier et al., 2016; Ouédraogo et al., 2017). Niles (2017) found that reduced food insecurity in 12 countries was associated with livestock ownership, providing cash for food purchases. Livestock ownership or switching to smaller, local breeds does not automatically translate into positive nutrition outcomes for women and children, although it may if communities see such animals as suitable for husbandry by women (Chanamuto and Hall, 2015); the relationship is complex (Nyantakyi-Frimpong and Bezner-Kerr, 2015; Dumas et al., 2018).
Options for adapting domestic livestock systems to increased exposure to heat stress (Table 5.7) include breeding and crossbreeding strategies, species switching, low-cost shading alternatives and ventilation and building-design options (Chang-Fung-Martel et al., 2017; Godde et al., 2021). In utero exposure to heat stress may increase adaptive capacity in later life, though the underlying mechanisms are incompletely understood (Skibiel et al., 2018). For confined livestock systems in temperate regions, the economic consequences of adapting to heat stress are still being quantified.
Table 5.7 | Selected adaptations to heat stress in livestock systems.
Adaptation | Example | Reference |
Breeding for heat stress tolerance | Sheep and cattle farming systems in southern Australia under IPCC Special Report on Emissions Scenarios (SRES) A2. Projected not to improve livestock productivity by 2070, even in drier locations. | |
‘Slick hair’ breeding | In the Caribbean, introduction of a ‘slick hair’ gene into Holstein cows by crossbreeding with Senepols to increase thermo-tolerance and productivity. An integrated approach to heat stress adaptation will still be needed, including shading strategies, for example. | |
Crossbreeding | Crossbreeding with Indigenous sheep breeds as an adaptation option in Mongolia produced some benefits in productivity and improved adaptation to winter cold. Best combined with other improved management interventions. In general, effectiveness of crossbreeding as an adaptation strategy will be dependent on context. | |
Species switching | Switching from large ruminants to more heat-resilient goats for dairy production in Mediterranean systems to adapt to increasing heat stress. Switching from cattle to more heat- and drought-resilient camels in pastoral systems of southern Ethiopia as an adaptation to increasing drought. | Wako et al. (2017) |
Shading, fanning, bathing | Low-capital relief strategies (shading with trees or different types of shed; bathing animals several times each day; installing electric fans in sheds) are effective at reducing heat stress impacts on household income in smallholder dairy systems in India. Different tree arrangements in silvopastoral systems in Brazil were effective in reducing thermal loads by up to 22% for animals compared with full-sun pasture. | |
Ventilation and cooling systems | A wide range of different ventilation systems, cooling systems and building designs for confined and seasonally confined intensive livestock systems (pigs, poultry, beef, dairy) in temperate regions. Economic consequences and profitability of different options under different RCPs are still being assessed. | Hempel and Menz (2019), Mikovits et al. (2019), Schauberger et al. (2019b) |
In utero exposure to heat stress | Potential as an adaption option is uncertain, as there are different effects of in utero heat stress exposure and the mechanisms are not completely understood:
|
New research is investigating the prospects for accelerating traditional and novel breeding processes for animal traits that may be effective in improving livestock adaptation as well as production (Stranden et al., 2019; Barbato et al., 2020). Even if the technical challenges of using new tools such as CRISPR-Cas9 for genome editing in livestock are overcome, the granting of societal approval to operate in this research space may be elusive (Herrero et al., 2020; Menchaca et al., 2020).
5.5.4.1 Contributions of Indigenous knowledge and local knowledge
Indigenous knowledge has a role to play in helping livestock keepers adapt (medium confidence), though the transferability of this knowledge is often unclear. Pastoralists’ local knowledge of climate and ecological change can complement scientific research (Klein et al., 2014), and local knowledge can be mobilised to inform adaptation decision making (Klenk et al., 2017). While Indigenous weather forecasting systems among pastoralists in Ethiopia (Balehegn et al., 2019; Iticha and Husen, 2019) and Uganda (Nkuba et al., 2020) are effective, synergies can be gained by combining traditional and modern knowledge to help pastoralists adapt. Sophisticated knowledge of feed resources among agro-pastoralists in West Africa is being used to increase system resilience (Naah and Braun, 2019). Understanding local knowledge for adaptation can present research challenges, for which new multi-disciplinary research methods may be needed (Reyes-Garcia et al., 2016; Roncoli et al., 2016). In particular, the complexities of knowledge, practice, power, local governance and politics need to be addressed (Hopping et al., 2016; Scoville-Simonds et al., 2020).
Box 5.5: Alternative Sources of Protein for Food and Feed
Alternative protein sources for human food and livestock feed are receiving considerable attention. Laboratory or ‘clean meat’ is one potential contributor to the human demand for protein in the future (SRCLL). Such technology may be highly disruptive to existing value chains but could lead to significant reduction in land use for pastures and crop-based animal feeds (Burton, 2019; Rosenzweig et al., 2020). The impacts on GHG emissions depend on the meat being substituted and the trade-off between industrial energy consumption and agricultural land requirements (Mattick et al., 2015; Alexander et al., 2017; Rubio et al., 2020b; Santo et al., 2020). Livestock feeds can make use of other protein sources: insects are generally rich in protein and can be a significant source of vitamins and minerals. Black soldier fly, yellow mealworm and the common housefly have been identified for potential use in feed products in the EU, for example (Henchion et al., 2017). Replacing land-based crops in livestock diets with some proportion of insect-derived protein may reduce the GHG emissions associated with livestock production, though these and other potential effects have not yet been quantified (Parodi et al., 2018; Section 5.13.2). Other sources are high-protein woody plants such as paper mulberry (Du et al., 2021) and algae, including seaweed. While microalgae and cyanobacteria are mainly sold as a dietary supplement for human consumption, they are also used as a feed additive for livestock and aquaculture, being nutritionally comparable to vegetable proteins. The potential for cultivated seaweed as a feed supplement may be even greater: some red and green seaweeds are rich in highly digestible protein. Asparagopsis taxiformis, for example, also decreases methane production in both cattle and sheep when used as a feed supplement (Machado et al., 2016; Li et al., 2018b). Novel protein sources may have considerable potential for sustainably delivering protein for food and feed alike, though their nutritional, environmental, technological and socioeconomic impacts at scale need to be researched and evaluated further.
5.6 Forestry Systems
Forests play a vital role in the ecology of the planet, including climate regulation, and provide a range of important ecosystem services within their local landscape. Moreover, they are essential to the well-being of millions of people around the world. Forests are sources of food contributing about 0.6% of global food consumption and provide important products, such as timber and non-timber forest products (NTFPs) (FAO, 2014). Indigenous Peoples and local communities are estimated to manage at least 17% of total carbon (or 293 × 109Mg) stored in forest in 64 assessed countries (RRI, 2018a). While small in number, numerous local communities around the world are highly or entirely dependent on forests for their food supply (Karttunen et al., 2017). An estimated 9% of the world’s rural population is lifted above the extreme poverty line because of income from forest resources (World Bank, 2016). Additionally, forest income plays a particularly important role in diversifying the income sources of poor households, reducing their vulnerability to loss from one source of income. This section covers an assessment of the impacts of climate change on forestry production systems and the adaptation options available. Non-timber forest products will be covered in the next section.
5.6.1 Observed Impacts
The IPCC AR5 stated that there is high confidence that numerous plants and animal species have already migrated, changed their abundance, and shifted their seasonal activities as a result of climate change (Settele et al., 2014). The report highlighted the widespread deaths of trees in many forested areas of the world. Forest die back could significantly affect wood production among other impacts.
The SRCCL (Barbosa et al., 2019) concluded that climate change will have positive and negative effects on forests, with varying regional and temporal patterns. For example, the SRCCL noted the increasing productivity in high-latitude forests such as those in Siberia. In contrast, negative impacts are already being observed in other regions such as increasing tree mortality due to wildfires.
In the past years, tree mortality continued to increase in many parts of the world. Large pulses of tree mortality were consistently linked to warmer and drier than average conditions for forests throughout the temperate and boreal biomes (high confidence) (Sommerfeld et al., 2018; Seidl et al., 2020). Long-term monitoring of tropical forests indicates that climate change has begun to increase tree mortality and alter regeneration (Hubau et al., 2020; Sullivan et al., 2020). Climate-related die back has also been observed due to novel interactions between the life cycles of trees and pest species (Kurz et al., 2008; Lesk et al., 2017; Sambaraju et al., 2019). A recent example of the impacts of climatic extremes is the European drought of 2018 (Buras et al., 2020), which led to a significant browning of the vegetation and resulted in widespread tree mortality (high confidence) (Brun et al., 2020; Schuldt et al., 2020). This brought markets for conifer timber close to collapse in parts of Europe, posing considerable challenges for timber-based forestry and leading to cascading impacts on society (Hlásny et al., 2021). Overall, there is robust evidence and medium agreement that provisioning services of boreal and temperate forests are affected negatively by forest disturbances, while for cultural services only limited evidence with medium agreement exists (Thom and Seidl, 2016).
Increasingly, climate impacts on the recovery of forests after disturbance are observed: using data from the past 20 years and 33 wildfires, it has been shown that post-fire regeneration of Pinus ponderosa and Pseudotsuga menziesii in the western USA has declined because of climate change and increased severity of fires (Davis et al., 2019). However, the observed patterns of post-disturbance recovery vary with region, with reduced tree regeneration reported for the western USA (Stevens-Rumann and Morgan, 2019; Turner et al., 2019) but robust recovery observed in Canada (White et al., 2017) and Central Europe (medium confidence) (Senf et al., 2019).
Also, the distribution and traits of trees are increasingly influenced by climate change, with impacts for local ecosystem service supply. In the USA, a study of 86 tree species/groups over the past three decades showed that more tree species have shifted westward (73%) than poleward (62%) in their abundance (Fei et al., 2017). This was due more to changes in moisture availability than to changes in temperature. As climate has warmed, trees are growing faster with longer growing seasons. However, a study of forests in Central Europe revealed that wood density has decreased since the 1870s (Pretzsch et al., 2018). This means that increasing tree growth might not directly translate to increased total biomass and carbon sequestration.
5.6.2 Projected Impacts
AR5 stated that other stressors such as human-driven land use change and pollution will continue to be the main causes of forest cover change in the next three decades (Settele et al., 2014). In the second half of this century, it was projected that climate change will be a strong stressor of change in forest ecosystems. Many forest species may not be able to move fast enough to adjust to new climate conditions. In some cases, a warmer climate could lead to extinction of species.
The SR15 concluded that limiting warming to 1.5°C will be more favourable to terrestrial ecosystems, including forests, relative to a 2°C warming (Hoegh-Guldberg et al., 2018). In general, a 2°C warming could lead to two times more area of biome shifts compared with a 1.5°C warming. As a result, keeping a cooler average global temperature will lead to lower extinction risks. The special report supports the AR5 conclusion that a warmer planet will impact wide swaths of forests adversely. For example, higher temperatures will promote fire, drought and insect disturbances. Consistent with AR5, SRCCL projected that tree mortality will increase with climate change (Barbosa et al., 2019). In addition, forests will be more exposed to extreme events such as extreme heat, droughts and storms. The incidence of forest fires will likewise increase.
Additional evidence since the above reports were published supports their overall conclusions. For example, at the global scale, modelling the vulnerability of 387 forest ecoregions under future climate change (to 2080 using the average of five Global Circulation Models (GCMs) and RCP4.5 and 8.5) across different biomes, biogeographical realms and conservation statuses showed that 8.8% of global forest ecoregions are highly vulnerable in a low-greenhouse-gas-concentration scenario, and 32.6% of the global forest ecoregions are highly vulnerable in a high-greenhouse-gas-concentration scenario (Wang et al., 2019a). Furthermore, a recent synthesis of the literature suggests that climate change will result in younger and shorter forests globally (McDowell et al., 2020). In Asia, a systematic review of climate change impacts on tropical forests revealed that future climate may lead to changes in species distribution and forest structure and composition as well as phenology (Deb et al., 2018).
Overall, studies indicate both negative and positive climate change impacts on forest production systems. Some forests in the USA could benefit slightly from CO2 fertilisation (using IGSM-CAM and MIROC3.2 till 2100) resulting in increased productivity especially for hardwoods (Beach et al., 2015). A study across Europe showed that both productivity gains (mostly in Northern and Central Europe, up to +33%) and losses (predominately in Southern Europe, up to −37%) are possible until the end of the 21st century (Reyer et al., 2017). The study further indicated that disturbances would reduce gains and exacerbate losses of productivity throughout Europe under climate change (Reyer et al., 2017). For Central and Eastern Canada, decreasing biomass production is projected as a result of increasing disturbance from wildfire and drought (Brecka et al., 2020). Climate-induced disturbances could also reduce the temporal stability of ecosystem service supply (Albrich et al., 2018), increasing the volatility of timber markets (medium confidence). More broadly, climate change could lead to abrupt changes and the crossing of tipping points, resulting in profoundly altered future forest development trajectories (Turner et al., 2020). Some studies suggest that such threshold could already be crossed at relatively low warming levels of +2°C (Elkin et al., 2013; Albrich et al., 2020), with substantial implications for ecosystem service supply (limited evidence, high agreement ).
Regional studies on the potential future effects of climate change on forest production systems indicate diverse impacts. In Germany, drier conditions in 2070 (RCP8.5; GCMs INM-CM4, ECHAM6 and ACCESS1.0) are expected to benefit the mean annual increment at biological rotation age of Scots pine and oak, while beech might suffer losses of up to 3 m 3 ha –1 yr –1 depending on climate scenario and region (Albert et al., 2018). In India, 46% of the forest grid points were found to have high, very high or extremely high vulnerability under future climate in the short term (2030s) under both RCP4.5 and 8.5, increasing to 49% and 54%, respectively, in the long term (2080s) (Sharma et al., 2017). In addition, forests in the higher rainfall zones show lower vulnerability as compared with drier forests under future climate, which is in contrast to dry forests in Central and South America cited above. Warming and drying trends are projected to reduce timber production in the neotropics in some cases (Hiltner et al., 2021). Also in India, a study using CMIP5 (RCP4.5 and 8.5 with two time slices 2021–2050 and 2070–2099) shows how forests in five districts in Himachal Pradesh in Western Himalayan region are vulnerable to global warming (Upgupta et al., 2015). In the Guiana Shield, climate projections under RCP2.5 and 8.5 led to decreasing the basal area, above-ground fresh biomass, quadratic diameter, tree growth and mortality rates of tropical forests (Aubry-Kientz et al., 2019). In Central Africa, projections under RCP4.5 and 8.5 showed a general increase in growth, mortality and recruitment leading to a strong natural thinning effect, with different magnitudes across species (Claeys et al., 2019).
On a global and regional scale, there is limited evidence and high agreement (medium confidence) that climate change will increase global and regional supply of timber and other forest products. To date, there are eight studies assessing the total economic impacts of climate change on the forestry sector at the global level. Some of them have assumed only flow effects of climate change by using the projected changes in yields of forest types from integrated economic models (Perez-Garcia et al., 1997; Perez-Garcia et al., 2002; Buongiorno, 2015), while other studies have assumed both flow and stock effects by accounting for changes in forest yields, die back effects and biome migration (Sohngen et al., 2001; Lee and Lyon, 2004; Tian et al., 2016; Favero et al., 2018; Favero et al., 2021).
According to these studies, global timber supply will increase as the result of an increase in global forest growth under climate change scenarios (medium confidence). Some studies indicate that timber supply is projected to increase more in tropical and subtropical areas because of the assumed availability of short-rotation species which might could adaptation easier for forest owners in these regions relative to others (Sohngen et al., 2001; Perez-Garcia et al., 2002; Tian et al., 2016), while others indicate that temperate areas will experience the largest increase in supply (Favero et al., 2018; Favero et al., 2021). The results are very sensitive to the climate change scenarios tested, the climate and vegetation models used and the climate drivers that are considered. For example, Tian et al. (2016) and Favero et al. (2018; 2021) used the same economic model (the global timber model) but different climate scenarios and vegetation models, obtaining different results.
The increasing supply induces lower global timber prices (medium confidence). Studies estimate that the prices will decline between 1% and 38% in 2100 with respect to a no climate change scenario depending on the model and the climate change scenario assumed (climate change is represented as a change in GHG concentration, global average temperature or radiative forcing) (Favero et al., 2018; Favero et al., 2021). Clearly, further studies are needed considering a wider set of vegetation and climate models and incorporating the impacts of extreme events (such as droughts and wildfires).
There are a number of national and regional scale studies exploring the impact of climate change on yields and markets of wood products, with mixed results. In Finland, it is projected that timber yield in the north will increase in Scots pine and birch stands by 33–145% and 42–123%, compared with the current climate, depending on the GCM and thinning regime using a 90-year rotation (10 individual GCM projections under the RCP4.5 and RCP8.5 forcing scenarios) (ALRahahleh et al., 2018). However, in Norway spruce stands, yield could decline by up to 35%, under GFDL-CM3 RCP8.5 and increase by up to 39%, under CNRM-CM5 RCP8.5, compared with the current climate.
In Germany, timber harvest was projected to increase slightly (<10%) in 2045 using the process-based forestry model (4C) driven by three management strategies (nature protection, biomass production and a baseline management) and an ensemble of regional climate scenarios (RCP2.6, RCP4.5, RCP8.5) (Gutsch et al., 2018). Similarly, average production of pulpwood in slash pine stands in the southeastern USA are projected to increase by 7.5 m 3 ha −1 for all climatic scenarios using the Physiological Processes Predicting Growth (3-PG) forest growth model by 2100 (RCP4.5 and RCP8.5; CanESM2) (Susaeta and Lal, 2018).
Box 5.6: Contributions of Indigenous and Local Knowledge: An Example
Indigenous and local people have long histories of adaptation to climate hazards in forests (see Eriksen and Hankins, 2014; Neale et al., 2019; Bourke et al., 2020; Long et al., 2020; Williamson, 2021 for notable examples in Australia and North America). In this section, we present a North American example of an indigenous adaptation practice developed by the Karuk Tribe in northern California. The Karuk Climate Adaptation Plan focuses on the use of cultural fire as climate adaptation, places a central importance on restoring human ecological caretaking responsibilities, and emphasises the need for collaboration, public education and policy advocacy to achieve these outcomes.
The Karuk Climate Adaptation Plan utilises a combination of Western science and Karuk traditional ecological knowledge. The plan centres on 22 focal species as cultural indicators as cues for human responsibilities and the particular techniques of fire application across seven habitat management zones (e.g., multiple forest types as well as riverine, riparian and montane systems). These adaptations range from specific prescriptions for the use of fire to lower river temperatures in acute scenarios (David et al., 2018), to protocols for treatment of grasslands and the use of high elevation meadows as fuel breaks. The plan also includes chapters on adaptations for tribal sovereignty, the mental and physical health effects of the changing climate and the protection of critical tribal infrastructure.
One aspect of Indigenous fire knowledge featured in the Karuk Climate Adaptation Plan is the culture-centric perspective on vegetation zones which are organised in relation to the elevation band in which smoke inversions occur (Figure Box 5.6.1). Within this system, burn timing follows a gradient that tracks the reproductive life cycles of season and elevational migrant species, the calving of elk and the nesting of birds. Within this system, elevational migrants are indicators of when to stop burning at one location and move upslope, following receding snows.
The plan also calls for the restoration of Indigenous fire science in emergency scenarios such as when rivers become too hot for salmon. With such fires localised, smoke inversions cool water temperatures through a variety of mechanisms, including shading river systems and reducing evapo-transpiration, thereby increasing stream flow (David et al., 2018).
Figure Box 5.6.1 | Seasonality and elevation dynamics of cultural indicators in Karuk Cultural Management Zones based in Karuk traditional ecological knowledge.
5.6.3 Adaptation
AR5 notes that natural ecosystems have built-in adaptation ability (Settele et al., 2014). However, this capacity will not be enough to prevent loss of forest ecosystem services because of projected climate change in this century under RCP6.0 and 8.5. Management actions could reduce the risks of impacts to forest ecosystems but only up to a certain point.
A systematic review of literature revealed that successful adaptation in forest management can be achieved if there are partnerships between key stakeholders such as researchers, forest managers and local actors (Keenan, 2015). Such partnerships will lead to a shared understanding of climate-related challenges and more effective decisions. Forest managers in some countries of the world seem to have high awareness of climate change (van Gameren and Zaccai, 2015; Seidl et al., 2016; Sousa-Silva et al., 2016). However, they need more information on how they can adjust their practices in response to climate change. Institutional and policy context needs to be considered to facilitate adaptation by forest managers (Sousa-Silva et al., 2016; Andersson et al., 2017).
5.6.3.1 Adaptation measures in sustainable forest management
A wide range of measures exist to adapt sustainably managed forests of the boreal and temperate zone to climate change (Kolström et al., 2011; Gauthier et al., 2014; Keenan, 2015). Evidence emerging since the last assessment report further bolstered the notion that adapting the tree species composition to more warm-tolerant and less disturbance-prone species can significantly mitigate climate change impacts (high confidence) (Duveneck and Scheller, 2015; Seidl et al., 2018). Assisting the establishment of species in suitable habitats is one option to achieve climate-adapted tree species compositions (Benito-Garzón and Fernández-Manjarrés, 2015; Iverson et al., 2019). Furthermore, increasing the diversity of tree species within stands can have positive effects on tree growth and reduce disturbance impacts (high confidence) (Neuner et al., 2015; Jactel et al., 2018; Ammer, 2019). Some studies also suggest a positive effect of increased structural diversity, such as on forest resilience (moderate confidence) (Lafond et al., 2013; Koontz et al., 2020). Managing for continuous forest cover can also help to maintain the forest microclimate and buffer tree regeneration and the forest floor community against climate change (high confidence) (De Frenne et al., 2013; Zellweger et al., 2020). Reducing stocking levels, such as through thinning, has been found to effectively mitigate drought stress (Gebhardt et al., 2014; Elkin et al., 2015; Bottero et al., 2017), yet effects vary with species and ecological context (robust evidence, medium agreement ) (Sohn et al., 2016; Castagneri et al., 2021). Also shortened rotation periods have been suggested in response to climate-induced increases in growth and disturbance (Jönsson et al., 2015; Schelhaas et al., 2015). However, recent evidence suggests that these measures diminish in efficiency under climate change and can have corollary effects on other important forest functions such as carbon storage and habitat quality (medium confidence) (Zimová et al., 2020). Also, measures targeting landscape structure and composition have proven effective for increasing the climate resilience of forest systems (medium confidence) (Aquilue et al., 2020; Honkaniemi et al., 2020). While an increasing number of adaptation measures exist for sustainably managed forests, many studies highlight that the lead times for adaptation in forestry are long and that some vulnerabilities might remain also after adaptation measures have been implemented. Furthermore, the costs and benefits of adaptation measures relative to other goals of sustainable forest management, such as the conservation of biological diversity, have to be considered (Felton et al., 2016; Zimová et al., 2020; see Cross-Chapter Paper 7.5 Adaptation Response Options).
5.6.3.2 Linking adaptation and mitigation through Reducing Deforestation and Forest Degradation plus
Reducing Deforestation and Forest Degradation plus (REDD+) is a climate mitigation strategy which could also provide important climate change adaptation co-benefits; for example, sustainable forest management could provide long term livelihoods to local communities and enhance resilience to climate risks (Turnhout et al., 2017). However, major challenges related to REDD+ implementation and forest use remain such that it has not been implemented successfully at scale (Table 5.8).
Table 5.8 | Challenges and solutions for REDD+
Challenges with REDD+ implementation | Solutions for successful forest management |
Legal: lack of carbon rights in national legislations (Sunderlin et al., 2018; RRI, 2018b); unclear forestland tenure systems (Resosudarmo et al., 2014). | There is high confidence that implementing social safeguards such as a Free Prior and Informed Consent (FPIC) is vital to adequately involving Indigenous Peoples and local communities in REDD+ (White, 2014; Raftopoulos and Short, 2019). Indigenous Peoples, consisting of at least 370 million people, manage or have tenure rights over a quarter of the world’s land surface (around 38 million km 2) encompassing about 40% of the world’s protected areas (Garnett et al., 2018; RRI, 2018a). |
Food security and livelihoods: negative impacts of REDD+ on food security, agroforestry and swidden agriculture (Fox et al., 2014; Holmes et al., 2017). | There is high agreement that REDD+ and other green adaptation and mitigation efforts need to cooperate with Indigenous Peoples and other local communities who depend on forest resources for their livelihoods and food security (Wallbott, 2014; Mccall, 2016; Brugnach et al., 2017; Vanclay, 2017; Garnett et al., 2018; Paneque-Galvez et al., 2018; Sunderlin et al., 2018; Schroeder and Gonzalez, 2019). |
Political and socio-cultural: land acquisition or ‘green grabbing’ (Asiyanbi, 2016; Corbera et al., 2017); (mis)communicating the concept of carbon (Kent and Hannay, 2020); and lack of influence of Indigenous and local communities’ representation in global and national REDD+ negotiations (Wallbott, 2014; Dehm, 2016). In the absence of social and environmental safeguards, REDD+ could drive large-scale land acquisitions by states and corporations, resulting in global land grabs (or green grabbing), negatively affecting the food security, livelihoods and tenure rights of Indigenous and local communities (limited evidence, high agreement ) (Carter et al., 2017; Lund et al., 2017; Borras et al., 2020). | There is low confidence as to whether community forestry is compatible with REDD+ (Hajjar et al., 2021). This is mainly due to lack of carbon payments and the variety of approaches to REDD+. There is high confidence that restoring land access and rights via transfer of formal land titles to Indigenous and local communities improves biodiversity conservation and carbon sequestration. |
5.7 Other Natural Products
Natural products such as medicinal plants, wild food (plants, animals, mushrooms) and resins (e.g., gum arabic and frankincense) have high commercial value and contribute an important source of livelihood in some regions. One in six persons globally live in or near forests, and many depend on forest resources for some of their livelihood and needs, particularly in low- and middle-income countries (Vira et al., 2016; Newton et al., 2020). The FAO has estimated that in 2011 non-wood forest products, including medicinal plants, contributed over 88 billion USD to the global economy (FAO, 2014). Greater diversity in local knowledge and Indigenous knowledge of natural resources supports resilience in the face of hazards, especially in environments with high levels of uncertainty (Berkes et al., 2003; Blanco and Carriere, 2016).
5.7.1 Medicinal Plants
The World Health Organization lists traditional medicine as an essential component of culturally appropriate healthcare (WHO, 2013). Medicinal plants make up the primary source of medicine for 70–95% of people in low- and middle-income countries and are used widely in wealthier countries (Applequist et al., 2020). Continued use of medicinal plants ensures millions of rural people have access to effective treatments for day-to-day illness and infection and thus improves their health and resilience to climate change.
Indigenous Peoples largely depend on medicinal plants for their healthcare need in different parts of the world (de Boer and Cotingting, 2014; Silva et al., 2020). Medicinal and aromatic plants can support the economy and generate livelihood options for rural people through preparing and selling traditional medicine; collecting from wild; and trade for income generation (Fajinmi et al., 2017; Zahra et al., 2020). Income from medicinal plant collection increases livelihood diversification, which is widely accepted to improve resilience.
5.7.2Resin and Gum
Resin and gum are economically important natural products, contributing 14–23% total household income in parts of Ethiopia and Sudan (Abtew et al., 2014; Fikir et al., 2016), Cambodia (Sakkhamduang et al.) and India (Tewari et al., 2017). They are an important source of raw material for many industries. For instance, in Africa, the genera Boswellia and Commiphora, which provide frankincense and myrrh resins, provide significant income generation and export value (Tilahun et al., 2015). Populations of many species that provide gums and resins are declining under pressure from unsustainable harvesting and deforestation, and climate change may threaten them further.
In Sri Lanka, Boswellia serrataRoxb. is critically endangered or possibly extinct (Weerakoon and Wijesundara 2012). In India, B. serrata populations are ‘vulnerable’ (Chaubey et al., 2015; Brendler et al., 2018), and declining in the Western Ghats (Soumya et al., 2019). Invasion of Lantana camara and Prosopis juliflora has resulted in poor regeneration of Commiphorawightii in central India (Jain and Nadgauda, 2013). Other resin-producing species under threat include: Daemonoropsdraco (dragon’s blood resin) in Indonesia (Yetty et al., 2013; Widianingsih et al., 2019), Pinus merkusii (tusam) in Sumatra (Indonesia) (Hartiningtias et al., 2020), Pinus pinaster in Spain, Pinus massoniana in China (Génova et al., 2014; Chen et al., 2015b) and Pistacia atlantica in Iran (Yousefi et al., 2020).
5.7.3 Wild Foods
Wild foods can include both native and introduced species that are not cultivated or reared but may be under various degrees of management by humans and may include escapees of species that are cultivated in some contexts (Powell et al., 2015). Information on the use and importance of wild foods for nutrition is growing but remains limited (FAO, 2019e). The AR4 covered wild food briefly in the polar regions and noted the inter-related nature of climate change and Indigenous knowledge loss in reducing access to wild food (Anisimov et al., 2001). AR5 did not address wild foods and other natural products. There is large variation in the importance of wild foods (Powell et al., 2015; Rowland et al., 2017; Dop et al., 2020). A recent survey of 91 countries found that 15 reported regular use of wild foods by most of the population, and 26 reported regular use of wild foods by a subsection of the population (FAO, 2019e). While they contribute little to food energy intake, their contribution to nutrition can be significant because most wild and forest foods (vegetables, fruits, mushrooms, insects and meat) are rich in proteins and micronutrients (Powell et al., 2015). The impacts of climate change on wild foods will vary in time and space and among species.
5.7.4 Observed and Projected Impacts
5.7.4.1 Medicinal plants
Research is limited on the effects of climate change on the distribution, productivity or availability of medicinal plants (Applequist et al., 2020), but some are facing threats due to climate change (Phanxay et al., 2015; Chirwa et al., 2017; Chitale et al., 2018). Climate change is projected to impact some medicinal plant species through changes in temperature, precipitation, pests and pathogens; unsustainable harvest of high-value species will significantly exacerbate these impacts (medium evidence, high agreement ) (Applequist et al., 2020). Table 5.9 highlights that climate change impacts on medicinal plant species will vary greatly by species. Medicinal plants that grow in arid environments are also highly susceptible to climate-induced change (Applequist et al., 2020). Arctic medicinal species may also be particularly at risk due to climate change (Cavaliere, 2009).
Table 5.9 | Observed and predicted impacts of climate change on selected medicinal plant species.
Region | Species | Observed and projected impacts of climate change | Assessment of evidence and level of agreement |
Egypt, Sub-Saharan Africa, Spain, Central Himalaya, China, Nepal | General assessment of medicinal plants | Habitat suitability and/or range distribution will shift or may be lost (Munt et al., 2016; Yan et al., 2017; Brunette et al., 2018; Chitale et al., 2018; Zhao et al., 2018; Applequist et al., 2020), including in high-elevation meadows which are home to some of the most threatened plant populations and contain a high number of and higher proportion of species used as medicine compared with lower-elevation habitats (Salick et al., 2009; Brandt et al., 2013). | Medium confidence |
Hindukush Himalaya | Gynostemmapentaphyllum | The elevated CO2 and temperature can increase biomass, but the health-promoting properties such as total antioxidants, phenols and flavonoids are expected to decrease (Chang et al., 2016). | Medium confidence |
Arctic | Golden root (Rhodiola rosea) | Population decline has been associated with drying of stream beds and alpine meadows, which are predicted to become more severe under climate change | Medium confidence |
North America | American ginseng (Panax quinquefolius) | Modelling of the combined impact of climate change (warming) and harvesting pressure indicates a nonlinear increase in extinction risk (Souther and McGraw, 2014). | Medium confidence |
Asia | Gentiana rigescens | A model evaluating future climate impacts shows a westward range shift and major loss of highly suitable habitats. Modelling also shows a potential decline in quality (chemical concentration of iridoid glycoside, which is highest in highly suitable habitats) due to climate change (Shen et al., 2021). | Medium confidence |
Africa | Alstoniaboonei | Modelling indicates that the range for this species remains relatively stable, with a possible modest expansion at the northern and southern margins of the range (Asase and Peterson, 2019). | Medium confidence |
Asia | Homonoia riparia | Modelling of future climate scenarios in Yunnan Province, China projects that habitat suitability improves (Yi et al., 2016). Modelling of future climate scenarios across the whole species range in China shows that both the suitable area and suitability of the habitat increase (Yi et al., 2018). | Medium confidence |
Asia | Notopterygiumincisum | Modelling for future climate change shows areas of suitable habitat will significantly decrease; however, the area of marginally suitable habitat will remain relatively stable (Zhao et al., 2020). | Medium confidence |
Himalayas | Himalayan yew Taxus wallichiana | Modelling shows projected shrink in climatic niche of the species by 28% (RCP4.5) and 31% (RCP8.5), highlighting the vulnerability to climate change impacts (Rathore et al., 2019). | Medium confidence |
Iran | Daphne mucronata | Modelling of future climate change projects disappearance of the species below 2000 m, significant change in distribution between 2000 and 3000 m and no change above 3000 m (Abolmaali et al., 2018). | Medium confidence |
Central America | Pericón or Mexican Mint Marigold Tagetes lucida | Models predict range to contract somewhat and shift northward (Kurpis et al., 2019). | Medium confidence |
Africa | Rooibos tea Aspalathus linearis | Modelling of future climate scenarios shows substantial range contraction of both wild and cultivated tea, with range shifts southeastwards and upslope (Lotter and Maitre, 2014). | Medium confidence |
Himalayas | Lilium polyphyllum | Habitats of this species will shrink by 38–81% under future climate scenarios and shift towards the southeast region in Western Himalaya, India (Dhyani et al., 2021). | Medium confidence |
Iran | Fritillaria imperialis | Modelling shows 18% and 16.5% of the habitats may be lost due to climate change by 2070 under RCP4.5 and RCP8.5, respectively. Further, it is observed that, under the current climatic conditions, the suitable habitat may become unsuitable in the future, resulting in local extinction (Naghipour Borj et al., 2019). | Medium confidence |
Himalayas/ China | Snow lotus (Saussurea spp. ) | Climate change is a significant threat to this species (Law and Salick, 2005). Laboratory and field trials show considerable plasticity and a wide thermal range for germination, which may help compensate for range reductions under climate change (Peng et al., 2019). | Medium confidence |
North Africa | Atlas cedar Cedrus atlantica | Modelling shows a significant and rapid contraction of distribution range, upward elevational range shift, increased fragmentation, and possible disappearance in many North African localities (Bouahmed et al., 2019). | Medium confidence |
Asia/South Korea | Paeonia obovata | Modelling of climate change scenarios shows significant loss of suitable habitat and possible disappearance of P. obovatain in South Korea after 2080 (Jeon et al., 2020). | Medium confidence |
Iran | Salvia hydrangea | A projected loss of habitat in the southeast of the range will not be compensated by the northward or upward elevational range migration (Ardestani and Ghahfarrokhi, 2021). | Medium confidence |
Patagonian, Argentina | Valeriana carnosa | Modelling for future climate scenarios projects a 22% loss of the suitable habitat (Nagahama and Bonino, 2020). | Medium confidence |
Western Ghats, India | Kokum Garcinia indica | Predictions of climate change impact on habitat suitability indicate drastic reduction in the suitability by over 10% under RCP8.5 for the years 2050 and 2070 (Pramanik et al., 2018). | Medium confidence |
Himalaya | Ophiocordyceps sinensis | A decline of the species is largely due to over harvesting, but ecological modelling indicates that climate warming is also contributing to this decline (Hopping et al., 2018). | High confidence |
Pacific islands | Noni (Morindacitrifoli), naupaka (Scaevola spp.), kukui (Aleurites moluccana) and milo (Thespesia populnea) | May be less susceptible to climate change as they are fast growing, have high reproduction rates, grow at sea level (and are often salt-tolerant) and have significant room for range shifts (Cavaliere, 2009). | Low confidence |
Changes in range distribution will interact with detailed local knowledge and Indigenous knowledge needed to harvest and use medicinal plants. Northward range shifts, for example, may mean certain plants still exist, but not where they have traditionally been important as medicine, and possibly moving suitable ranges outside of areas where plants species have sufficient protection (Kaky and Gilbert, 2017). Climate-induced phenological changes are already observed as a threat to some species (Gaira et al., 2014; Maikhuri et al., 2018). Other major climate-induced impacts on medicinal plants will be via the phytochemical content and pharmacological properties of medical plants (Gairola et al., 2010; Das et al., 2016a). Experimental trials have shown that drought stresses increase phytochemical content, either by decreasing biomass or increasing metabolites production (high confidence) (Selmar and Kleinwachter, 2013; Al-Gabbiesh et al., 2015).
5.7.4.2 Wild food
5.7.4.2.1 Wild food in the Arctic, North America and Europe
Changes to the availability, abundance, access and storage of wild foods associated with changing climate are exacerbating high rates of food insecurity (high confidence) (Ford, 2009; Beaumier and Ford, 2010; Herman-Mercer et al., 2019). Wild foods are central to the food systems of communities throughout the Arctic and sub-Arctic (Kuhnlein et al., 1996; Ballew et al., 2006; Kuhnlein and Receveur, 2007; Johnson et al., 2009) and play an essential role in people’s physical and emotional health (Section CCP6.2.5; 2.8) (high confidence) (Loring and Gerlach, 2009; Cunsolo Willox et al., 2012). Wild foods consumed in the Arctic and northern regions include animals and a wide variety of plant foods (Wein et al., 1996; Ballew et al., 2006; Kuhnlein and Receveur, 2007). Wild foods contribute most of important nutrients in the diets of northern and Arctic people (Johnson et al., 2009; Wesche and Chan, 2010; Kenny et al., 2018). However, the use of traditional wild foods is declining across the region, lowering diet quality (Rosol et al., 2016). Indigenous communities in the Arctic perceive climate change related impacts on traditional wild foods, and availability and access to wild foods are forecast to continue to decline (Brinkman et al., 2016). Some communities hold positive views of the new opportunities a warmer climate will bring, seeing them as a favourable trade-off relative to the loss of some forms of subsistence hunting (Nuttall, 2009). Climate change is causing ecological changes that impact Arctic wild food availability and abundance in many different ways, including changes to breeding success, migration patterns and food webs (Table 5.10, Markon et al., 2018).
Table 5.10 | Observed and predicted impacts of climate change on selected wild food species.
Region | Species | Observed and projected impacts of climate change | Assessment of evidence and level of agreement |
Arctic region | Ringed seals (Pusahispida) | Drastic declines in population size and major changes in population structure (Hammill, 2009; Reimer et al., 2019); habitat (dependent on snow cover or ice breathing holes for lairs) will decline by approximately 70%, and significantly reduce survival rates of pups (Freitas et al., 2008). | High confidence |
Arctic region | Bearded seal (Erignathus barbatus) | Climate change affects the availability and stability of at least 11 ice-associated species, including bearded seal. Potential impacts due to climate change will reduce available habitat for birthing (Moore and Huntington, 2008; Fink, 2017). | Medium evidence, high agreement |
Arctic region | Walrus (Odobenus rosmarus) | Declines in the climate-vulnerable Pacific walrus populations, induced by overharvesting (Taylor et al., 2018); however, the species is considered highly vulnerable to loss of sea ice (Lydersen, 2018). Possible diet changes (related to climate-induced changes in food web) raise concerns about the health of the population (Clark et al., 2019). | High confidence |
Arctic region | Narwhal (Monodon monoceros) | The impacts of climate change on other sea ice-associated marine mammals are somewhat less clear (Moor et al., 2017). Climate change may threaten narwhal given their vulnerability to ice entrapment (Laidre and Heide-Jorgensen, 2005) and the narrow range of prey in their diet (Heide-Jørgensen, 2018). In Greenland, hunters report that narwhal now frequent fjords and other areas where manoeuvring a boat is difficult (Nuttall, 2017). | Low evidence, medium agreement |
Arctic region | Beluga (Delphinapterus leucas) | Belugas are thought to be less sensitive to climate change than some other sea mammals but can perish in large groups from ice entrapment. Climate impacts likely increased human activity (noise) (O’Corry-Crowe, 2009). Changes in migrating timing have been documented (Hsiang et al., 2017). | Low evidence, low agreement |
Arctic region | Bowhead (Balaena mysticetus) | The movements of some whale species are linked to SSTs (Moore and Huntington, 2008; Chambault et al., 2018). Some whale hunting communities are now reporting that whales pass by at a time of year when launching boats is impaired by rough weather and poor sea ice conditions (Noongwook et al., 2007; Huntington et al., 2017). | Medium confidence |
Arctic region | Other sea ice associated marine mammals (harp seal, hooded seal) | The impacts of climate change on other sea ice associated marine mammals are somewhat less clear (Moor et al., 2017). | Low confidence |
Arctic and northern regions | Reindeer and caribou (Rangifer tarandus) | Large herbivores are highly dependent on their food sources such as mosses, lichens and grasses which are sensitive to climate change (Istomin and Habeck, 2016). Combined impacts of climate change and other inter-related factors suggest significant declines in caribou and reindeer populations, although to varying extents from one population to another (Kenny et al., 2018; Mallory and Boyce, 2018). Warming has led to increased plant productivity and associated increases in body mass of some reindeer populations (Albon et al., 2017; Mallory and Boyce, 2018). Increasing primary production, warming will also change the plant composition, leading to increases in woody/shrubby vegetation which will have negative nutritional consequences for caribou and reindeer (Elmendorf et al., 2012; Mallory and Boyce, 2018). The loss of lichens, a key winter food source, due to increased wildfire or replacement by grasses and herbs that die back in the winter, may also be detrimental to caribou and reindeer, although there is currently no consensus on this among experts (Mallory and Boyce, 2018). Rain on snow and icing events during winter, which are predicted to become more frequent, have been documented to lead to large increases in arctic herbivore mortality because they create an ice barrier making access to food more difficult (Putkonen and Roe, 2003; Tyler, 2010; Stien et al., 2012; Hansen et al., 2013; Forbes et al., 2016). Rain-on-snow events may also impact reproductive success, although recent research suggests this relationship in not straightforward (Douhard et al., 2016). Increased summer insect harassment is also predicted to increase and further stress large herbivores both by the additional parasitic load and by decreasing the amount of time spent grazing as animals seek to outrun pests (Mallory and Boyce, 2018). Finally, many caribou and reindeer populations rely on sea and freshwater ice to facilitate their movement and migration; loss of ice may make some populations no longer viable (Mallory and Boyce, 2018). | Medium confidence |
Arctic and northern regions | Moose (Alces alces) | The distributional changes of Rangifer populations might be affected by the range expansions and the northward expansion of moose (Mallory and Boyce, 2018). This is due to increases in productivity on the tundra and more frequent wildfire activity resulting in improved habitat quality for moose northward. | Medium confidence |
North America | Geese (Branta canadensis, Answer spp. , Branta spp. ) | Phenological mismatch developing between the berries and migration timing may mean that Canadian geese no longer stop near some communities (Downing and Cuerrier, 2011). | Medium confidence |
Arctic and northern regions | Berries (Vaccinium spp. , Rubus spp. and others) | Berries are among the most important and widely consumed wild foods of plant origins in Arctic and northern regions (Vaara et al., 2013; Hupp et al., 2015; Boulanger-Lapointe et al., 2019). Berry production will be impacted by climate change, including snow cover, rainfall, soil moisture, air temperature and availability of insect pollinators (Herman-Mercer et al., 2020) and possible risk from sea-level-rise-associated soil salinisation (Cozzetto et al., 2013). Increased growth of woody shrub vegetation, driven by increased temperatures, can also make moving across the land difficult, impairing access to berry patches (Boulanger-Lapointe et al., 2019). Conversely, a recent modelling experiment suggested that the >2°C warming experienced by Arctic communities over the past three decades has had minimal impact on overall trail access (Ford et al., 2019). In Alaska, communities perceive berry abundance as declining and/or becoming more variable (Kellogg et al., 2010; Hupp et al., 2015). In a Gwich’in community in Canada, Parlee and Berkes (2005) recorded that local women perceived climate change, especially extreme weather events, as the greatest risk to traditional berry patches (cranberry, blueberry and cloudberry). The expansion of trees and shrubs may cause shading and negatively impact the productivity of berry plants (Downing and Cuerrier, 2011; Lévesque et al., 2012). Berries are predicted to be increasingly susceptible to negative impacts of invasive species (which compete for pollinators) (Spellman and Swenson, 2012) and infections (Turner and Clifton, 2009) as climate change progresses. Suitable area of huckleberry (Vaccinium membranaceum) would shrink by 5–40% by the end of the 21st century (Prevéy et al., 2020). Phenological shifts are also important. Many communities report changes in phenology including failed ripening or ‘all of the berries are ripening at the same time’ (Turner and Clifton, 2009; Herman-Mercer et al., 2020). Competition with growing populations of geese is viewed by many communities to be an important threat to berry harvesting. (Boulanger-Lapointe et al., 2019). In Labrador, Canada, changes in permafrost, vegetation, water and weather have had an impact on cloudberry (bakeapple) productivity, phenology and patch fragmentation. Moreover, changes in summer settlement patterns (which are now farther from berry patches) are making it more difficult for people to respond to variations in growth and timing (Anderson et al., 2018). In Montana, USA, Crow Nation elders have noted that many of their important berry resources have been impacted by climate change, either because they bud earlier and are then vulnerable to cold snaps, or the timing of fruit production has changed (with many now ripening at the same time) (Doyle et al., 2013). Similarly, the Wabanaki Nations in Maine and Eastern Canada worry that climate change will impact berry resources already under pressure from dwindling territory and pollution (Lynn et al., 2013). | High confidence |
North America (Washington State, USA) | Salmon (Salmonidae) | Indigenous communities in Washington State, USA report devastation of their salmon fishery due to loss of glacial run off and associated warming river and stream temperatures; potential damage to shellfish resources due to sea level rise and ocean acidification (Lynn et al., 2013). The Karuk people in California have also experienced losses in salmon (Lynn et al., 2013; Vinyeta et al., 2016). | Medium confidence |
North America (California) | Acorns from oak trees (Querus) | In the arid southwest of the USA, wild foods are less widely consumed today, but their revitalisation is important to identity and well-being of many Indigenous People. The Karuk people of the Klamath River in California have experienced an almost complete loss of two key traditional wild foods: salmon and acorns, foods which once made up 50% of a traditional Karuk diet (Lynn et al., 2013; Vinyeta et al., 2016), as well as huckleberry (Vinyeta et al., 2016). Using regional climate models, Kueppers (2005) showed a major reduction in the range of two species of oak in California that are used in traditional diets. Increasing frequency of severe fires in the western USA threaten a number of traditional wild food resources, especially acorns (Vinyeta et al., 2016). | Medium confidence |
North America | Wild rice (Zizania spp.) | Significant reductions in wild rice area in Great Lakes have been associated with mining, dams and other activities, but climate change may lead to further reductions (Cozzetto et al., 2013; Lynn et al., 2013). | High confidence |
North America | Camas tuber (Camassia quamash) | Historic changes in fire regimes, linked to changes in climate, are believed to have altered availability of the important camas tuber (Camassia quamash) (Lepofsky et al., 2005). | Medium confidence |
North America | Wapato tuber (Sagittaria latifolia) | The aquatic Sagittaria latifolia (the roots of which are consumed by Indigenous groups across North America) is vulnerable to both water salinity and temperature (Delesalle and Blum, 1994). | Medium confidence |
North America | Springbeauty (Claytonia lanceolate) | Claytonia lanceolata is particularly vulnerable to changes in snow melt and other climatic changes owing to advancement in the flowering (Renner and Zohner, 2018). | Medium confidence |
North America | Seaweed (Porphyra abbottiae, among others) | In British Columbia, Canada, Indigenous People Gitga’at elders noted that the ripening of an important edible seaweed (Porphyra abbottiae) had rarely synchronised with weather patterns that enabled them to process it in the traditional way (drying on rocks and then ripening and re-drying) (Turner and Clifton, 2009). | Low confidence |
Africa | Baobab (Adansonia digitata) | Baobab is thought to be vulnerable to climate change because it is long-lived, it can take up to 23 years to start fruiting and leaf harvesting is often so intensive that it depresses fruit production. Modelling study using different records model shows that the percentage of present distribution predicted to be suitable in the future ranged from 5% to 91% (Sanchez et al., 2011). | Low confidence |
Africa | Shea (Vitellaria paradoxa) | Shea (Vitellaria paradoxa) was expanded through human intervention and is linked to human migration; fruit traits such as fruit size and shape, pulp sweetness and kernel fat content are determined both by temperature and rainfall, as well as human selection for preferred traits (Maranz and Wiesman, 2003). There is limited and conflicting evidence of the impacts of climatic conditions and future projected climate variations on V. paradoxa (Tom-Dery et al., 2018). Mixed evidence of the impact of climate and rainfall on fruit production and timing is reported (Tom-Dery et al., 2018). Fruit production was negatively correlated with mean annual temperature and positively correlated with annual rainfall (Bondé et al., 2019). | Limited evidence, medium agreement |
North Africa (Morocco) | Argan (Argania spinosa) | Climate change projections suggest a 32% decrease in habitat suitable for Argania spinosa under some scenarios (Alba-Sánchez et al., 2015; Moukrim et al., 2019). | Medium confidence |
Asia (Nepal) | Fruit species and vegetables (e.g., Asparagus racemosus, Urticadioica) | In Nepal, Thapa (2015) reports phenological changes in semi-domesticated fruit species, as well as decreased availability of a number of wild plants that can be consumed as vegetables. | Limited evidence, medium agreement |
Worldwide, most important in Europe and Asia | Mushrooms | Wild mushrooms production (including truffles) is closely linked to climate factors including temperature and precipitation as well as tree growth and carbohydrate production (Tahvanainen et al., 2016). Some species are sensitive to high temperatures (Büntgen et al., 2012; Le Tacon et al., 2014; Ágreda et al., 2015; Bradai et al., 2015; Taye et al., 2016; Alday et al., 2017; Karavani et al., 2018; Büntgen et al., 2019; Thomas and Buntgen, 2019). Models for some varieties suggest ‘declines of 78–100% in European truffle production are likely for 2071–2100’ (Thomas and Buntgen, 2019). For some species in northern Europe, the season is expanding (starting earlier and/or ending later), likely linked to warming (Büntgen et al., 2012; Le Tacon et al., 2014; Ágreda et al., 2015; Bradai et al., 2015; Taye et al., 2016; Alday et al., 2017; Karavani et al., 2018; Büntgen et al., 2019; Thomas and Buntgen, 2019). Matsutake mushroom (Tricholoma matsutake), highly prized in China, is sensitive to timing and amount of precipitation and temperature (Yang et al., 2012), and suitable habitat for this species is predicted to significantly decrease and highly suitable habitat to nearly disappear under various climate change scenarios (Guo et al., 2017). | High confidence |
North America (California) | Acorns, nuts and berries and other fire-dependent wild foods | Low-intensity traditional burning practices increased pyro-diversity (Vinyeta et al., 2016). Climate change will exacerbate the risks posed by exotic pathogens that attack oak species and further reduce access to acorns as well as other foods founds in oak ecosystems (Voggesser et al., 2013). | High confidence |
South America (Amazon region) | Aguaje, (Mauritia felxuosa), Brazilian nut (Bertholletiaexcelsa), fishing and hunting in general | Local communities perceived a lower yield of aguaje due to drought (Hofmeijer et al., 2013). In another study from the Colombian Amazon, wild food use was reported to be vulnerable to extreme climate events which impact species migration patterns or restrict access to fishing and hunting rounds (Torres-Vitolas et al., 2019). In some humid regions, the range of some wild food species may be extended by climate change, such as the Brazilian nut (Bertholletiaexcelsa) (Thomas et al., 2014). | Medium confidence |
Small islands (Papua New Guinea) | Sweet potato | Increases in the ENSO were associated with drought which increased sweet potato losses (Jacka, 2016) in highlands humid forest. | Limited evidence, medium agreement |
Australasia (Australia) | General wild foods | Aboriginal communities in North Queensland, a humid tropical region of northern Australia, reported some climate impacts on wild foods, although primarily for marine resources and those found in dry forest ecosystems (McIntyre-Tamwoy et al., 2013). | Limited evidence, medium agreement |
Asia (Indonesia) | Sago (Metroxylon sagu) | People in a sago-dependent community in Papua Indonesia viewed climate variation as less important than other factors (logging, mining, infrastructure), but still expressed concerns about salinity of water supplies, floods and reduced hunting success (Boissière et al., 2013). | Limited evidence, medium agreement |
Climate-change-induced impacts of access to wild foods are also of concern in Arctic regions (high confidence). Coastal and inland communities of Alaska found that 60% of climate impacts on food security listed by hunters were related to access (Brinkman et al., 2016). Reduced duration, thickness and quality of sea ice are some of the most cited impacts of climate change on wild food consumption (Ford, 2009; Laidler et al., 2009; Downing and Cuerrier, 2011; Huntington et al., 2017; Nuttall, 2017; Fawcett et al., 2018; Ford et al., 2018; Markon et al., 2018). Lack of snowfall reduces and delays the ability to travel on land using snowmobiles (Downing and Cuerrier, 2011), impacting safety of travel, time needed and costs of accessing wild foods (Cold et al., 2020).
Rising temperatures and humidity are also impacting wild food storage and increasing the risk of food-borne diseases (Cozzetto et al., 2013; Nuttall, 2017; Markon et al., 2018). Changes in AT and humidity can mean that whale and fish meat no longer dry properly, or meat may spoil before hunters can get it home (Downing and Cuerrier, 2011; Nuttall, 2017). Traditional permafrost ice cellars are no longer reliable (Downing and Cuerrier, 2011; Nyland et al., 2017; Herman-Mercer et al., 2019). Climate-related environmental change compounded with social, economic, cultural and political change have had complex but overall negative impacts on wild foods (Section CCP6.4, Lujan et al., 2018) .
Communities across other (non-Arctic) parts of North America and Europe also report declining availability of wild foods, with climate change among the perceived drivers for decline (medium confidence) (Table 5.10, Serrasolses et al., 2016; Smith et al., 2019a). Even when climate change may not always be the primary driver of loss of these wild food resources, climate may interact with other stressors to exacerbate loss of wild foods (Lynn et al., 2013; Reo and Parker, 2013).
5.7.4.2.2 Wild food in the arid and semi-arid environments
Wild foods are also impacted by climate change in arid and semi-arid landscapes around the world (medium evidence, high agreement ) (Table 5.10). A number of wild species are important traditional foods of Indigenous Peoples or local communities across arid regions of North America (Messer, 1972; Kuhnlein and Calloway, 1977; Santos-Fita et al., 2012; Vinyeta et al., 2016), South America (e.g., Argentina; Ladio and Lozada, 2004; Altrichter, 2006; Eyssartier et al., 2011), Australia (Scelza et al., 2014), the Mediterranean Basin (Hadjichambis et al., 2008; Powell et al., 2014), India and the Himalayas (Pingle, 1975; Gupta and Sen, 1980; Delang, 2006; Bhatt et al., 2017).
Wild foods such as baobab, shea and nere from plants and animals make an important contribution to diets and nutrition in arid and semi-arid regions of Africa (Boedecker et al., 2014; Leßmeister et al., 2015; Bélanger and Pilling, 2019) and are being impacted by climate change (Moseley et al., 2015; Sango and Godwell, 2015; Hitchcock, 2016) (see Chapter 9). There has been little published research on the impacts of climate change on wild food in arid regions of Australia, although Aboriginal elders in one report suggested that climate-related changes are impacting wild food (Memmott et al., 2013).
5.7.4.2.3 Wild food in tropical humid environments
Wild foods are important to many communities that live in and adjacent to humid tropical forests, but climate change impacts are mixed (Table 5.10, Dounias et al., 2007; Colfer, 2008; Powell et al., 2015; Rowland et al., 2017; Reyes-García et al., 2019). In some humid tropical forest regions, bushmeat is particularly important (Golden et al., 2011; Nasi et al., 2011; Fa et al., 2015; Powell et al., 2015; Rowland et al., 2017). In humid tropical regions, the impact of climate change on wild food availability, access and consumption is currently unclear and research is limited. There are, however, important interrelationships between climate change and wild food use in humid forests. For example, the loss of large mammals to bushmeat consumption and global trade will likely slow the regeneration of tropical forests in which a large number of tree species are dependent on large mammals for seed dispersal (Brodie and Gibbs, 2009). Conversely, others argue that bushmeat provides local communities with an important incentive to support local maintenance of forest cover and, thus, carbon sequestration (Bennett et al., 2007).
5.8 Ocean-Based and Inland Fisheries Systems
The livelihoods of 10–12% of the world’s population depend on fisheries and aquaculture (FAO, 2020c). Globally, fish provide more than 3.3 billion people with 20% of their average per capita intake of animal proteins, reaching 50% or more in countries such as Bangladesh, Cambodia, The Gambia, Ghana, Indonesia, Sierra Leone, Sri Lanka and several Small Island Developing States (FAO, 2020c). Between 1961 and 2017, the average annual apparent global food fish consumption increased (3.1% per year; from 9.0 kg per person in 1961 to 20.5 kg in 2018), exceeding the rate of increase in consumption of meat from all terrestrial animals combined (2.1% annually, currently around 40 kg per person) (FAO, 2020d). Fish are a rich source of protein and specific vitamins and minerals (Khalili Tilami and Sampels, 2018), and are an essential food source in regions in need of nutritious, affordable food (Thilsted et al., 2016; FAO et al., 2018; Hicks et al., 2019; Cross-Chapter Box MOVING PLATE this chapter).
Overall capture fishery production has remained relatively static since the 1990s, reaching 96.4 million tonnes in 2018, with over 87% of the production coming from marine environments and the rest from inland fisheries (FAO, 2020c). Finfish represent 85% of global marine seafood production, with small pelagic fishes (anchovies, sardines and herrings) as the major contributor. Almost 60% of the total global marine catches come from China, Peru, Indonesia, the Russian Federation, the USA, India, Viet Nam, Japan, Norway and Chile (FAO, 2020c). Inland fisheries are found on every continent other than Antarctica and provide 158 million people the equivalent of all dietary animal protein (McIntyre et al., 2016). Inland production accounted for 12 million tonnes in 2018, with nearly 70% of capture from low-income Asian and African countries (Harrod et al., 2018a).
The aquaculture and fisheries’ share of gross domestic product (GDP) varies mostly from 0.01% to 10% (Cai et al., 2019), but the relative importance in countries’ economies and welfare is greater in several low-income countries, especially in many African and Pacific Island states. Approximately 60 million people are directly employed in fisheries value chains, from harvesting to distribution (Vannuccini et al., 2018); around 95% of them are in small-scale fisheries of low- and middle-income countries, and almost half are women.
5.8.1 Observed Impacts
Ocean systems are already facing significant impacts of climate change. At the ocean surface, temperature has on average increased by 0.88 [0.68–1.01] °C from 1850–1900 to 2011–2020 (Fox-Kemper et al., 2021; Gulev et al., 2021). Marine heatwaves have increased in frequency over the 20th century, with an approximate doubling since the 1980s (high confidence), and their intensity and duration have also increased (medium confidence) (IPCC, 2021, Box 9.2). In the Northeast Pacific, for example, an intense and long-lasting marine heatwave during 2013–2015 bridged to the strong 2015–2016 El Niño (Tseng et al., 2017) resulted in over 5 years of warmer-than-normal temperatures affecting the migration, distribution and abundance of several marine species, including fisheries resources (Cornwall, 2019; Jiménez-Quiroz et al., 2019). The surface open ocean pH has declined globally over the last 40 years by 0.003–0.026 pH per decade (virtually certain), and a decline in the ocean interior pH has been observed in all ocean basins over the past two to three decades (high confidence) (Gulev et al., 2021). The ocean is losing dissolved oxygen (very likely ) in the range of 0.5–3.3% between 1970 and 2010 for the 0–1000 m depth stratum (Bindoff et al., 2019; Canadell et al., 2021), salt content is being redistributed (very likely ) (Liu et al., 2019a; Gulev et al., 2021) and vertical stratification is increasing (virtually certain) (HLPE, 2017a; Fox-Kemper et al., 2021; Ranasinghe et al., 2021). There is high confidence that all these new physical, chemical and biological conditions affect marine organisms’ physiology, distribution and ecology, with an overall shift in biomass and species composition affecting ecosystem structure and function (Chapter 3). Under climate change, freshwater ecosystems are highly exposed to eutrophication, species invasion and rising temperatures (Lynch et al., 2016; Hassan et al., 2020). Major threats to wetland fisheries include water stress, sedimentation, weed proliferation, sea level rise and loss of wetland connectivity (Naskar et al., 2018).
Changes in aquatic ecosystems directly affect humans by altering livelihood, cultural identity and sense of self, and seafood provision, quality and safety. The state of marine fishery resources has continued to decline, with the proportion of fish stocks at biologically unsustainable levels of exploitation increasing from 10% in 1974 to 34.2% in 2017 (FAO, 2020d). There is medium confidence that fisheries production declines in different world regions can be partly attributed to climate change, along with overfishing and other socioeconomic factors. It has been estimated that, from 1930 to 2010, the amount of fish that can be sustainably harvested from several marine fish populations has decreased by 4.1% globally due to ocean warming, with some regions (East Asian Marginal Seas, the North Sea, the Iberian Coast and the Celtic-Biscay Shelf), experiencing losses of 15–35% (Free et al., 2019). There is regional variation such as redistribution of fishing grounds, due to climate-induced fish species migrations (Cross-Chapter Box MOVING PLATE this chapter). In Tanzania, for example, most small-scale fishers (75%) have reported shifting fishing grounds from nearshore to offshore areas during the last decade, due to perceived combined effects of overfishing and environmental impacts (Silas et al., 2020). Observed impacts in some inland aquatic systems indicate substantial productivity reductions (medium confidence). For example, sustained warming in Lake Tanganyika during the last ∼150 years has affected the biological productivity by strengthening and shallowing stratification of the water column (Cohen et al., 2016). Still, over 60% of the published reports on directly observed impacts of climate change on freshwater biota are on salmonids in North America and Europe, highlighting significant literature gaps for other fish species and regions (Myers et al., 2017a).
There is low confidence in climate change affecting the nutritious value of seafood. Contrasting evidence suggests that ocean warming and acidification could be altering the nutritional quality of commercial mollusks, primarily by reducing healthy fatty acids content (Tate et al., 2017; Ab Lah et al., 2018; Lemasson et al., 2019), but Coleman (2019) found no significant changes in a widely distributed coastal fish species.
In terms of food safety, there is high confidence that climate change increases the trends in seafood consumption related illnesses due to biological agents such as algae-produced toxins, ciguatera and Vibrio (Cross-Chapter Box ILLNESS in Chapter 2, Sections 5.11 and 5.12). Increased surface water warming changes the occurrence, intensity, species composition and toxicity of marine and freshwater algae and bacteria, and expansion to areas where they had not been reported before (Botana, 2016; McCabe et al., 2016; Griffith et al., 2019). There is limited evidence suggesting that risks linked to the bioaccumulation of chemicals are also of concern, such as neurotoxic methylmercury (MeHg) and heavy metals, due to water quality and trophic changes induced by climate change (Shi et al., 2016; Schartup et al., 2019).
5.8.2 Assessing Vulnerabilities
In the absence of adaptive measures, climate-induced changes in the abundances and distributions of fish will impact the provision, nutrition and livelihood security of many people (high confidence) as well as regional and global trade patterns (medium confidence).
5.8.2.1 Food security: provision and nutrition
The importance of seafood in food security and nutrition is increasing, largely due to its contribution as high-quality food (high confidence) (Hicks et al., 2019), as seafood contains unique long-chain polyunsaturated fatty acids (LC-PUFAs) and highly bioavailable essential micronutrients—vitamins (A, B and D) and minerals (calcium, phosphorus, iodine, zinc, iron and selenium). These compounds, often not readily available elsewhere in diets, have beneficial effects for adult health and child cognitive development (HLPE, 2014). Changes in marine and freshwater fish production can have significant consequences for human nutrition (Colombo et al., 2020). These changes are of particular concern in regions with few nutrition alternatives, such as low-income countries in Africa, Asia, Australasia, and Central and South America (high confidence) (Ding et al., 2017; Kibria et al., 2017).
Freshwater ecosystems that support most inland fisheries are under continuing threat from changes in land use, water availability and pollution and other pressures that will be exacerbated by climate change (high confidence) (Section 4.3.5). Declines in dissolved oxygen in freshwater are 2.75–9.3 times greater than observed in the world’s oceans (Jane et al., 2021). These systems have a relatively low buffering capacity and are therefore more sensitive to climate-related shocks and variability (Harrod et al., 2018b). Freshwater faunae are projected to be highly vulnerable; in the tropics because organisms are closer to approaching their thermal physiological limits and in the northern hemisphere (30–50°N) because the rate of temperature change is faster (Comte and Olden, 2017). The worldwide spatial confluence of productive freshwater fisheries and low food security highlights the critical role of rivers and lakes in providing locally sourced, low-cost, nutritious food sources (McIntyre et al., 2016).
Deltas and other wetland fisheries are extremely vulnerable to climate change and home to a large and growing proportion of the world’s population. In India, Ghana and Bangladesh, where three of the most populated Deltaic systems are located, subsistence fisheries provide 12–60% of the animal protein in people’s diets (Lauria et al., 2018).
The concern over aquatic food products’ safety due to climate change is increasing (high confidence). A strong positive relationship exists between specific bacterial growth rates and temperature, including pathogenic species of the genera Vibrio, Listeria, Clostridium, Aeromonas, Salmonella, Escherichia and others, whose distributional area is expanding with changing climate conditions (Cross-Chapter Box ILLNESS in Chapter 2, Section 5.12.1).
5.8.2.2 Social vulnerabilities, including gender and marginalised groups and cultural services
There is high confidence that climate change is and will continue to be a threat to the livelihood of millions of fishers, with the most vulnerable being those with fewer opportunities and less income (Barange and Cochrane, 2018; Section 3.4.3). The social vulnerability can differ largely between locations, even between relatively close coastal or inland communities (Bennett et al., 2014; Maina et al., 2016; Ndhlovu et al., 2017; Martins et al., 2019) and among inhabitants within a location, depending on factors such as access to other economic activities, education, health, adults in the household, and political connections (high confidence) (Senapati and Gupta, 2017; Abu Samah et al., 2019; Lowe et al., 2019).
Indigenous coastal communities consume 1.5–2.8 million metric tonnes of fish per year (about 2% of global yearly commercial marine catch), and reach a per capita consumption estimated to be 15 times greater than that of non-Indigenous country populations (Cisneros-Montemayor et al., 2016). There is high confidence that some Indigenous fishing communities are particularly vulnerable to climate change through a reduced capacity to conduct traditional harvests because of limited access to, or availability of, fish resources (Weatherdon et al., 2016), with consequences that include dietary shifts with significant nutritional and health implications (Marushka et al., 2019), displacement and loss of cultural identity (Sullivan and Rosenberg, 2018) and loss of social, economic and cultural rights (Finkbeiner et al., 2018). Areas of high risk for Indigenous Peoples include the Arctic, coastal communities with a high dependency on marine and freshwater fisheries, and Small Island States and Territories (Finkbeiner et al., 2018; Hanich et al., 2018, Section CCP6.2.5.1).
Women play a crucial role along the entire fisheries value chain, providing labour force in industrialised and small-scale fisheries all around the world (FAO, 2020d). For small-scale fisheries alone, women represent about 11% of the labour force, and their activity is generally in subsistence fisheries, highlighting their role in household food security (Harper et al., 2020). In general, gendered division of labour tends to cause lower salaries for women and different perception and experience of risk to climate change impacts (high confidence) (Lokuge and Hilhorst, 2017).
5.8.2.3 Management, economic and geopolitical vulnerabilities
Local, national, regional and international fisheries are mostly underprepared for geographic shifts in marine animals driven by climate change over the coming decades (high confidence) (Pinsky et al., 2018; Oremus et al., 2020; Pinsky et al., 2020). With fisheries distribution changes, sometimes into areas dedicated to different historical uses or new ventures, the current management regimes will face constraining legal frameworks (Farady and Bigford, 2019; Pinsky et al., 2020), which will demand interventions in the form of policies, programmes and actions, at multiple scales (Cross-Chapter Box MOVING PLATE this chapter). Coordinated fisheries management can substantially expand capacity to respond to a changing climate (Pinsky et al., 2020), but a great deal of political will, capacity building and collective action will be necessary (high confidence) (Teslić et al., 2017; Burden and Fujita, 2019; Section 5.8.4).
Today, approximately half the world’s population (~4 billion out of 7.8 billion people) are assessed as being currently subject to severe water scarcity for at least 1 month per year (medium confidence) (Box 4.1), and freshwater inland fisheries are particularly vulnerable as they are given lower priority for water resources than other sectors (high confidence). In some cases, this situation results in the total loss of freshwater fisheries. Examples include diversion of water for agriculture, shifts from food provision to recreational fisheries, conserving biodiversity, and the requirement for high-quality water for drinking water supply (Section 5.13, Harrod et al., 2018a).
There is high confidence that climate change increases the risk of conflicts due to the redistribution of stocks and their abundance fluctuations, with subsequent impacts on resource sharing (Spijkers and Boonstra, 2017; Pinsky et al., 2018; Spijkers et al., 2018; Mendenhall et al., 2020; Pinsky et al., 2020). High vulnerability and lack of adaptive capacity to climate change impacts (including fisheries-dependent livelihoods, attachment to place, and pre-existing tensions) increase the risk of conflicts, including among fishery area users and authorities (Ndhlovu et al., 2017; Shaffril et al., 2017; Spijkers and Boonstra, 2017; Mendenhall et al., 2020). Similarly, shifts in the distribution of transboundary fish stocks under climate change alter the current sharing of resources between countries and create conflicts as well as new opportunities (Cross-Chapter Box MOVING PLATE this chapter, Spijkers and Boonstra, 2017; Pinsky et al., 2018).
5.8.3 Projected Impacts
There is medium confidence that climate change will reduce global fisheries’ productivity (Section 3.4.4.2.3), with more significant reductions in tropical and subtropical regions and gains in the poleward areas (Bindoff et al., 2019; Oremus et al., 2020). Through an ensemble of marine ecosystem models and Earth System Models, mean global animal biomass in the ocean has been estimated to decrease by 5% under the RCP2.6 emissions scenario and 17% under RCP8.5 by 2100, with an average decline of 5% for every 1°C of warming (Lotze et al., 2019), affecting food provision, revenue distribution, and potentially hindering the rebuilding of depleted fish stocks (Britten et al., 2017). The projected declining rates result in a 5.3–7% estimated global decrease in marine fish catch potential by 2050 (Cheung et al., 2019), particularly accentuated in tropical marine ecosystems and affecting many low-income countries (Barange and Cochrane, 2018; Bindoff et al., 2019; Cross-Chapter Box MOVING PLATE this chapter). Projections indicate that by 2060 the number of exclusive economic zones (EEZs) with new transboundary stocks will increase to 46 under strong mitigation RCP2.6, and up to 60 EEZs under the RCP8.5 GHG emissions scenario (Pinsky et al., 2018). Similarly, by combining six intercompared marine ecosystem models, Bryndum-Buchholz et al. (2019) projected that under the RCP8.5 scenario a total marine animal biomass decline of 15–30% would occur in the North and South Atlantic and Pacific and the Indian Ocean by 2100. In contrast, polar ocean basins would experience a 20–80% increase. In the eastern Bering Sea, simulations based on RCP8.5 predict declines of pollock (>70%) and cod (>35%) stocks by the end of the century (Holsman et al., 2020). Temperate tunas (albacore, Atlantic bluefin and southern bluefin) and the tropical bigeye tuna are expected to decline in the tropics and shift poleward by the end of the century under RCP8.5, while skipjack and yellowfin tunas are projected to increase abundance in tropical areas of the eastern Pacific but decrease in the equatorial western Pacific (medium confidence) (Erauskin-Extramiana et al., 2019). In the western and central Pacific, redistribution of tropical tuna due to climate change is projected to affect license revenues from purse seine fishing and shift more fishing into high seas areas (Bell et al., 2018; Table 15.5). For the east Atlantic, observational evidence indicates that not only will tuna distribution change with temperature anomalies, but also fishing effort distribution (Rubio et al., 2020a). There is medium confidence that climate change will create new fishing opportunities when exploited fish stocks shift their distribution into new fishing regions in enclosed seas, such as the Mediterranean and the Black Sea (Hidalgo et al., 2018; Pinsky et al., 2018). However, in general, where land barriers constrain the latitudinal shifts, the expected impacts of climate change are population declines and reduced productivity (high confidence) (Oxenford and Monnereau, 2018). Besides direct impacts on the abundance of fisheries-targeted species, climate-change-induced proliferation of invasive species could also affect fisheries’ productivity (low confidence) (Mellin et al., 2016; Goldsmith et al., 2019).
Shifting marine fisheries will affect national economies (high confidence) (Bindoff et al., 2019). It has been suggested that, without government subsides, fishing is already non-profitable in 54% of the international waters (Sala et al., 2018). Projections are that fishing maximum revenue potential from landed catches will decrease further by 10.4% (±4.2%) by 2050 relative to 2000 under RCP8.5, close to 35% greater than the decrease projected for the global maximum catch potential (7.7±4.4%); (Lam et al., 2016). The global revenue potential loss for that period ranges from USD 6 to 15 billion (depending on the model), but impacts may be amplified at the regional scale for fisheries-dependent and low-income countries. The maximum revenue potential percentage decrease in the EEZ under RCP8.5 is estimated to be over 2.3 times larger than that of the high seas (Lam et al., 2016). Ocean acidification is also expected to drive large global economic impacts (medium confidence) (Cooley et al., 2015; Fernandes et al., 2017; Macko et al., 2017; Hansel et al., 2020), and there is high confidence that the integrated economic consequences of all interacting climate change-related factors would result in even larger losses. Changes in the frequency and intensity of extreme events will also alter marine ecosystems and productivity. Marine heatwaves can lead to severe and persistent impacts, from mass mortality of benthic communities to decline in fisheries catch (IPCC, 2021, Box 9.2). These events have very likely doubled in frequency between 1982 and 2016 and have also become more intense and longer (Smale et al., 2019; Laufkotter et al., 2020); for all future scenarios Earth System Models project even more frequent, intense and longer-lasting marine heatwaves (Eyring et al., 2021; IPCC, 2021, Box 9.2).
In addition to temperature and water availability stress, climate change will bring new water quality challenges in freshwater systems, including increased dissolved organic carbon and toxic metal loads (high confidence) (Chen et al., 2016). Harrod et al. (2018a) found that the two major inland fishery producers (China and India) will face significant stress in the future, a large group of countries that produce around 60% of total yield is projected to face medium stress, and a small group of 17 countries has the least severe repercussions (medium confidence). Climate warming may enhance northward colonisation of water bodies of commercial freshwater species in the Arctic, where there are few ecological competitors (medium confidence) (Campana et al., 2020) but at the same time may also accentuate the age-truncation effect of harvesting, elevating the population’s vulnerability to environmental perturbations (Smalås et al., 2019). Detailed information on many of the most important inland fisheries is limited.
In terms of food safety, major concerns linked to climate change include the continued trend of increasing HABs, and the quantity of pollutants reaching aquatic systems (Box 3.3; Section 5.11).
5.8.4 Adaptation
Adaptation options in land- and aquatic-based culturing food production systems include both governance actions and changes in the factors of production (Section 5.4.4, 5.5.4, Reverter et al., 2020). In contrast, adaptation options in fisheries are primarily concentrated in the socioeconomic dimension, especially governance and management (Brander et al., 2018; Holsman et al., 2019), and given the scale of the problem, there are relatively few intentional, well-documented examples of implemented tactical responses (Bell et al., 2020).
The proportion of fisheries operating at levels that are considered biologically unsustainable by the FAO has increased from 10% in 1974 to 34.2% in 2017 (FAO, 2020d). There is high confidence that reducing stresses on marine ecosystems reduces vulnerability to climate change and augments resilience (Barange, 2019; Woodworth-Jefcoats et al., 2019; Ogier et al., 2020). Specifically, overfishing is the most critical non-climatic driver affecting the sustainability of fisheries, and therefore improving management could help rebuild fish stocks, reduce ecosystem impacts and increase the adaptive capacity of fishing (high confidence); (Barange, 2019; Das et al., 2020). Pursuing sustainable fisheries practices under a low-emissions scenario would decrease risk by 63%; in contrast, under the most extreme RCP8.5, both profit and harvest decline relative to today even under the most optimistic assumptions about global fisheries management reforms (Gaines et al., 2018; Sumaila et al., 2019; Free et al., 2020).
One adaptation strategy in the fishing sector is developing the capacity to recognise and respond to new opportunities that might arise from climate change by establishing a policy and planning setting that augments the fishers’ flexibility to change target species of fisheries or even engage in different productive activities. A key element would be the design and implementation of management schemes that consider flexible permits, sharing quotas, rethinking boundaries and reference points in response to system changes (Brander et al., 2018; Cross-Chapter Box MOVING PLATE this chapter). Large-scale distribution and productivity changes of commercial fish species will demand the ability to implement cooperative fishing strategies (Cisneros-Montemayor et al., 2020; Østhagen et al., 2020), and adjust multi-lateral treaties and other legal instruments used for managing shared transboundary ecosystems (Butler et al., 2019; Cross-Chapter Box MOVING PLATE this chapter).
There is high confidence that making climate change and adaptive capacity a mainstream consideration in global, regional, environmental and fisheries governance structures can improve the response capacity to ocean change (Gaines et al., 2018; Bindoff et al., 2019; Holsman et al., 2020; Ojea et al., 2020). For example, spatial management that includes strategies such as Territorial Use Rights for Fishing (TURFs), locally managed marine areas (LMMAs) and customary tenure is an approach that has climate change adaptation potential in small-scale fisheries but will require adjustments in governing and managing institutions that allow them to be more dynamic and flexible (Le Cornu et al., 2018). In regions where some of these measures have already been tested, institutional, legal, financial and logistical barriers to successful adaptation have been encountered, such as market failures stemming from uncertainty around new or emerging species, or policy barriers derived from the fact that the creation of scientific information needed to change regulations is likely slower than the pace of changes in stocks (Peck and Pinnegar, 2018).
Adaptation capacity is limited by the financial capacity of some countries (Bindoff et al., 2019). For example, in West African fisheries, adaptation costs associated with replacing the loss of coastal ecosystems and productivity is estimated to require 5–10% of countries’ GDP (Zougmoré et al., 2016). For Pacific Islands and Coastal Territories, fisheries adaptation will require significant investment from local governments and the private sector (Rosegrant et al., 2016), and reducing dependence on or finding alternatives to vulnerable marine resources (Johnson et al., 2020; Mabe and Asase, 2020).
Adaptive capacity is strongly associated with social capital (i.e., the networks, shared norms, values and understandings that facilitate cooperation within or among groups) (high confidence) (Stoeckl et al., 2017; D’agata et al., 2020) and depends on to what extent stakeholders are aware of climate change and their perception of risk (Ankrah, 2018; Martins and Gasalla, 2018; Chen, 2020). Improving information flows allows for a more efficient co-management implementation (medium confidence) (Vasconcelos et al., 2020). Utilisation of local and Indigenous knowledge has the potential to facilitate adaptation (Bindoff et al., 2019), not only because it represents actual experiences and autonomous adaptations, but also because it facilitates reaching shared understanding among stakeholders and adoption of solutions. Challenges to hybridising local ecological knowledge and scientific knowledge include differences in stakeholder or governance perceptions about the validity of each knowledge set and issues of expertise and trust (Harrison et al., 2018). Engaging Indigenous Peoples and local communities as partners across climate research ensures this knowledge is utilised, enhancing the usefulness of assessments (Bindoff et al., 2019) and facilitating the co-construction and implementation of sustainable solutions (medium confidence) (Braga et al., 2020; Bulengela et al., 2020). Building climate resilience in the fishing sector also involves recognising gender and other social inequities (Call and Sellers, 2019), and ensuring that all stakeholders are equally involved in the adaptation plans, including their design and the capacity-building training programmes.
There is high confidence that, for the freshwater fisheries systems, the most immediate adaptation option is the effective linkage of fisheries management to the adaptation plans of other sectors, especially water management (hydropower, irrigation and the commitment to maintaining environmental flows) (Harrod et al., 2018a; Kao et al., 2020). In some regions, organisations are already addressing this issue; for example, The Office of Water (OW) in the USA is aimed at ensuring that drinking water is safe while ecosystem is conserved to provide healthy habitat for fish, plants and wildlife; however, success strongly depends on the possibility of integrating the jurisdictional framework of different agencies (Poesch et al., 2016), the implementation of effective monitoring programmes (Paukert et al., 2016) and finding ways to incentivise the early restoration of degraded systems (Ranjan, 2020).
Cross-Chapter Box: MOVING PLATE: Sourcing Food When Species Distributions Change
Authors: H. Gurney-Smith (Canada/UK), W. Cheung (Canada), S. Lluch Cota (Mexico), E. Ojea (Spain), C. Parmesan (France/UK/USA), J. Pinnegar (UK) P. Thornton (Kenya/UK), M-F. Racault (UK/France), G. Pecl (Australia), E.A. Nyboer (Canada), K. Holsman (USA), K. Miller (USA), J. Birkmann (Germany), G. Nelson (USA) and C. Möllmann (Germany)
This Cross-Chapter Box, the ‘moving plate’, addresses climate-induced shifts and domesticated production suitability of food species consumed by people. Marine, freshwater and terrestrial systems are already experiencing species shifts in response to climate change (very high confidence) (see also Sections 2.4.2.1. and 3.4.3., Figure MOVING PLATE.1 this chapter), with subsequent impacts on food provisioning services, pests and diseases (high confidence) (see Box 5.8 and Cross-Chapter Box ILLNESS in Chapter 2). This Box highlights food insecurity and malnutrition of vulnerable peoples under climate change for both wild and domesticated aquatic and terrestrial species, and discusses challenges for adaptation and the roles that management (transboundary and ecosystem-based) can play to enable food security, reduce conflicts and prevent resource over-extraction.
Range contractions, shifts or extirpations are projected for terrestrial and aquatic species under warming, with greater warming leading to larger shifts and losses, where mitigation would therefore benefit climate refugia and reduce projected biodiversity declines (Smith et al., 2018; Warren et al., 2018). Marine species are moving poleward faster than terrestrial and freshwater species, despite faster warming on land (Pecl et al., 2017; Lenoir et al., 2019; Woolway and Maberly, 2020), leading to new or exacerbated socioeconomic conflicts within and between countries (see Figure MOVING PLATE.1 this chapter, see Sections 13.5.2.2., 15.3.4.4., FAQ 15.3., Mendenhall et al., 2020). There is large variation in the magnitude and pattern of species shifts, even among similar species within a region, leading to changes in communities in a given region (Brown et al., 2016; Pecl et al., 2017). The number of extreme heat stress days are projected to increase for domesticated species like cattle (see Figure MOVING PLATE.1 this chapter), leading to shifts in suitable habitat for raising livestock in the open with associated impacts in animal productivity and the costs of adapting in Africa, Asia, and Central and South America (Thornton et al., 2021).
Nutritional dependency, cultural importance, livelihood, or economic reliance on shifting species will increase impacts of climate change, especially for small-scale fishers (marine and freshwater), farmers, women and communities highly dependent on local sources of food and nutrition (high confidence) (see Figures MOVING PLATE.1 and MOVING PLATE.3 this chapter, Sections 3.5.3., 8.2.1.2. and 15.3.4.4, McIntyre et al., 2016; Blasiak et al., 2017; Kifani et al., 2018; Bindoff et al., 2019; Atindana et al., 2020; Hasselberg et al., 2020; Farmery et al., 2021). Micronutrient concentrations from marine fisheries vary with species, providing higher concentrations of calcium, iron and zinc in tropical regions and higher concentrations of omega-3 fatty acids in polar regions (Hicks et al., 2019). While consumption of smaller species rich in micronutrients may provide significant benefits against deficiencies in Asia and Africa, local dietary changes in fish consumption may be linked to food preferences, fish availability due to international trade or illegal fishing and competing usage of fish (see Figure MOVING PLATE.3 this chapter, Hicks et al., 2019; Sumaila et al., 2020; Vianna et al., 2020). Industrial fleets are likely to switch target species (Belhabib et al., 2016) and inhibit small-scale fishers via illegal, unreported or unregulated fishing in EEZs (Belhabib et al., 2019; Belhabib et al., 2020). Extreme events can exacerbate issues, as fisheries are frequently increasingly exploited as a coping mechanism under times of crisis, increasing illegal fishing activities and conflict among maritime users (Pomeroy et al., 2016; Mazaris and Germond, 2018). Spatial conflicts between artisanal and commercial foreign fishing fleets are already occurring in Ghana (Penney et al., 2017), and from climate-induced tropical tuna shifts in the Western and Central Pacific Ocean Islands (see Section 15.3.4.4., (Bell et al., 2018 a)). Properly managed small-scale fisheries can reduce poverty and improve localised food security and nutrition in low-income countries but will likely require restriction in the number of fishers, boat size or fishing days (Purcell and Pomeroy, 2015; Hicks et al., 2019).
Shifting species have negative implications for the equitable distribution of food provisioning services, increasing the complexity of resolving sovereignty claims and climate justice (high confidence) (Allison and Bassett, 2015; Ayers et al., 2018; Baudron et al.; Ojea et al., 2020; Palacios-Abrantes et al., 2020). Higher-latitude countries generally have higher GHG emissions and will benefit from poleward-migrating resources from tropical poorer and lower-emitting GHG countries (Free et al., 2020). In this context, climate justice supporting fishing arrangements could offset socioeconomic impacts from exiting species (Mills, 2018; Lam et al., 2020) and have negative implications particularly for small-scale operators (Farmery et al., 2021), However, considerations of climate justice have not been used by Regional Fisheries Management Organizations (RFMOs) allocation shares to date (Engler, 2020). Species shifting from one historical jurisdiction to another may result in an incentivised depletion of the resource by the country the stock is shifting away from; reforming management to allocate resource sharing of quotas and permits or stock-unrelated side payments in bilateral or multilateral cooperative agreements may compensate or prevent loss (Diekert and Nieminen, 2017; Free et al., 2020; Ojea et al., 2020; Østhagen et al., 2020; Cross-Chapter Paper Polar 6.2.).
Strong governance, ecosystem-based and transboundary management are considered fundamental to ameliorate the impacts of climate change (high confidence) but may be limited in effectiveness by the magnitude of change projected under low or no mitigation scenarios (see Sections 2.6.2., 14.4.2.2. and 15.3.4.4., Harrod et al., 2018c; Pinsky et al., 2018; Holsman et al., 2020; Ojea et al., 2020). Flexible and rapid policy reform and management adaptation will help to meet sustainability targets (Nguyen et al., 2016; Pentz and Klenk, 2020), and may only be available for countries with the scientific, technical and institutional capacity to implement these (high confidence) (Peck and Pinnegar, 2018; Figures MOVING PLATE.2 and 3 this chapter). Other adaptation options include ‘follow the food’ thereby migrating further (Belhabib et al., 2016), provision of alternative livelihoods (Thiault et al., 2019; Cross-Chapter Box MIGRATE in Chapter 7, Free et al., 2020), increasing ecosystem resilience by rebuilding coastal mangroves (Tanner et al., 2014; and Box 1.3) and riparian areas of freshwater ecosystems (Mantyka-Pringle et al., 2016) and autonomous adaptations, such as harvesting gear modifications to access new target species (Harrod et al., 2018c; Kifani et al., 2018), practice change, and early-warning systems (see Section 11.3.2.3; Pecl et al., 2019; Melbourne-Thomas et al., 2021). Adaptive capacity will change with country, region, scale (commercial, recreational, Indigenous) of fishery, jurisdiction, and resource dependence (see Figure MOVING PLATE.2 this chapter for adaptation options for marine, freshwater and terrestrial systems). While shifting fishing fleets or herding may be an adaptation option to follow resources, limits to feasibility include institutional, legal, financial and logistical barriers such as costs of sourcing food and operational economic viability (Belhabib et al., 2016); this could potentially lead to maladaptation through increased GHG emissions from fuel usage and cultural displacement from traditional fishing and herding lands. Overall, decreases in GHG emissions under future scenarios would reduce increases in global temperatures and limit species shifts, thereby lowering the likelihood of conflicts and food insecurity (high confidence).
Cross-Chapter Box: MOVING PLATE
Coastal Regions of the Gulf of Guinea: Ghanian Fisheries
Marine fisheries in Ghana are dominated by artisanal fishers with overfished stocks, high nutritional fish dependency, high illegal fishing, low governance capacity (−0.21 2018, (World Bank, 2019)) and low climate awareness in regional fisheries management (Figure MOVING PLATE.3 this chapter; see Chapter 9; Nunoo et al., 2014; Belhabib et al., 2015; Belhabib et al., 2016; Kifani et al., 2018; Belhabib et al., 2019). Artisanal fishing plays a pivotal role in reducing poverty and food insecurity, and the impacts of climate change will risk developing poverty traps (see Section 8.4.5.6., (Kifani et al., 2018)). Climate change induced species redistribution is a large risk to Ghanian fisheries, with projections of over 20 commercial fish species exiting the region with no new species entering under RCP4.5 by 2100 (Oremus et al., 2020), and has already seen increases in warmer-water species with declining stocks. Adaptation options being applied are extending fishing ranges, increasing fishing effort (and cost) to access declining fish (with government fuel incentives) (Kifani et al., 2018; Muringai et al., 2021), developing aquaculture for alternative livelihoods, implementing fleet monitoring to reduce illegal fishing, and developing a robust Fisheries Information and Management System that accounts for environmental and climate drivers (Johnson et al., 2014; FAO, 2016; Kassi et al., 2018). However, fisheries remain insufficiently regulated, there is a lack of a skilled workforce, and there is low access to credit; collectively, these factors limit options for artisanal fishers to find alternative sustainable employment (FAO, 2016).
Shifting Distributions of Freshwater Fishery Resources: Knowledge Gaps
Freshwater fisheries provide the primary source of animal protein and essential micronutrients for an estimated 200 million people globally and are especially important in tropical developing nations (see Section 9.8, Lynch et al., 2017; Funge-Smith and Bennett, 2019.). There is evidence that freshwater fishes have undergone climate-induced distribution shifts (Comte and Grenouillet, 2015; see Section 9.8.5.1.), and further shifts are projected as water temperatures rise and hydrological regimes change, with the largest effects predicted for equatorial, subtropical and semi-arid regions (Barbarossa et al., 2021). Currently, the effects of distribution shifts on local fishery catch potential, food security and/or nutrition have not been quantified for any major inland fishery, representing a key knowledge gap for anticipating future adaptation needs for freshwater fishing societies. However, studies on fishers’ perceptions of climate-induced changes in fishery catch rates have revealed that using local knowledge to adjust management practices (see Chapter 12 Central and South America this volume; Oviedo et al., 2016) and shifting gears, fishing grounds and target species (see Section 9.8.5.3.; Musinguzi et al., 2016) can be effective adaptation options.
Terrestrial Species Shifts
There is robust evidence of shifts that terrestrial species have shifted poleward in high latitudes, with general declines of sea-ice dependent as well as some extreme-polar-adapted species (high confidence) (Arctic and Siberian Tundra, see Section 2.4.2.2., Cross-Chapter Paper 6), with often deleterious effects on the food security and traditional knowledge systems of Indigenous societies (Horstkotte et al., 2017; Pecl et al., 2017; Mallory and Boyce, 2018; Forbes et al., 2020). Recent decades have seen declines in Arctic reindeer and caribou (see Section 2.5.1., Cross-Chapter Paper 6), and adaptation responses include utilisation of Indigenous knowledge with scientific sampling to maintain traditional management practices (Pecl et al., 2017; Barber et al.; Forbes et al., 2020). Preserving herder livelihoods will necessitate novel solutions (supplementary feeding, seasonal movements), where governance, ecological and socioeconomic trade-offs will be balanced at the local level (Horstkotte et al., 2017; Pecl et al., 2017; Mallory and Boyce, 2018; Forbes et al., 2020). Wild meat consumption plays a critical, though not well understood, role in the diets and food security of several hundred million people (medium evidence), for example in lower latitudes such as Central Africa and the Amazon basin (Bharucha and Pretty, 2010; Godfray et al., 2010; Nasi et al., 2011; Friant et al., 2020). Although illegal in many countries, wild meat hunting occurs either in places where there is no or limited domesticated livestock production, or in places where shock events such as droughts and floods threaten food supply, forcing increased reliance on wild foods including bush meat (Mosberg and Eriksen, 2015; Bodmer et al., 2018). Appropriate management of wild meat for reliant peoples under projected climate change will necessitate incorporating social justice elements into conservation and public health strategies (see Cross-Chapter Box ILLNESS in Chapter 2, Cross-Chapter Box COVID in Chapter 7, Friant et al., 2020; Ingram, 2020; Pelling et al., 2021).
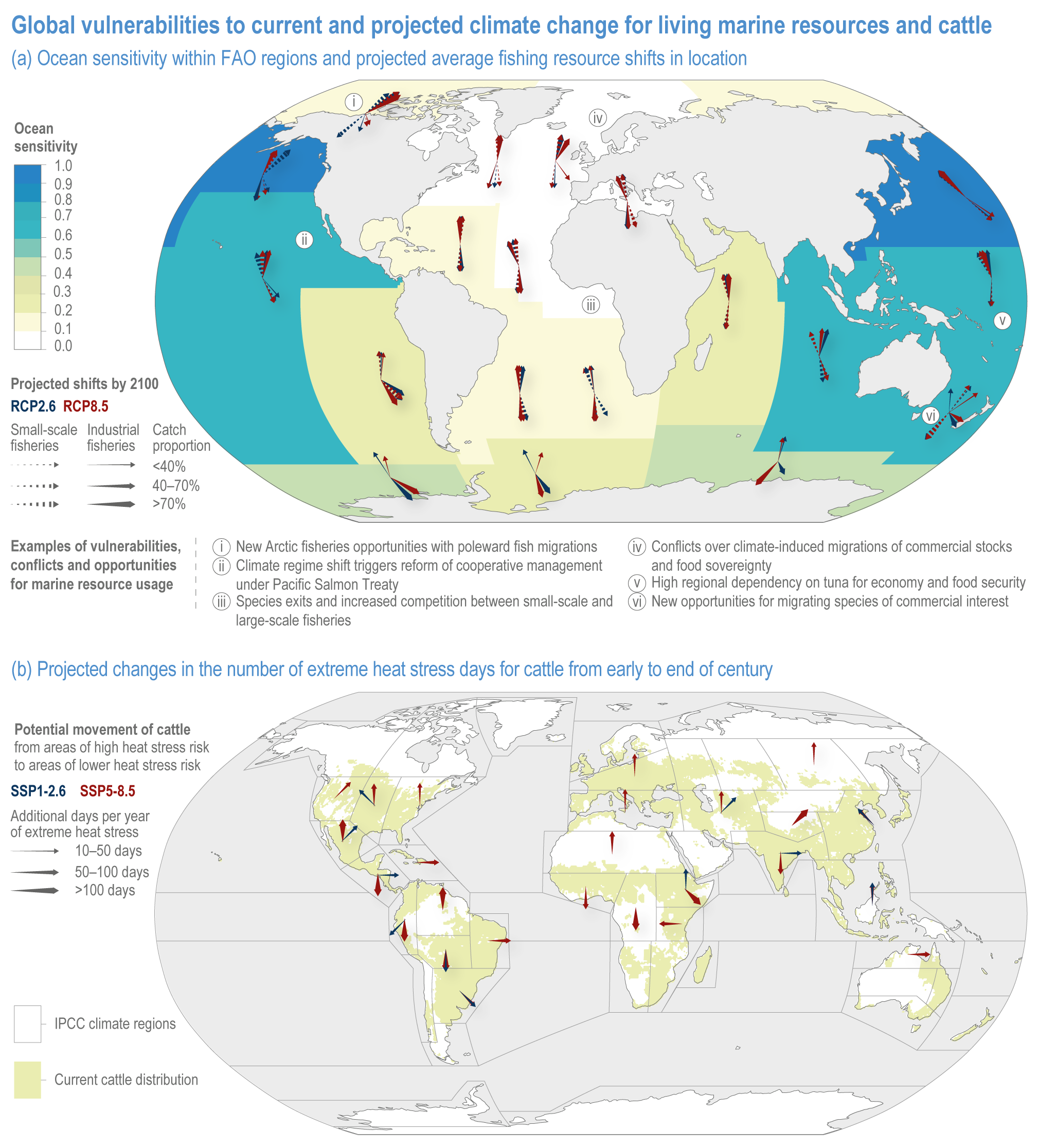
Figure Cross-Chapter Box MOVING PLATE.1 | Global vulnerabilities to current and projected climate change for living marine resources and cattle.
(a) Ocean areas are delineated into FAO (Food and Agricultural Organization of the United Nations) regions. Ocean sensitivity is calculated from aggregated sensitivities from Blasiak et al. (2017) S1 country data based on number of fishers, fisheries exports, proportions of economically active population working as fishers, total fisheries landings and nutritional dependence, which was subsequently re-analysed for each FAO region depicted here. Arrows denote projected average commercial and artisanal fishing resource shifts in location under RCP2.6 and under RCP8.5 (dark-blue and red arrows, respectively) scenarios by 2100. Text boxes highlight examples of vulnerabilities (Bell et al., 2018 a), conflicts (Miller et al., 2013; Blasiak et al., 2017; Østhagen et al., 2020) or opportunities for marine resource usage (Robinson et al., 2015; Stuart-Smith et al., 2018; Meredith et al., 2019).
(b) Projected changes in the number of extreme heat stress days for cattle from early (1991–2010) to end of century (2081–2100) under SSP1-2.6 and SSP5-8.5, shown as arrows rooted in the most affected area in each IPCC sub-region pointing to the nearest area of reduced or no extreme heat stress. Arrows are shown only for sub-regions where >1 million additional animals are affected. Areas in green are those with >5000 animals per 0.5° grid cell in the eary 21st century (Thornton et al., 2021).
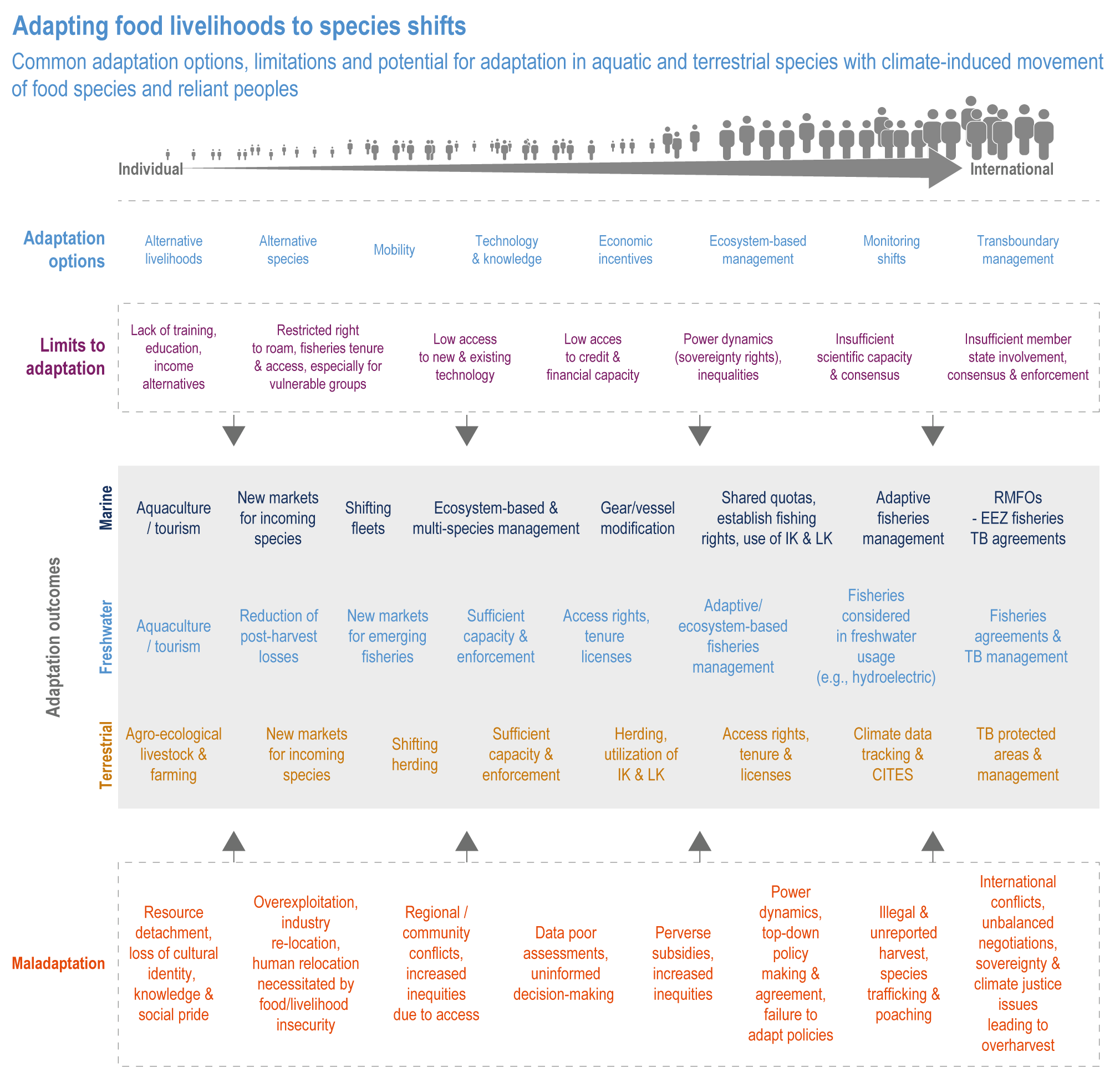
Figure Cross-Chapter Box MOVING PLATE.2 | Common adaptation options, limitations and potential for adaptation and maladaptation in aquatic and terrestrial species with climate-induced movement of food species and reliant peoples.
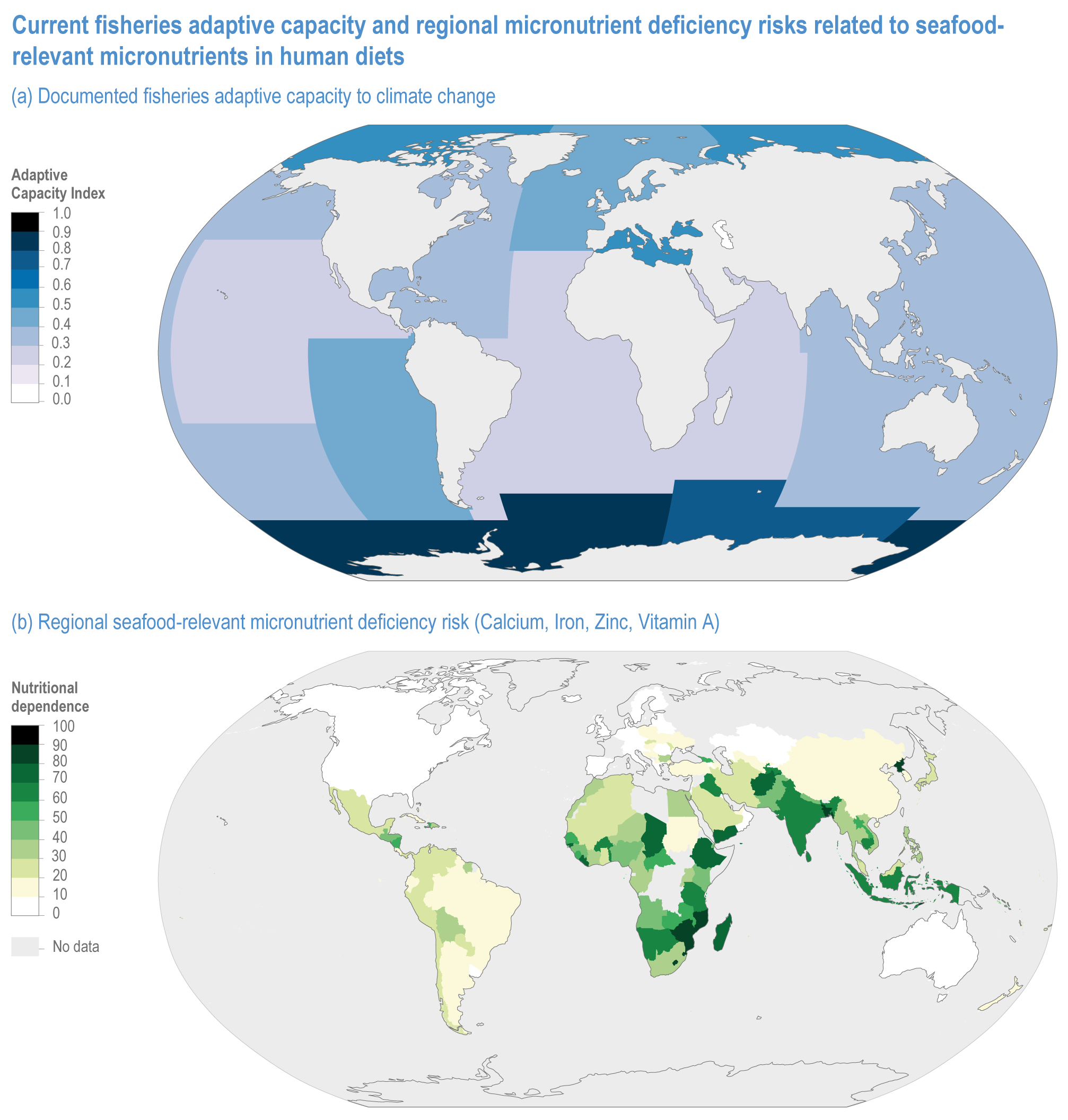
Figure Cross-Chapter Box MOVING PLATE.3 | Global documented fisheries adaptive capacity to climate change and regional seafood micronutrient deficiency risk. Ocean areas are delineated into FAO (Food and Agricultural Organization of the United Nations) regions. Fisheries management adaptive capacity is a function of: averaged GDP World Development Indicators for 2018 (World Bank, 2020); climate awareness assessments of 30 of the FAO recognised most recent RFMOs with direct fisheries linkages (see Supplementary Material SM5.5); governance effectiveness index based on six aggregate indicators (voice and accountability, political stability and absence of violence/terrorism, government effectiveness, regulatory quality, rule of law, control of corruption) from 2018 World Governance Indicator (World Bank, 2019) data; and heterogeneity of countries within each FAO zone (highly heterogeneous regions are less likely to establish sustainable and efficient fisheries management for the entire FAO zone). Land area represents the percentage regional averaged seafood micronutrient deficiency risk of calcium, iron, zinc and vitamin A from 2011 data (Beal et al., 2017).
In terrestrial, marine and freshwater systems, human populations already impacted by poverty and hunger experience greater risk under climate change. Future food security will depend on access to other sustainable sources either via transnational agreements or resource/livelihood diversification. Sudden shocks across food production systems (Cottrell et al., 2019) can lead to increases in fisheries harvest and wild meat consumption, and following food species may result in community relocations or disruption and loss of access to historical places of attachment (high confidence) (Pecl et al., 2017; Lenoir et al., 2019; Meredith et al., 2019; Melbourne-Thomas et al., 2021; see Cross-Chapter Box MIGRATE in Chapter 7). Ecosystem-based management approaches exist for terrestrial, marine and freshwater systems, but have proved successful only with early engagement of local small-scale, subsistence fishers/harvesters, utilising Indigenous knowledge and local knowledge and needs, in addition to those of larger-scale operators (high confidence) (Huntington et al., 2015; McGrath and Costello, 2015; Huq and Stubbings, 2016; Huq et al., 2017; Raymond-Yakoubian et al., 2017; Nalau et al., 2018; Raymond-Yakoubian and Daniel, 2018; Pecl et al., 2019; Planque et al., 2019). Currently, there are large regional differences in climate literacy in RFMOs (Sumby et al., 2021) which, when combined with low governance and GDP per capita, will limit adaptation capacity and increase vulnerabilities, particularly for tropical and subtropical regions already at increased risk due to poleward species migrations (see Figure MOVING PLATE.3 this chapter). Trade will be an alternative to compensate for the moving plate but has specific risks that can amplify inequities and maladaptation (Asche et al., 2015; Vianna et al., 2020).
Cross-Chapter Box: MOVING PLATE
5.9 Ocean-Based and Inland Aquaculture Systems
Global aquaculture provides more fish for human consumption than wild capture fisheries, with projected provisioning of 60% by 2030 (FAO, 2018c). Aquaculture can contribute to SDGs by reducing poverty and food insecurity, filling increasing aquatic food demand shortages from declining capture fisheries production (medium confidence) (Figure 5.13a and c, World Bank, 2013; Béné et al., 2016; Hambrey, 2017; Beveridge et al., 2018b; Kalikoski et al., 2018; Belton et al., 2020) and improving social inequities for poor rural communities (Béné et al., 2016; FAO, 2018c; Vannuccini et al., 2018; Pongthanapanic et al., 2019). Global aquaculture production reached 82 million tonnes (Mt) of food fish, crustaceans, molluscs and other aquatic animals from inland (51 Mt) and marine (31 Mt) systems, and 32 Mt of aquatic plants in 2018 (FAO, 2020d). China, India, Indonesia, Vietnam, Bangladesh, Egypt, Norway and Chile are major production regions (FAO, 2020d). The range of species, farming methods and environments makes aquaculture the most diverse, long-standing farming practice in the world, with an estimated global sectoral value of USD 250 billion in 2018 (Figure 5.13b and 5.14d, Bell et al., 2019; Harland, 2019; FAO, 2020d; Houston et al., 2020; Metian et al., 2020), but it is dominated by 20 finfish, 9 mollusc and 6 crustacean species (FAO, 2020). Inland aquaculture in freshwater and coastal ponds accounts for 85–90% of farmed production (Beveridge et al., 2018b; Naylor et al., 2021). Globally, 20.5 million people are engaged in aquaculture (FAO, 2020d), where marine finfish farming is primarily conducted by high-income countries and inland production is dominated by small-scale producers in lower-middle-income countries (Vannuccini et al., 2018).
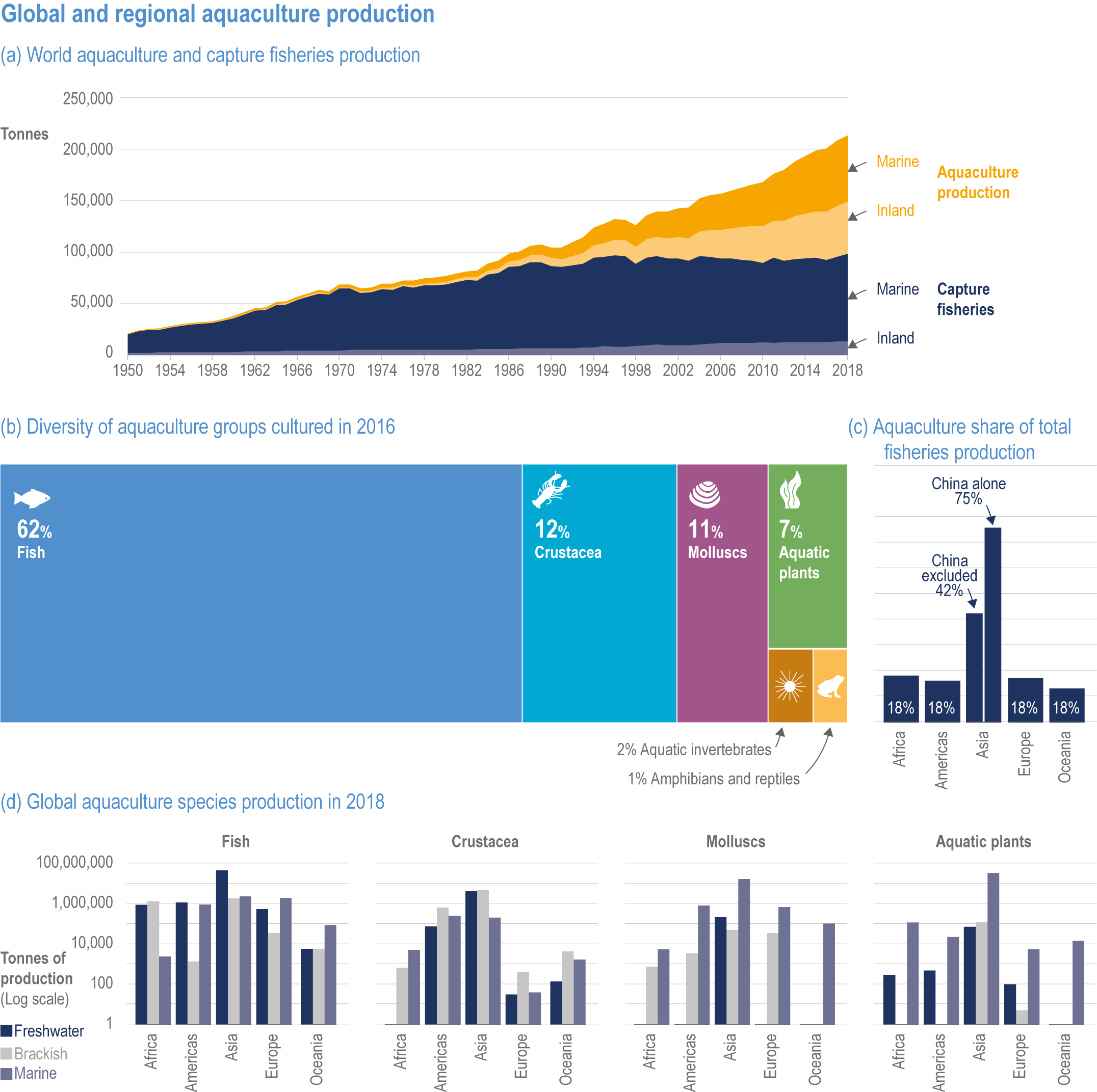
Figure 5.13 | Global and regional aquaculture production. (a) World wild capture fisheries and aquaculture inland (freshwater and brackish) and marine production from 1950 to 2018; (b) diversity of aquaculture groups cultured in 2016; (c) regional aquaculture share of total fisheries production; and (d) global aquaculture species production in 2018 by region and type (freshwater, brackish or marine) on a logged scale (FAO, 2018c; FAO, 2020c; FAO, 2020d).
5.9.1 Observed Impacts
Marine aquaculture food production is being impacted directly and indirectly by climate change (high confidence) (Bindoff et al., 2019). Ocean pH and oxygen levels are declining, whereas global warming, sea level rise and extreme events are increasing (Cross-Chapter Box SLR in Chapter 3, Canadell et al., 2021; Eyring et al., 2021; Fox-Kemper et al., 2021; Lee et al., 2021;). Marine heatwaves have been increasing in both incidence and longevity over the past century (Frolicher and Laufkotter, 2018; Oliver et al., 2018; Bricknell et al., 2021), with productivity consequences for marine aquaculture (mariculture), carbon sequestration and local species extinctions (high confidence) (Weatherdon et al., 2016; Smale et al., 2019). Temperature increases related to El Niño climatic oscillations have caused mass fish mortalities either through warming waters (e.g., Pacific threadfin in Hawaii (McCoy et al., 2017)) or associated HABs (e.g., 12% loss of Atlantic salmon as well as other fish and shellfish in Chile in 2016, with estimated USD 800 million in losses (high confidence) (Clement et al., 2016; Apablaza et al., 2017; Leon-Munoz et al., 2018; Trainer et al., 2020)). Increases in sea lice parasite infestations on salmon are related to higher salinity and warmer waters (medium confidence) (Groner et al., 2016; Soto et al., 2019). Ocean acidification is having negative impacts on the sustainability of mariculture production (high confidence) (Bindoff et al., 2019), with observed impacts on shellfish causing significant production and economic losses for regions, estimated at losses of nearly USD 110 million by 2015 in the Pacific Northwest (Barton et al., 2015; Ekstrom et al., 2015; Waldbusser et al., 2015; Zhang et al., 2017b; Doney et al., 2020). Ocean oxygen levels are declining due to climate change (Hoegh-Guldberg et al., 2018; IPCC, 2021), and decreased oxygen (hypoxia) has negative impacts on fish physiology (Cadiz et al., 2018; Hvas and Oppedal, 2019; Martos-Sitcha et al., 2019; Perera et al., 2021), fish growth, behaviour and sensitivity to concurrent stressors (high confidence) (Stehfest et al., 2017; Abdel-Tawwab et al., 2019).
Observed impacts on inland systems have generally been site and region specific (high confidence) (Hoegh-Guldberg et al., 2018; Sainz et al., 2019; Lebel et al., 2020). Salinity intrusions into freshwater aquaculture systems have changed oxygen and water quality of inland ponds, resulting in mortalities in areas such as India and Bangladesh (medium confidence) (Dubey et al., 2017; Dabbadie et al., 2018). Rapid changes in temperature, precipitation, droughts, floods and erosion have created significant production losses for aquatic farmers in Cambodia, Laos, Myanmar, Thailand, Viet Nam and Ghana (medium confidence) (Asiedu et al., 2017; Pongthanapanic et al., 2019; Lebel et al., 2020). Algal blooming and inland lake browning related to warming was found to negatively affect fish biomass (van Dorst et al., 2018). Observed indirect effects of climate change on aquaculture include extreme weather events that damage coastal aquaculture infrastructure or enable flooding, both leading to animal escapees (e.g., fish, shrimp), damaged livelihoods and interactions with wild species (high agreement , medium evidence) (Beveridge et al., 2018b; Dabbadie et al., 2018; Kais and Islam, 2018; Pongthanapanic et al., 2019; Ju et al., 2020).
5.9.2 Assessing Vulnerabilities
Aquaculture vulnerability assessments have shown that countries from both high and low latitudes are highly vulnerable to climate change, where vulnerability is driven by particular exposures, economic reliance, type of production sector (freshwater, brackish, marine) and adaptive capacity (high confidence) (Handisyde et al., 2017; Soto et al., 2018). Regional aquaculture vulnerabilities and risk mitigation potentials for the major FAO reporting regions are shown in Figure 5.14. Best practice guidelines for assessments exist (Brugère et al., 2019; FAO, 2020d), but in practice most only cover some climatic drivers (medium agreement , limited evidence) (Soto et al., 2018). Holistic vulnerability assessments include ecosystem services (Custódio et al., 2020; Gentry et al., 2020) and farming practices which can exacerbate production pressures (stocking densities, eutrophication, fish stress) (Soto et al., 2018; Sainz et al., 2019). Common vulnerabilities to inland and marine aquaculture include increasing incidence and toxicity of HABs related to warming waters, causing fish kills and product consumption risks, negatively impacting the productivity and stability of production sectors and reliant communities (high confidence) (Soto et al., 2018; Aoki et al., 2019; Bannister et al., 2019).
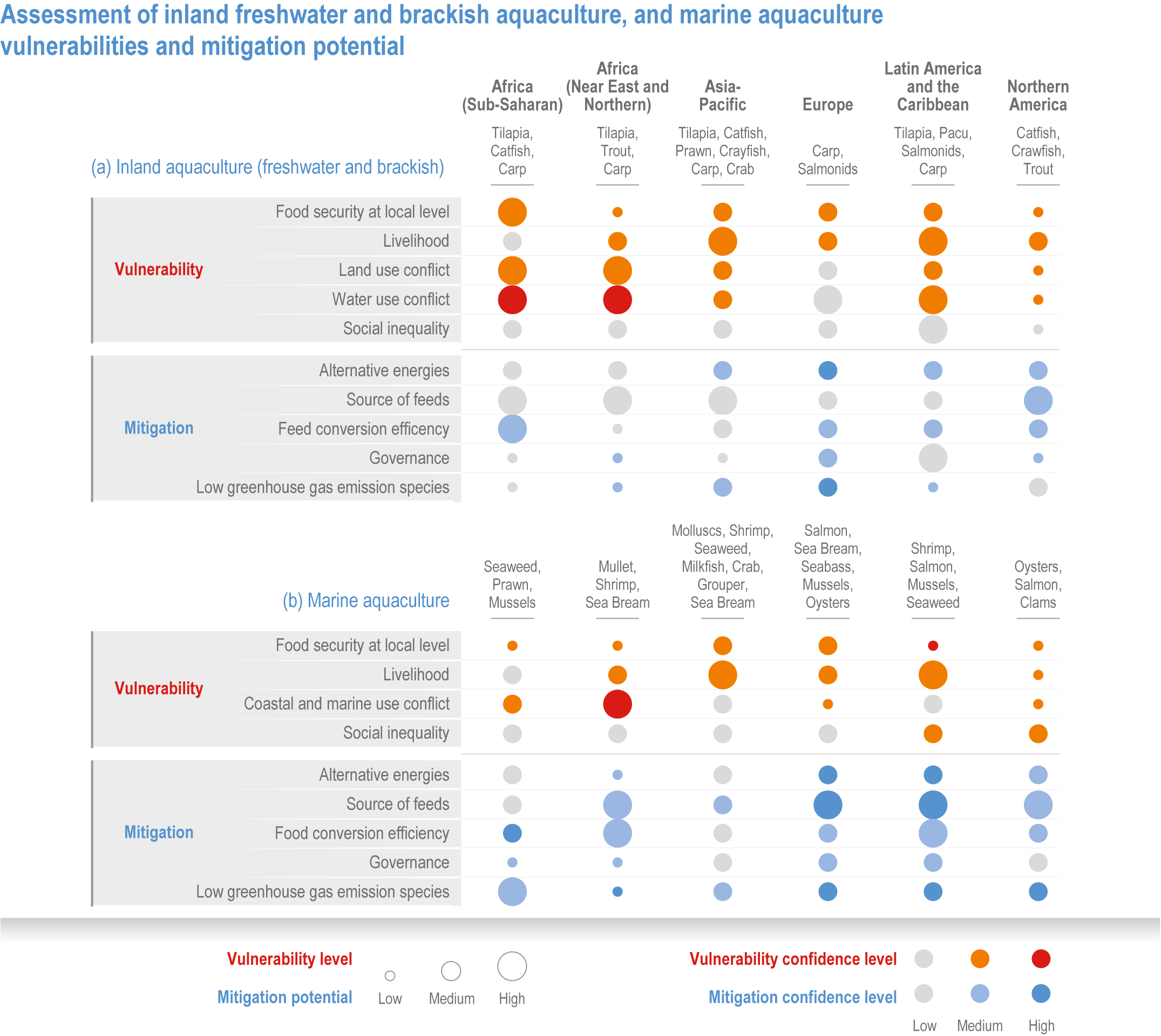
Figure 5.14 | Assessment of inland freshwater and brackish aquaculture (salinities of <10 ppm and/or no connection to the marine environment) (a) and marine aquaculture vulnerabilities and mitigation potential per major FAO production zones (b). See SM5.6 (Tables SM5.5, 5.6, 5.9, 5.10) for assessment methodologies.
There is high confidence that inland aquaculture in Southeast Asia is highly vulnerable to climate change, due to fluctuations in water resources either through climatic variability in precipitation, flooding or salinity inundation or through competition (Handisyde et al., 2017; Nguyen et al., 2018; Soto et al., 2018; Islam et al., 2019; Nguyen et al., 2019b; Prakoso et al., 2020). Studies in Bangladesh and Indonesia highlighted regional and species-specific vulnerabilities (Prakoso et al., 2020) and roles of governance in vulnerability reduction (Islam et al., 2019).
In the marine sector, vulnerability models (Brugère and De Young, 2015; Handisyde et al., 2017) have been adapted and applied to semi-quantitative spatial risk assessments for Chilean Atlantic salmon, where analysis of exposure threat coupled with mortality and temperature farm data could enhance salmon production (Soto et al., 2019). Vulnerability assessments in Korea (RCP8.5 temperature increase of 4–5°C by 2100) (Kim et al., 2019a) and the USA (ocean acidification, Barton et al., 2015; Ekstrom et al., 2015) found major exposure-related vulnerabilities for seaweeds and shellfish, with reduced vulnerabilities under higher production control and adaptive capacity. Global bivalve vulnerability assessments (RCP8.5 by 2100) show high vulnerabilities for major producing countries related to cyclones (China, Japan, South Korea, Thailand, Viet Nam and North Korea), regional risk of high sensitivity and low adaptive capacity (Chile, Peru, Spain, Italy), with few major producers (France, the Netherlands and USA) anticipated to remain moderately vulnerable by 2100 (Stewart-Sinclair et al., 2020).
Climate uncertainty and data limitations hinder vulnerability assessments (high confidence), so broader vulnerabilities and qualitative assessments can be used (Brugère and De Young, 2015; Soto et al., 2018; Brugère et al., 2019; Cochrane et al., 2019). Filling data gaps with monitoring (high confidence), increasing governmental support to assist particularly vulnerable small- and medium-scale farmers with increased costs associated with risk management and uncertainty (medium confidence) and the early inclusion of community stakeholders (high agreement , medium evidence) can reduce vulnerabilities (Handisyde et al., 2017; Dabbadie et al., 2018; Soto et al., 2018; Bindoff et al., 2019; Cochrane et al., 2019).
5.9.2.1 Gender and other social vulnerability and roles in aquaculture
There are regional differences in women’s roles, responsibilities and involvement in adaptation strategies in the aquaculture sector. Women comprise 14% of the 2018 global aquaculture workforce of 20.5 million (FAO, 2020c), representing up to 42% of the salmon workforce in Chile (Chávez et al., 2019), predominantly in processing roles (Gopal et al., 2020). In the majority of lower-middle-income countries, seaweed culture is dominated by women in family-owned businesses as in Zanzibar and the Philippines (Brugere et al., 2020; Ramirez et al., 2020), where women are not always paid directly but contribute to family incomes (high confidence) (Msuya and Hurtado, 2017; Brugere et al., 2020; Ramirez et al., 2020). In India, women collect stocking juveniles and assist in pond construction; in Bangladesh, women do the same tasks as men; and in Ghana, women undertake post-harvest fishing activities (Lauria et al., 2018). Women employed in aquaculture cooperatives gained adaptive capacity, which reduced gender inequities (medium confidence) (Farquhar et al., 2018; Gonzal et al., 2019), but lack of financial access for women can create gender inequity at larger commercial scales (Gurung et al., 2016; Call and Sellers, 2019). Women in aquaculture experience competing roles between employment, childcare and home duties (high confidence) (Morgan et al., 2015; Lauria et al., 2018; Chávez et al., 2019; see Cross-Chapter Box GENDER in Chapter 18) and differ from men in terms of perceptions of environmental risk, climate change and adaptation behaviour, with limited contributions to decision making (medium confidence) (Barange and Cochrane, 2018). Therefore, effective climate aquaculture adaptation options need to address gender inequity, such as suitable technology designs that fit with social norms and access to credit to facilitate independent uptake (medium evidence, high agreement ) (Morgan et al., 2015; Oppenheimer et al., 2019). Generalised best practices for gender-sensitive approaches to adaptation are relevant for aquaculture (UNFCCC, 2013).
5.9.3 Projected Impacts
Projected impacts on regional inland and marine aquaculture production are summarised in Figure 5.15.
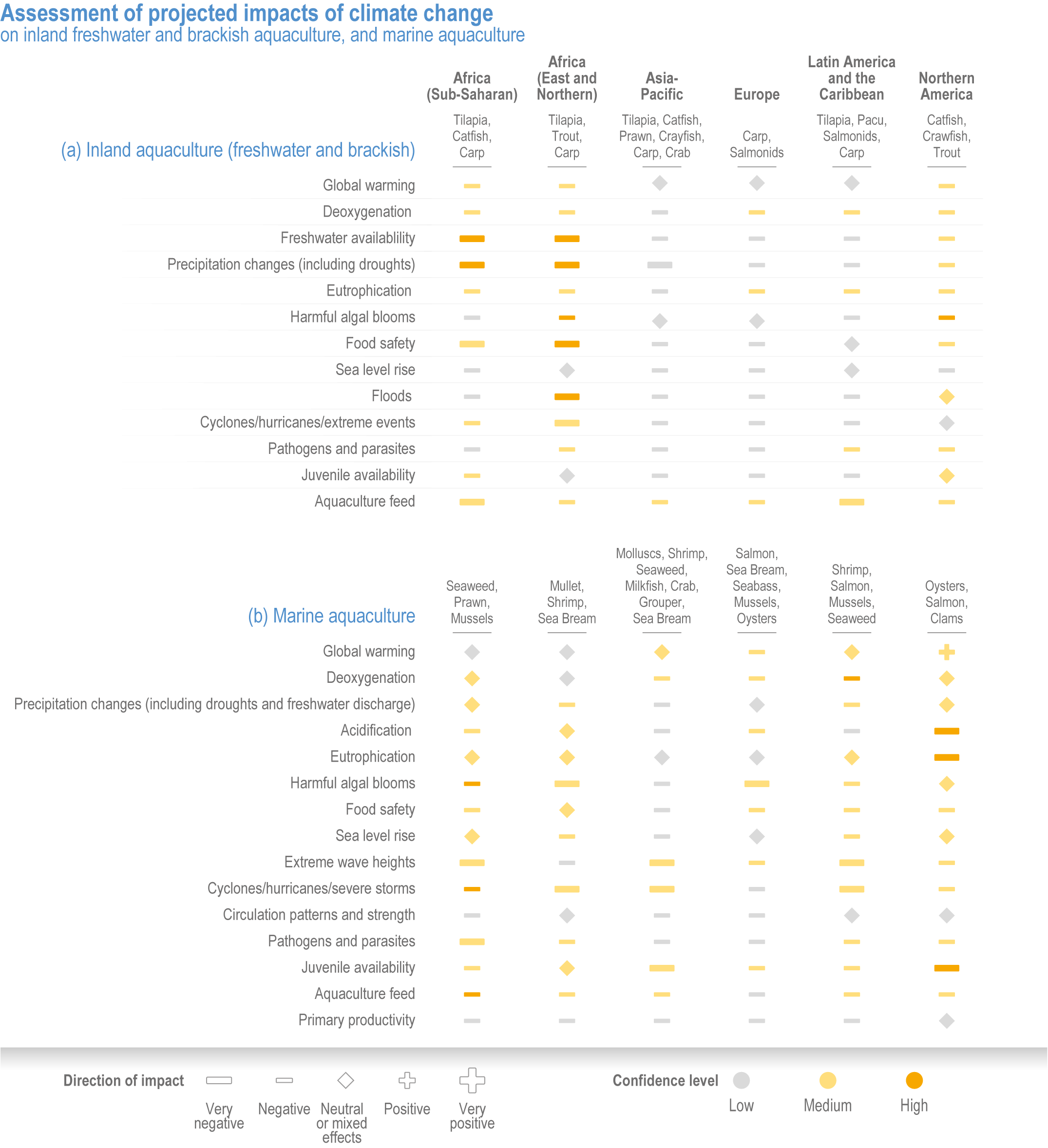
Figure 5.15 | Assessment of projected impacts of climate change on inland freshwater and brackish aquaculture (salinities of <10 ppm and/or no connection to the marine environment) (a) and marine aquaculture (b) per major FAO production zones. See SM5.6 (Tables SM5.7, 5.11) for assessment methodologies.
5.9.3.1 Inland freshwater and brackish aquaculture
Predicted sea level and temperature rise will result in coastal inundation into brackish and inland aquaculture systems (high confidence) (Mehvar et al., 2019; Nhung et al., 2019; Oppenheimer et al., 2019; Fox-Kemper et al., 2021), with negative impacts on aquaculture production in Viet Nam, East Africa and Jamaica (medium confidence) (Lebel et al., 2018; Nguyen et al., 2018; Bornemann et al., 2019). Precipitation and temperature changes will cause drought and flooding, negatively affecting near-shore fishpond productivity (limited evidence) (Canevari-Luzardo et al., 2019), but provide competitive advantages to non-native shrimp in Australia (limited evidence) (Cerato et al., 2019). Warming and acidification will increase HAB toxicity in freshwater systems, but responses may be strain-specific (Griffith and Gobler, 2020; Hennon and Dyhrman, 2020). As for molluscs in marine systems, projected climate change in freshwater and brackish systems may limit the availability of wild-sourced juveniles from fisheries (Beveridge et al., 2018). Projected impact studies for the inland and small-scale aquatic sectors are very limited (Halpern et al., 2019; Galappaththi et al., 2020b); therefore, this is a noted knowledge gap.
5.9.3.2 Marine Aquaculture
5.9.3.2.1 Finfish culture
Global projections of ocean warming, primary productivity and ocean acidification predict suitable habitat expansions and short-term growth benefits for finfish aquaculture for some regions (medium confidence) (see Figure 5.15) until thermal tolerances or productivity constraints are exceeded by 2090 (Beveridge et al., 2018b; Dabbadie et al., 2018; Froehlich et al., 2018a; Catalán et al., 2019; Thiault et al., 2019; Falconer et al., 2020a). Sensitivities for marine finfish may be high even under +1.5–2.0°C (medium confidence) (Gattuso et al., 2018), resulting in finfish farms moving northward to maintain productivity (e.g., Arctic (Troell et al., 2017)). Downscaled projections of regionally specific tolerances (Klinger et al., 2017) may be particularly useful for management and planning; a 0.5°C rise is predicted for Chilean salmon aquaculture (Soto et al., 2019), and potential negative impacts on productivity in Norway by 2029 have been projected (limited evidence) (Falconer et al., 2020a). Marine heatwaves are predicted to increase in occurrence, intensity and persistence under RCP4.5 or RCP8.5 by 2100 (Oliver et al., 2019; Bricknell et al., 2021), with risk partly mitigated by husbandry (medium confidence) (McCoy et al., 2017). Generally, negative impacts are predicted for marine species, with residual risk increasing with level of exposure (Sara et al., 2018; Smale et al., 2019), where warming will affect oxygen solubility and reduce salmon culture capacity (limited evidence) (Aksnes et al., 2019, Chapter 3) and combine with increasing incidence of HABs (high confidence) resulting in negative impacts for food security and nutrition and health (Oppenheimer et al., 2019; Colombo et al., 2020; Glibert, 2020; Raven et al., 2020). Climate change is predicted to affect the incidence, magnitude and virulence of finfish disease such as Vibriosis (Barber et al., 2016; Mohamad et al., 2019a; Mohamad et al., 2019b), but specific host–pathogen–climate relationships are not yet established (high confidence) (Slenning, 2010; Marcogliese, 2016; Montanchez et al., 2019; Bandin and Souto, 2020; Behringer et al., 2020; Filipe et al., 2020; Montanchez and Kaberdin, 2020). Projected climate change will also increase competition for feed ingredients between aquatic and terrestrial animal production systems (see Section 5.13.2.).
5.9.3.2.2 Shellfish culture
Globally, there is overall high confidence that suitable shellfish aquaculture habitat will decline by 2100 under projected warming, ocean acidification and primary productivity changes, with significant negative impacts for some regions and species before 2100 (Table 5.9, Froehlich et al., 2018a; Ghezzo et al., 2018). Shellfish growth will increase with warming waters until tolerances are reached, such as through extreme El Niño events (high confidence) (Beveridge et al., 2018b; Dabbadie et al., 2018; Liu et al., 2018b; Liu et al., 2020). Rising temperatures and ocean acidification will result in losses of primary productivity and farmed species from tropical and subtropical regions, and gains in higher latitudes (high confidence) (Froehlich et al., 2018a; Aveytua-Alcazar et al., 2020; Chapman et al., 2020; Des et al., 2020; Oyinlola et al., 2020), but net marine production gains could be achieved under strong mitigation (Thiault et al., 2019). Shellfish Vibrio infections will increase with warming waters and extreme events, increasing shellfish mortalities (medium confidence) (Green et al., 2019; Montanchez et al., 2019), with ocean acidification impairing immune responses (limited evidence) (Cao et al., 2018b). Bivalve larvae are known to be highly vulnerable to ocean acidification (high confidence) (see Section 3.3, Bindoff et al., 2019), with projected regional and species-specific levels of impact (high confidence) (Ekstrom et al., 2015; Zhang et al., 2017b; Mangi et al., 2018) (Greenhill et al., 2020). Ocean acidification is also projected to weaken shells, affecting productivity and processing (high confidence) (Martinez et al., 2018; Cummings et al., 2019) and dependent livelihoods (Doney et al., 2020).
5.9.3.2.3 Aquatic plant culture
There is medium confidence that cultivated seaweeds are predicted to suffer habitat loss resulting in population declines and northward shifts (Table 5.11).
Table 5.11 | Projected impacts of climate on specific inland, brackish and marine culture systems and species.
Exposure | Scenario | Region | Production system | Species | Impact | Reference |
Temperature increase | RCP4.5 and RCP8.5 by 2050 | Northern Thailand | Inland | Nile tilapia | Reduced productivity | |
Precipitation change (drought, hurricane, heavy rainfall) | – | Jamaica | Inland | Tilapia | Reduced productivity, infrastructure damage | Canevari-Luzardo et al. (2019) |
Temperature increase | 4°C increase, B2, A1B by 2100 | Australia | Inland | Freshwater shrimp | Increased production in non-native zones | |
Temperature increase, ocean acidification, primary productivity declines | CMIP5 RCP8.5 in 20-year increments to 2090 | Global | Marine | Finfish species | Increased suitable habitat expansion for regions (Russia, Norway, USA Alaska, Denmark, Canada). By 2100, reduction in productivity for major producers (Norway, China) | Froehlich et al. (2018a), |
Temperature increase | 2–5°C increase under RCP8.5 | Europe | Marine | Atlantic salmon | Increased growth | |
Temperature increase | RCP4.5 to 2029 | Norway | Marine | Atlantic salmon | Growth threshold reached by 2029 | |
Temperature increase | Downscaled CM2.6 by 2050 | Global | Marine | Atlantic salmon, cobia and sea bream | Increased or decreased growth rates depending on region | Klinger et al. (2017) |
Temperature increase, ocean acidification, primary productivity declines | CMIP5 RCP8.5 in 20-year increments to 2090 | Global | Marine | Shellfish | Overall declines in suitable habitat globally, up to 50–100% reductions in regions in China, Thailand and Canada | Froehlich et al. (2018a) |
Temperature increase | CMIP5 RCP8.5 by 2050, 2100 | Italy | Marine | Clams | Negative impacts for juvenile timing, spatial distribution, and quality | |
Temperature increase | CMIP5 RCP2.6 and RCP8.5 by 2035, 2070 | France | Marine | Oysters | Increase incidence of oyster mortality; increase by 2035 to annual occurrence by 2070 | |
Temperature increase | RCP2.6 and RCP8.5 by 2050 | Global | Marine | Shellfish | Species reduction (10–40%) in tropical and subtropical regions, with increase (40%) in higher latitudes | |
Temperature increase, ocean acidification | Ecopath with RCP8.5 by 2100 (2.8°C warming and pH 7.89) | USA | Marine | Shellfish | Reduction primary productivity and subsequent bivalve carrying capacity | |
Temperature increase, stratification change | RCP8.5 by 2088–2099 | Spain | Marine | Mussels | Decline in mussel optimal culture conditions of 60% in upper and 30% in deeper waters by 2099 | |
Temperature increase, ocean acidification | RCP2.6 and 8.5 by 2070–2090 | Global | Marine | Shellfish | Under RCP8.5, a decline in shellfish production due to primary productivity reduction in tropical regions and gains in high latitudes. Under RCP2.6, marine production will have net gain | |
Temperature increase | 4°C increase | Global | Marine | Vibrio spp. (mortality causative agent) | Increased virulence | |
Temperature increase (marine heatwave) | 5°C increase | Global | Marine | Oysters | Increased oyster mortality | |
Ocean acidification | ~2000 ppm CO2 | Global | Marine | Oysters | Impaired immune function | |
Ocean acidification | RCP8.5 in 20-year increments to after 2099 | USA | Marine | Shellfish | Regional projected vulnerabilities; southern Alaska and Pacific Northwest at more immediate risk | |
Ocean acidification | A1B and RCP8.5 by 2100 | UK | Marine | Shellfish | Regional projected vulnerabilities; Wales and England at more immediate risk | |
Ocean acidification | RCP2.6 and RCP8.5 by 2300 | East China | Marine | Shellfish | Carbonate saturation projected to decrease by 13% and 72% under RCP2.6 and RCP8.5 respectively, projecting decreased shellfish productivity | RCP2.6 and RCP8.5 by 2300 (Zhang et al., 2017b) |
Increased temperature | RCP2.6 and RCP8.5 by 2100 | North Sea | Marine | Seaweed | Northward population shift by 110–163 km and 450–635 km under RCP2.6 and RCP8.5, respectively | |
Increased temperature | RCP4.5 and RCP8.5 by 2090 | Japan | Marine | Kelp | Habitat decline to 30–51% and 0–25% under RCP4.5 and RCP8.5, respectively |
5.9.3.2.4 Societal impacts within the production system
Marine aquaculture provides distinct ecosystem services through provisioning (augmenting wild fishery catches), regulating (coastal protection, carbon sequestration, nutrient removal, improved water clarity), habitat and supporting (artificial habitat) and cultural (livelihoods and tourism) services (Gentry et al., 2020), which vary with species, location and husbandry (Alleway et al., 2019). Projected thermal increases of 1.5°C will reduce ecosystem services, further reduced under 2°C warming, with associated increases in acidification, hypoxia, dead zones, flooding and water restrictions (medium confidence) (Hoegh-Guldberg et al., 2018). Sudden production losses from extreme climate events can exacerbate food security challenges across production sectors, including aquaculture, increasing global hunger (high confidence) (Cottrell et al., 2019; Food Security Information Network, 2020). While aquaculture provides positive influences such as food security and livelihoods, there are negative concerns over environmental impacts (including high nutrient loads from sites) and socioeconomic conflicts (Alleway et al., 2019; Soto et al., 2019), and adoption of ecosystem approaches is dependent on particular user groups and regions (Gentry et al., 2017; Brugère et al., 2019; Gentry et al., 2020). In coastal Bangladesh, projected saline inundation to wetland ecosystem services will result in ecosystem services losses of raw materials and food provisioning, ranging from USD 0 to 20.0 million under RCP2.6 to RCP8.5 scenarios (Mehvar et al., 2019). Mangrove deforestation for shrimp farming in Asia negatively impacts ecosystem services and reduces climate resilience (medium confidence) (Mehvar et al., 2019; Nguyen and Parnell, 2019; Reid et al., 2019; Custódio et al., 2020), while mangrove reforestation efforts may have some effectiveness in re-creating important nursery grounds for aquatic species (low confidence) (Gentry et al., 2017; Chiayarak et al., 2019; Hai et al., 2020). Families are highly vulnerable to climate change where nutritional needs are being met by self-production, such as in Mozambique, Namibia (Villasante et al., 2015), Zambia (Kaminski et al., 2018) and Bangladesh (high confidence) (Pant et al., 2014). Climate change will therefore affect multiple ecosystem services where ultimately decisions on balance or trade-offs will vary with regional perceptions of service value (high confidence).
5.9.4 Aquaculture Adaptation
5.9.4.1 Adaptation planning
Aquaculture is often viewed as an adaptation option for fisheries declines, thereby alleviating food security from losses of other climate change impacts (Sowman and Raemaekers, 2018; Johnson et al., 2020) such as Pacific Islands freshwater aquaculture, Bangladesh crop-aquaculture systems or Viet Nam rice–fish cultivations (Soto et al., 2018). Many adaptations are specific to regions, countries or sectors, implemented on a regional to national scale (FAO, 2018c; Galappaththi et al., 2020b). Adaptation likelihood (potential), effectiveness and risk of maladaptation was assessed per major FAO production region for inland, brackish and marine aquaculture (Figure 5.16) production systems. Potential adaptation measures to reduce production loss can be built upon existing adaptation planning and guidelines, to reduce the risk of maladaptation including feedback loops (e.g., FAO, 2015; Bueno and Soto, 2017; Dabbadie et al., 2018; FAO, 2018c; Poulain et al., 2018; Brugère et al., 2019; Pham et al., 2021; Soto et al., 2021). Large climate change adaptation strategies for the aquaculture sector exist, such as in the USA (Link et al., 2015), Australia (Hobday et al., 2017) and South Africa (Department of Environmental Affairs, 2016). Lower-income countries often lack financial, technical or institutional capacity for adaptation planning (Galappaththi et al., 2020b), but examples include Bangladesh and Myanmar (FAO, 2018c), with programmes offering adaptation funding (Dabbadie et al., 2018). Early participation of stakeholders in adaptive planning has promoted action and ownership of results (high confidence), such as in India and the USA (Link et al., 2015; FAO, 2018c; Soto et al., 2018) Early outreach, education and knowledge gap assessments raise awareness, where utilisation of local knowledge and Indigenous knowledge and scientific involvement support informed adaptive planning and uptake for all stakeholders (high confidence) (Cooley et al., 2016; FAO, 2018c; Rybråten et al., 2018; Soto et al., 2018; McDonald et al., 2019; Galappaththi et al., 2020b), as perceptions of climate risk and capacity will vary (Tiller and Richards, 2018). Supporting the active involvement of women helps address gender inequity and perceived risk, particularly for smallholder farmers (high confidence) (Morgan et al., 2015; Barange and Cochrane, 2018; FAO, 2018c; Avila-Forcada et al., 2020). However, regional and national political influences, financial and technical capacity, governance planning and policy development will ultimately support or hinder adaptation for aquaculture (high confidence) (Cooley et al., 2016; FAO, 2018c; Galappaththi et al., 2020b; Greenhill et al., 2020).
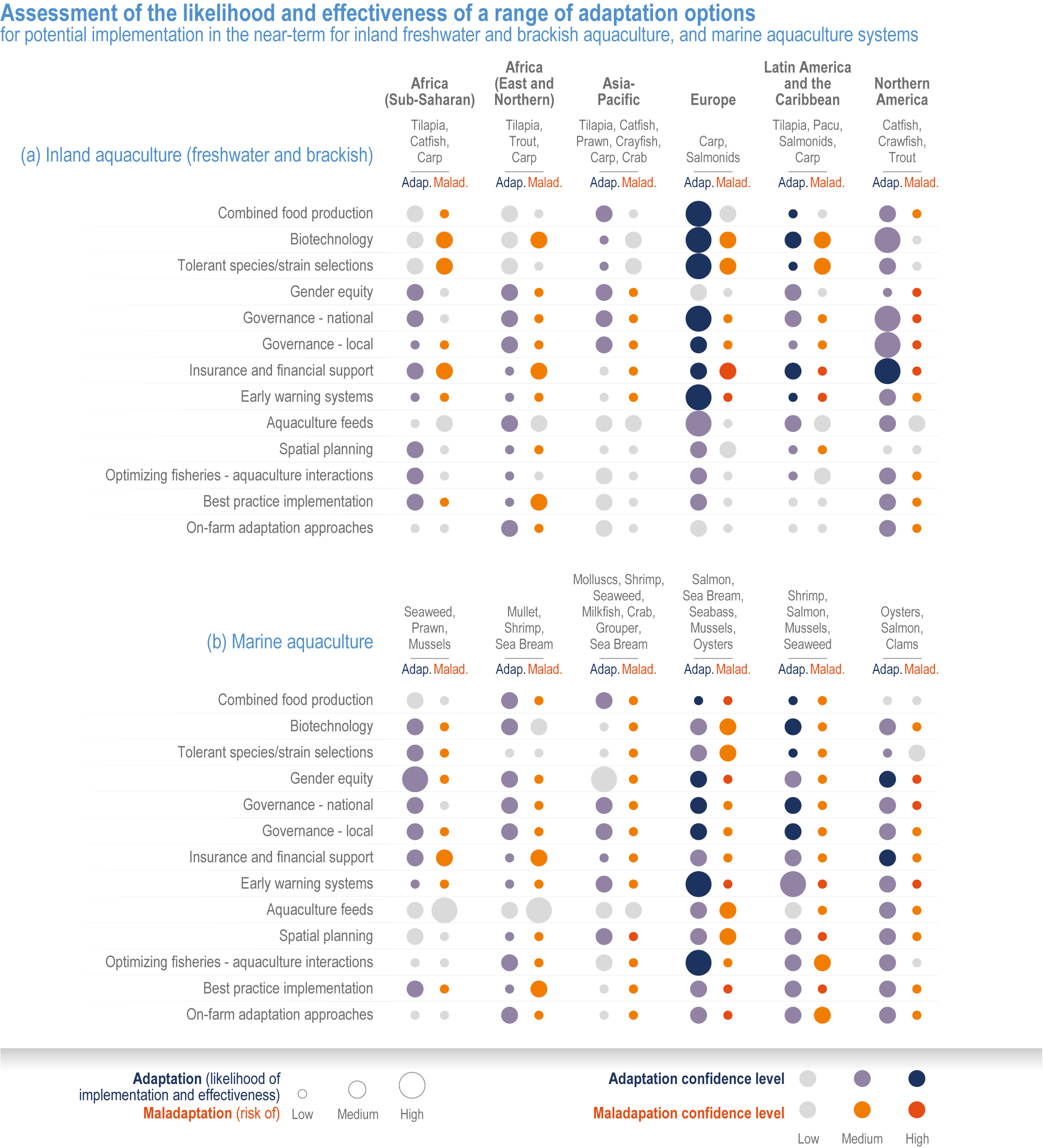
Figure 5.16 | Assessment of the likelihood and effectiveness of a range of adaptation options for potential implementation in the near term (next decade) for inland freshwater and brackish aquaculture (salinities of <10 ppm and/or no connection to the marine environment) (a) and marine aquaculture systems (b) per major FAO production zone. See SM5.6 (Tables SM5.8, 5.12) for assessment methodologies.
5.9.4.2 Species selections and selective breeding
Adaptation options at the operational level include species selections, such as cultivation of brackish species (shrimp, crabs) during dry seasons, and rice-finfish in wetter seasons in Thailand (Chiayarak et al., 2019), use of salt-tolerant plants in Viet Nam (Nhung et al., 2019; Paik et al., 2020), converting inundated rice paddies into aquaculture, rotating shrimp, and rice culture (high confidence) (Chiayarak et al., 2019). Species diversification through co-culture, integrated aquaculture–agriculture (e.g., rice–fish) or integrated multi-trophic culture (e.g., shrimp–tilapia–seaweed or finfish–bivalve–seaweed) may maintain farm long-term performance and viability by: creating new aquaculture opportunities; promoting societal and environmental stability; reducing GHG emissions through reduced feed usage and waste; and carbon sequestration (medium confidence) (see Section 5.10, Ahmed et al., 2017; Bunting et al., 2017; Gasco et al., 2018, Soto et al., 2018; Ahmed et al., 2019; Dubois et al., 2019; FAO, 2019c; Li et al., 2019; Freed et al., 2020; Galappaththi et al., 2020b; Prasko et al., 2020; Tran et al., 2020). In practice, most aquaculture operations concentrate on single-species systems (Metian et al., 2020), and barriers such as land availability, freshwater resources and lack of credit access may limit the uptake and success of integrated adaptation approaches to climate change (Ahmed et al., 2019; Tran et al., 2020; Kais and Islam, 2021).
Selective breeding can promote climate resilience (medium confidence) (Klinger et al., 2017; Fitzer et al., 2019), and operations have already intentionally, or unintentionally, selected for production traits for changing conditions (de Melo et al., 2016; Tan and Zheng, 2020). Exposure of broodstock to future climate conditions may or may not confer advantages to offspring (moderate evidence, low agreement ) (Parker et al., 2015; Griffith and Gobler, 2017; Thomsen et al., 2017; Durland et al., 2019). Traditional pedigree developments require extensive phenotypic data, but genomic selections can rapidly select for robust climate-associated traits (Sae-Lim et al., 2017; Gutierrez et al., 2018; Zenger et al., 2018; Houston et al., 2020; Tan and Zheng, 2020). Genomic resources are available for salmon, rainbow trout, coho, carp, tilapia, seabass, bream, turbot, flounder, catfish, yellow drum, scallops, oysters and shrimp, but have been developed for disease and growth selections rather than climate resistance (Dégremont et al., 2015a; Dégremont et al., 2015b; Abdelrahman et al., 2017; Gjedrem and Rye, 2018; Gutierrez et al., 2018; Guo et al., 2018; Liu et al., 2018a; FAO, 2019d; Houston et al., 2020), although bivalve selections for ocean acidification and warming resiliency are underway (Tan and Zheng, 2020). Targeted genome editing could modify phenotypes of major aquaculture species (Li et al., 2014 a; Elaswad et al., 2018; Yu et al., 2019; Houston et al., 2020), but uptake is dependent upon national regulatory and public approvals. Local adaptations within species with higher climate resiliencies may assist in selections (Thomsen et al., 2017; Falkenberg et al., 2019; Scanes et al., 2020; Toomey et al., 2020), but highlight the need to consider specific farming environments for selective processes (Houston et al., 2020). Projections of climate on aquaculture production traits are not well understood (Lhorente et al., 2019); therefore, genetic diversity needs to be maintained to ensure population fitness (high confidence) (Bitter et al., 2019; Lhorente et al., 2019; Visch et al., 2019; Houston et al., 2020; Mantri et al., 2020).
5.9.4.3 Farm site selection, infrastructure and husbandry
Land-based aquaculture systems including hatcheries may reduce exposure to climatic extremes (due to better control of the culture environment), limit water usage, reduce juvenile reliance and buffer climate effects using optimal diets (high confidence) (Barton et al., 2015; Reid et al., 2019; Cominassi et al., 2020). However, land-based aquaculture requires large capital and operational costs and use of land, increasing conflicts between land and water use, have increased energy demands (increasing GHG if fossil fuels are the primary energy source), require necessary expertise and will not reduce outgrowing exposures (high confidence) (see Section 5.13, Beveridge et al., 2018b; Soto et al., 2018; Tillotson et al., 2019; Costello et al., 2020; Prakoso et al., 2020).
Geographical selection of marine farm sites may prevent climate productivity declines (medium confidence) (Froehlich et al., 2018a; Sainz et al., 2019; Oyinlola et al., 2020), particularly for temperature-related mortality hotspots (Garrabou et al., 2019), HAB occurrences (Dabbadie et al., 2018) or extreme events (Liu et al., 2020; Wu et al., 2020). However, while downscaled climate forecasts facilitate localised adaptation planning (Falconer et al., 2020a), such projections are rare (Whitney et al., 2020). GIS can be used for climate adaptive planning along with routine site assessments (Falconer et al., 2020b; Galappaththi et al., 2020b; Jayanthi et al., 2020). Building coastal protection, stronger cages and mooring systems, and deeper ponds and using sheltered bays can reduce escapees and mortalities related to flooding, increased storms and extreme events (medium confidence) (Dabbadie et al., 2018; Bricknell et al., 2021; Kais and Islam, 2021). Inshore aquaculture in low-lying areas prone to sea level salinity intrusion (e.g., Mekong delta and Viet Nam) have already implemented adaptation measures, such as conversion of land to mixed plant–animal systems (Nguyen et al., 2019a), conversion of freshwater ponds to brackish or saline aquaculture (Galappaththi et al., 2020b), building of dams and dykes (Renaud et al., 2015) and intensification of shrimp or fish pond culture to reduce water and land usage (Nguyen et al., 2019b; Johnson et al., 2020). Other adaptation options for limited water supply are government equitable water allocations and water storage (high confidence) (Bunting et al., 2017; Galappaththi et al., 2020b).
Feed formulations and improved feed conversion can reduce climate-associated stress for freshwater species, significantly reducing waste and increase sustainability (medium confidence) (FAO, 2018c; Gasco et al., 2018; Chen and Villoria, 2019). Projected decreases in fish meal and global targets of limiting warming to under 2°C may increase the ratio of plant-based diets but reduce fish nutritional content (see Sections 5.10 and 5.13, Hasan and Soto, 2017; Johnson et al., 2020). Companies provide insurance in major production areas, but aquaculture is considered high risk with large levels of small claims (Secretan et al., 2007). Insurance covers natural disasters and disease, helping to reduce and cope with climate-induced risk, enabling faster livelihood recoveries and preventing poverty (high agreement , limited evidence) (Xinhua et al., 2017; Kalikoski et al., 2018; Soto et al., 2018). For example, small-scale shrimp farmers were willing to pay higher premiums to manage risk, after participation in government pilot insurance schemes, ensuring greater pay-outs if a mortality event occurred (Nyguyen and Pongthanapanic, 2016; Pongthanapanic et al., 2019). Technological innovations are more widely implemented in larger operations, with Internet access promoting adoption at the farm site (Joffre et al., 2017; Salazar et al., 2018). Improved farm management is a key opportunity (high confidence) to reduce climate risks on aquaculture, where Best Management Practices can increase resiliency (Soto et al., 2018) and lower additional risk from non-climatic stressors (Gattuso et al., 2018; Smith and Bernard, 2020), and decision-tree frameworks can provide adaptation choices when events occur (Nguyen et al., 2016).
5.9.4.4 Early-warning and monitoring systems
Globally, monitoring is increasing to fill scientific uncertainties (Goldsmith et al., 2019) but is not often at spatial scales which facilitate farm or regional adaptation management (Whitney et al., 2020) or data complexities prevent direct uptake by operators, resource managers and policymakers (medium confidence) (Soto et al., 2018; Gallo et al., 2019). Specialised industry portals (Pacific shellfish) and government-established monitoring programmes (Chilean salmon) and other observational networks (e.g., Global Ocean Acidification Observing Network (GOA-ON)) can provide real-time monitoring and early-warning event alerts and facilitate aquaculture decision making (medium confidence) ( Cross et al., 2019; Farcy et al., 2019; Soto et al., 2019; Tilbrook et al., 2019; Bresnahan et al., 2020; Peck et al., 2020). Seasonal forecasting, downscaled models and early-warning systems provide valuable regional or farm site risk information (Hobday et al., 2018; Galappaththi et al., 2020b; Whitney et al., 2020), but monitoring will need to be useful for farmers, involve farmers, and be accurate, timely, cost-effective, reviewed and maintained in order to ensure uptake (high confidence) (Soto et al., 2018). Early-warning systems for HABs enable rapid decision making and risk mitigation (medium confidence), such as ocean colour monitoring in South Africa (Smith and Bernard, 2020), where early harvesting and additional husbandry were used to minimise production and economic losses (Pitcher et al., 2019). New tools, strategies and observations are needed to predict HAB occurrences and range shifts with changing climate (high confidence) (Schaefer et al., 2019; Tester et al., 2020), as there is uncertainty on drivers of incidence and toxicity (Wells et al., 2020).
5.9.5 Contributions of Indigenous, Traditional and Local Knowledge
Indigenous mariculture practices, such as intertidal clam gardens, have been occurring for thousands of years, providing knowledge of traditional practices still applicable to mariculture (Deur et al., 2015; Jackley et al., 2016; Poulain et al., 2018; Bell et al., 2019; Toniello et al., 2019). Indigenous groups differ in opinions on aquaculture acceptability, implications for coastal management and territorial rights (high confidence) (Young et al., 2019). Such perceptions may determine culturally appropriate types and benefits of aquaculture (employment, food diversification, income, building autonomy and skillsets), such as in Australia (Petheram et al., 2013) and Canada (Young and Liston, 2010). Marginalised people, like small-scale aquaculture farmers in lower-income and lower-middle-income countries, are often overlooked and are not represented at a governance level (Barange et al., 2014; Kalikoski et al., 2018). Therefore policy, economic, knowledge and other support must ensure representation with traditional and other stakeholder ecological knowledge at national, regional and local levels to facilitate climate change adaptation and safeguard human rights for poor and vulnerable groups (high confidence) (Kalikoski et al., 2018; Poulain et al., 2018).
5.10 Mixed Systems
The food and livelihoods of many rural people depend on combinations of crops, livestock, forestry and fisheries, and still information on these mixed systems is scarce. Rural households in low- and middle-income countries earn almost 70% of their income through mixed production systems (Angelsen et al., 2014). These systems produce about half of the world’s cereals, most of the fruits, vegetables, pulses, roots and tubers, and most of the staple crops and livestock products consumed by poor people in lower-income countries (Herrero et al., 2017). They can help in adapting to climatic risks and reducing GHG emissions by improving nutrient flows and improving the recycling of nutrients within the production system and by increasing food production and diet quality per unit of land and diversifying income sources (Smith et al., 2019c). Indigenous groups often practice mixed production, integrating crops, animals, fisheries, forestry and agroforestry through traditional ecological knowledge.
Some evidence exists of the buffering capacity that integrated systems can provide in the face of climate change (Gil et al., 2017). This buffering, often affecting the farming system as a whole rather than the individual agricultural enterprises involved, applies to some aquaculture–agriculture systems as well as to crop–livestock systems (Bunting et al., 2017; Stewart-Koster et al., 2017). In some situations, there may be trade-offs and constraints at the household level that affect this resilience-conferring ability: for instance, mixed systems often need relatively high levels of management skill, and extra labour may be required (van Keulen and Schiere, 2004; Thornton and Herrero, 2015). The diversification of food production systems offers promise for enhanced resilience at the global level (Kremen and Merenlender, 2018; Dainese et al., 2019; Section 5.4.4.4), though policies need to provide adequate incentives for resource efficiency, equity and environmental protection (Havet et al., 2014; Thornton and Herrero, 2014; Troell et al., 2014).
5.10.1 Observed Impacts
5.10.1.1 Mixed crop–livestock systems
Overall, there is high confidence that farm strategies that integrate mixed crop–livestock systems can improve farm productivity and have positive sustainability outcomes (Havet et al., 2014; Thornton and Herrero, 2014; Herrero et al., 2015; Thornton and Herrero, 2015; HLPE, 2019). The scale of the improvement varies between regions and systems and is moderated by overall demand in specific food products and the policy context. Integrated crop–livestock systems present opportunities for the control of weeds, pests and diseases. They can also provide a range of environmental benefits, such as increased soil carbon and soil water retention, increased biodiversity and reduced need for inorganic fertilizers (Havet et al., 2014; Thornton and Herrero, 2014; Herrero et al., 2015; Thornton and Herrero, 2015; HLPE, 2019).
Research indicates that mixed crop–livestock systems are often more resilient to climate change (medium confidence). In the southern Afar region of Ethiopia, crop–livestock households were more resilient than livestock-only households to climate-induced shock (Mekuyie et al., 2018). However, the benefits of managing crop–livestock interactions in response to climate change depend on local context. For example, in higher-rainfall zones in Australia, Nie et al. (2016) found some yield reductions and difficulty in maintaining groundcover. The systematic review of Gil et al. (2017) concluded that the integration of crop and livestock enterprises as an adaptation measure can enhance resilience (FAQ 5.1).
Reconfiguration of mixed farming systems is occurring. In semi-arid eastern Senegal, Brottem and Brooks (2018) found increasing reliance on livestock production mostly because of changing climate conditions. Many poorer households are having to rely on migration to compensate for shortfalls in crop production arising from a changing climate. Some farmers have successfully shifted to crop–livestock systems in Australia, where they have allocated land and forage resources in response to climate and price trends (Bell et al., 2014).
Mixed livestock–crop systems may increase burdens on women, require managing competing uses of crop residues, and have higher requirements of capital and management skills. These factors can be challenging in many lower-income countries (Rufino et al., 2013; Thornton and Herrero, 2015; Jost et al., 2016; Thornton, 2018). The policy actions needed for the successful operation of mixed crop–livestock systems may be similar across widely different situations: good access to credit inputs and capacity building needed to facilitate uptake (Hassen et al., 2017; Marcos-Martinez et al., 2017), and good levels of market infrastructure (Ouédraogo et al., 2017; Iiyama et al., 2018).
5.10.1.2 Mixed crop–aquatic systems
Households may have a mix of aquatic and land-based food production, contributing to food security and nutrition and income generation (Freed et al., 2020; see also discussion of aquaponics and hydroponics in Section 5.10.4.3. and combined rice–aquatic species production in Section 5.9.4). Failures in agricultural outputs due to climate-associated factors may result in diversification to fisheries as a way of alleviating food production shortfalls; for example, fisheries landings may dramatically increase after agricultural failures following hurricanes, which can subsequently create overfishing collapses (Cottrell et al., 2019). Where climatic impact drivers affect multiple sectors, adaptation may become more difficult because of the interacting challenges (Cottrell et al., 2019). One study of 12 countries with high food insecurity levels found that fish-reliant households utilised as much land as those not reliant on fish (Fisher et al., 2017). To meet food security requirements, most of these households needed to both farm and fish, illustrating the interdependence of aquatic–terrestrial food systems.
5.10.1.3 Agroforestry systems
Agroforestry is frequently mentioned as a strategy to adapt to and mitigate climate change and address food security (de Coninck et al., 2018; Smith et al., 2019c). There is strong evidence of net positive biophysical and socioeconomic effects of agroforestry systems under both smallholder and large-scale mechanised production systems (Quandt et al., 2017; Hoegh-Guldberg et al., 2018; Sida et al., 2018; Wood and Baudron, 2018; Table 5.10; Cross-Chapter Box NATURAL in Chapter 2; Quandt et al., 2019). Many of these effects also reduce climate risk. At the same time, agroforestry systems are subject to impacts from climate change, potentially reducing the benefits they provide. Still, there is limited evidence of observed climate impacts on agroforestry systems, and modelling climate impacts is more complex for agroforestry than for single cropping systems (Luedeling et al., 2014).
5.10.2 Assessing Vulnerabilities
5.10.2.1 Assessing vulnerability in mixed systems
Important information gaps exist concerning the costs and benefits of many adaptation options in mixed systems, where the interactions between farming enterprises may be complex. Among communal crop–livestock farmers in Eastern Cape Province of South Africa, Bahta (2016) reported high levels of vulnerability to drought and highlighted the need for more coordination between monitoring agencies in terms of reliable early-warning information that can be communicated appropriately, between farmers’ organisations and the private sector to facilitate adaptation options that can overcome feed shortages such as fodder purchases in times of drought, and between government departments at the national and provincial level that address the concerns and needs of affected communities. Nyamushamba (2017) reviewed the use of indigenous beef cattle breeds in smallholder mixed production systems in southern Africa. Some of these breeds exhibit adaptive traits such as drought and heat tolerance and resistance to tick-borne diseases. However, their adaptation potential in crossbreeding programmes is essentially unknown, as most African cattle populations are still largely uncharacterised.
5.10.2.2 Social vulnerabilities
As in other production systems, Indigenous groups, gender, race and other social categories can result in heightened vulnerability to climate change in mixed production systems owing to historical and current marginalisation and discrimination (high confidence) (Parraguez-Vergara et al., 2016; Baptiste and Devonish, 2019; Moulton and Machado, 2019; Popke and Rhiney, 2019; Fagundes et al., 2020). A study of the Mapuche Indigenous group in Chile found that marginalisation and discrimination worsened their vulnerability and observed impacts of climate change because they had less access to services and lower incomes and were not as high a priority as other groups (Parraguez-Vergara et al., 2016). Among fisherfolk on Lake Wamala, Uganda, Musinguzi (2018) found evidence of considerable diversification to crop and livestock production as a means of increasing households’ food security and income, but women had greater workloads and less control over new income sources than men. Ngigi (2017) evaluated adaptation actions within households in rural Kenya and found that women tended to adopt adaptation strategies related to crops, and men to livestock and agroforestry activities. Chingala (2017) found substantial gender- and age-related differences in control of access to animal feed, animal health and water resources in beef producers in mixed crop–livestock systems in Malawi. In a review of agriculture–aquaculture systems in coastal Bangladesh, Hossain et al. (2018) showed that existing policies and adaptation mechanisms are not adequately addressing gender power imbalances, and women continue to be marginalised, leading to increasing feminisation of food insecurity. Such studies highlight the need to consider gender and other social inequities when examining adaptation in mixed production systems, particularly in situations in which men and women have different levels of control over productive assets (Cross-Chapter Box GENDER in Chapter 18).
5.10.3 Projected Impacts
The impacts of climate change on risk in mixed farming systems are projected to be dependent on market, ecosystem and policy context (medium evidence, low agreement ). In mixed crop–livestock farms in a semi-arid region of Zimbabwe, Descheemaeker (2018) found that feeding forages and grain could alleviate dry-season feed gaps to the 2050s, but their effectiveness depended on the household’s livestock stocking density. In comparing different commercial production systems, Tibesigwa (2017) found that, under South African conditions, climate change to the 2050s will reduce productivity across the agricultural sector, with the largest impacts occurring in specialised commercial crop farms owing to their relative lack of diversity. Mixed farming systems were the least vulnerable in terms of relative effects on farm output; this applied to commercial and subsistence sectors (Tibesigwa et al., 2017). Other studies suggest increased risk in mixed systems in semi-arid conditions. In northern Burkina Faso, Rigolot (2017) examined different crop fertilisation and animal supplementation levels under RCP8.5 to the 2050s. They found that, although aggregate profits could be increased via moderate levels of inputs, the use of external inputs may increase risk because of marginal costs exceeding marginal benefits in lower rainfall years. In the Western Australian wheat belt, Thamo (2017) assessed climate-change-induced shifts in farm profitability to the 2050s. For most options, the adverse effects on profitability were greater than the advantageous effects, profit margins being much more sensitive to climate change than production levels. However, in the same system, Ghahramani (2018) evaluated adaptation options to 2030 and found that a shift to a greater reliance on livestock could be profitable, even in years with low rainfall.
Risk management in integrated production systems may constitute a barrier to uptake of adaptation options (Rigolot et al., 2017). Watson (2018) highlighted the current lack of financial risk management tools that could be used in smallholder coastal communities. Alongside other risk management tools such as weather-based index insurance, risk pooling may find wide application in different farming systems as an effective adaptation measure (medium agreement , limited evidence) (Hansen et al., 2019a).
Climate change impacts on productivity of agroforestry systems are similar to individual perennial crops, although there is limited research on tree crops (see Section 5.4.1.2). Impacts include increased temperature or water stress, an increase in pathogens affecting crops, changes to pollinator abundance, and changes in the nutrient content of one or more of the agroforestry components. Many tree products such as fruits and nuts are grown in agroforestry settings. The quality and nutrition of these products and other specialty crops are often negatively affected by rising temperatures, ambient CO2 concentrations and tropospheric ozone (Ahmed and Stepp, 2016). There is also evidence that the fungus coffee rust will be positively affected by climate change (Avelino et al., 2015; Bebber et al., 2016), with adverse effects on coffee agroforestry systems.
While shade trees can ameliorate increasing stand temperatures that will significantly impact arabica coffee (Ovalle-Rivera et al., 2015; Schroth et al., 2015), the opposite can also be true. Comparing shade and full-sun coffee systems in Ghana, Abdulai (2018) concluded that the leguminous tree species providing shade and additional nitrogen led to soil water competition with the coffee trees during severe drought, resulting in enhanced coffee mortality. On the other hand, experimentally induced drought in a soybean-intercropping agroforestry system in eastern Canada led to crop losses in the monocropping system only, whereas N-fixation declined in both systems (Nasielski et al., 2015). Thus, balancing the synergies and trade-offs of multiple component systems is necessary based on local context. While species diversification can enhance resilience to climate shocks, lack of water can constrain the implementation of agroforestry practices in arid locations (Apuri et al., 2018).
For people reliant on both agriculture and fisheries for food production, regional differences in productivity effects of climate change are expected; populations in LMICs that are already vulnerable will be most affected by simultaneous reductions in fisheries and agricultural productivity (Blanchard et al., 2017). Twelve out of 17 high-income countries in Europe showed projected increases in agricultural production where adaptive capacity is higher, and agricultural and food fisheries’ dependence was lower. Some LMIC countries (Nigeria, Cameroon, Ghana and Gabon) showed relative reductions in both fisheries and agricultural production, where food insecurity, human population growth and fisheries overexploitation rates are high (Blanchard et al., 2017). Model projections under the RCP6.0 scenario show decrease in marine and terrestrial production to 2050 in 87 out of the 119 coastal countries studied, even though there is a wide variance in adaptive capacity and relative and combined dependencies on fisheries and agriculture (Blanchard et al., 2017). A projected 2050 move towards greater consumption of cultured seafood and less meat showed that aquaculture requires less feed crops and land, but was regionally dependent upon differing patterns of production, trade and feed composition (Froehlich et al., 2018b).
Box 5.7: Perspectives of Crop and Livestock Farmers on Observed Changes in Climate in the Sahel
The Sahel region of West Africa has experienced some of the most severe multi-decadal rainfall variations in the world: excessive rainfall in the 1950s–1960s followed by two decades of deficient rainfall, leading to a large negative trend until the mid-to-late 1980s with a decrease in annual rainfall of between 20% and 30%. Recently, there has been a partial recovery of annual rainfall amounts, more significant over the central than the western Sahel. This recovery is characterised by new rainfall features, including false starts and early cessation of rainy seasons, increased frequency of rainy days, increased precipitation intensity and more frequent and longer dry spells (Salack et al., 2015; Sanogo et al., 2015; Salack et al., 2016; Biasutti, 2019). The Sahel is experiencing a new era of rainfall extremes (Bichet and Diedhiou, 2018; Panthou et al., 2018), suggesting an intensification of the hydrological cycle (Doblas-Reyes et al., 2021).
The ways in which crop and livestock farmers in the Sahel have responded to climatic variability have been studied widely (Sissoko, 2011; Gonzalez et al., 2012; Jalloh et al., 2013; Gautier et al., 2016; Sultan and Gaetani, 2016; Zougmoré et al., 2016; Segnon, 2019). Local communities have developed an extensive Indigenous ecological knowledge system, enabling them to make use of ecosystem services to support their livelihoods and to survive environmental change (Nyong et al., 2007; Mertz et al., 2009; Lahmar et al., 2012; Segnon et al., 2015). These knowledge systems have been crucial in people’s resilience to and recovery from major environmental change, such as the severe drought period experienced in the region in the 1970s and 1980s (Nyong et al., 2007; Lahmar et al., 2012; Segnon et al., 2015; Gautier et al., 2016; Zouré et al., 2019). As climate change became evident and a primary concern on the global agenda, interest in local people’s knowledge and understanding of climate change has also increased (Mertz et al., 2009; Tambo and Abdoulaye, 2013; Traore et al., 2015; Kosmowski et al., 2016; Sanogo et al., 2017; Segnon, 2019).
There is no simple understanding of crop and livestock farmers’ response in the Sahel to rainfall variability. Nielsen and Reenberg (2010) developed human–environment timelines for the period 1950–2008 for a small village in northern Burkina Faso, relating livelihood diversification and crop–livestock management changes that map closely to local rainfall variability, such as fields abandoned in dry years and intense animal manure use in wet years. Although they found a significant correlation between crop–livestock management practice changes and major climatic events, the climate is only one of many interacting factors that influence local adaptation strategies (Mortimore, 2010; Nielsen and Reenberg, 2010; Sendzimir et al., 2011). Robust attribution of observed changes to specific change drivers remains a challenge.
Crop and livestock farmers’ knowledge and perceptions of increases in temperature and temperature-related stressors (heatwaves, number of extreme hot or cold days) are consistent with the observed meteorological data (Mertz et al., 2009; Mertz, 2012; Tambo and Abdoulaye, 2013; Traore et al., 2015; Sanogo et al., 2017; Segnon, 2019). Their perceptions of changes in rainfall amounts have not always been consistent with the observational record (Mertz, 2012; Segnon, 2019). Nevertheless, their perception of increases in dry spell occurrence during the rainy season and changes in rainfall pattern (onset, cessation, intensity and distribution) were consistent with the recent observations (Barbier et al., 2009; Ouédraogo et al., 2010; Tambo and Abdoulaye, 2013; Salack et al., 2015; Traore et al., 2015; Kosmowski et al., 2016; Salack et al., 2016; Segnon, 2019). Rainfall patterns within the season, rather than the total amounts of rainfall, matter more for crop and livestock farmers in the Sahel (Segnon, 2019).
Crop and livestock farmers in the Sahel have a sophisticated understanding of the local climate. There is considerable potential to harness this knowledge, coupled with an enabling institutional environment, in developing policies and adaptation plans (Rasmussen et al., 2018); the Sahel is a region where meteorological stations and observed data are scarce (Buytaert et al., 2012; Nkiaka et al., 2017). A deeper understanding of the resilience of local ecological knowledge systems, in light of the hydro-climatic intensification currently experienced in the region and future changes, may well provide further insights into their long-term effectiveness.
5.10.4 Adaptation Strategies
5.10.4.1 Increasing integration and diversity within mixed systems
There is medium confidence in the effectiveness of changing the nature of the integration between crops and livestock as an adaptation: moving from crops to livestock, moving from livestock to crops, and moving from one species of livestock to others, for example (Roy et al., 2018). Such transitions that increase integration between farm enterprises may contribute to risk reduction and increased food security. In areas with adequate rainfall and relatively limited rainfall variability under climate change, where agricultural diversity is the greatest, transitions towards more diverse and integrated systems may bring substantial adaptation benefits (Waha et al., 2018).
Barriers to increasing integration and diversification include policies which support cereals and crop specialisation, lack of markets, limited post-harvest processing, limited technical or biophysical research on implementation and poor market infrastructure (Keatinge et al., 2015; Bodin et al., 2016; Garibaldi et al., 2016; Bassett and Koné, 2017; Kongsager, 2017; Rhiney et al., 2018; Roesch-McNally et al., 2018; Clay and King, 2019; Ickowitz et al., 2019). Proactive policy and market development are needed to reduce these barriers (Clay and King, 2019; Ickowitz et al., 2019; See 5.14.3.8 for Insurance).
5.10.4.2 Agroforestry as an adaptation–mitigation strategy for mixed systems
Agroforestry, the purposeful integration of trees or shrubs with crop or livestock systems, increases resilience against climate risks through a range of biophysical and economic effects (high confidence). Traditional agroforestry has been practiced for millennia and provides prime examples of sustainable agroecological production systems meeting the production, income and socio-cultural needs of farming communities within their ecological niches, but market forces have often led to their demise (McNeely and Schroth, 2006; Plieninger and Schaar, 2008; García-Martínez et al., 2016; Krčmářová and Jeleček, 2016; Coq-Huelva et al., 2017; Paudel et al., 2017; Doddabasawa et al., 2018; Maezumi et al., 2018; Lincoln, 2020). The wide range of options to associate different trees with crops, livestock and aquaculture allows agroforestry to be practiced in most regions, including those with precipitation regimes ranging from semi-arid to humid. While most agroforestry systems occur in smallholder settings, there are examples of successful industrial-scale mechanised agroforestry systems (Feliciano et al., 2018; Lovell et al., 2018). Agroforestry delivers medium to large benefits to all five land challenges described in the SRCCL—climate change mitigation, adaptation, desertification, land degradation and food security—and is considered to have broad adaptation and moderate mitigation potential compared with other land challenges (Smith et al., 2019c). Agroforestry is also able to deliver multiple biophysical and socioeconomic benefits (Table 5.12).
Table 5.12 | Some of the biophysical and socioeconomic benefits of agroforestry.
Contribution | Pathway | References |
Increased food security and household income | Diversification of production, avoiding trade-offs between crop and tree products | Nath et al. (2016), Coulibaly et al. (2017), Montagnini and Metzel (2017), Waldron et al. (2017), Blaser et al. (2018), Sida et al. (2018), Quandt et al. (2019), Amadou et al. (2020) |
Increased productivity per unit of land | Introduction of multiple species leading to higher land equivalency ratios | |
Improved biophysical site properties | Via limiting soil erosion, facilitating water infiltration, increasing nutrient use efficiency, improving soil physical properties, improving crop nutritional quality, modifying the site micro-climate, and helping to buffer against extreme events | Nguyen et al. (2013); Carsan et al. (2014), Rosenstock et al. (2014), Quandt et al. (2017), Hoegh-Guldberg et al. (2018), Sida et al. (2018), Wood and Baudron (2018), de Leeuw et al. (2020), Muchane et al. (2020), Nyberg et al. (2020) |
Enhanced biodiversity and supporting ecosystem services | Via integrating different perennial and annual species in different spatial or temporal associations, thereby providing greater habitat diversity for other species, including pollinators and predators | McNeely and Schroth (2006), Imbach et al. (2017), Isbell et al. (2017), Sonwa et al. (2017b), Tran and Brown (2019) |
Enhanced CES | Enhanced recreational, cultural and spiritual uses | |
Carbon dioxide removal | Via enhanced above-ground carbon sequestration compared with most cropping or livestock systems, ranging from 2.6 to 10 Mg C ha −1 yr −1 depending on regional and climatic conditions (>0.7 Gt CO2 e yr −1 globally between 2000 and 2010) | Ramachandran Nair et al. (2009), Zomer et al. (2016), Rochedo et al. (2018), Wolz et al. (2018), Crous-Duran et al. (2019), Platis et al. (2019) |
Enhanced gender balance | Via providing women with more diversified income sources | Kiptot et al. (2014), Ngigi et al. (2017), Benjamin et al. (2018) |
Strengthened urban and peri-urban agricultural systems | Via provision of regulating and provisioning ecosystem services such as shade, water infiltration, new food and livelihood opportunities | See Section 5.12 |
The adoption and maintenance of agroforestry practices require appropriate incentives or the removal of barriers (high confidence). Agroforestry adoption has been limited to date in both higher-income and lower-income countries. Several constraints need to be carefully addressed for successful scaling-up of agroforestry systems, including costs of establishment, limited short-term benefits, lack of reliable financial support to incentivise longer-term returns on investments, land tenure, knowledge of and experience with trees and the management of multiple component systems, and inadequate market access, (Coulibaly et al., 2017; Iiyama et al., 2017; Jacobi et al., 2017; Kongsager, 2017; Hernández-Morcillo et al., 2018; Iiyama et al., 2018; Lincoln, 2019). Kongsager (2017) and Roupsard et al. (2020) also highlight the need for vertical integration of measures from local to national scales to successfully address local barriers to adoption. Although there are few studies evaluating the long-term performance of agroforestry systems (Coe et al., 2014; Meijer et al., 2015; Brockington et al., 2016; Kongsager, 2017; Toth et al., 2017), the available results suggest that successful adoption of agroforestry practices depends strongly on the local enabling environment, including appropriate markets, technologies and delivery systems (medium evidence, high agreement ).
5.10.4.3 Links between crops and aquaponics–hydroponics as adaptation
Hydroponic systems produce plants in a soilless environment requiring mineral fertilizers to meet plant nutritional needs, whereas aquaponics combines an aquaculture production system with hydroponics, where fish waste provides nitrogen, phosphorous and potassium for plant growth and nitrifying and mineralising bacteria act as filters (Goddek et al., 2015; Pérez-Urrestarazu et al., 2019; Ghamkhar et al., 2020). The relative environmental impact of hydroponic systems is lower compared with conventional systems owing to the significant reductions in land use and fertilizer usage (high confidence) (Goddek et al., 2015; Datta et al., 2018; Pantanella, 2018; Suhl et al., 2018; El-Essawy et al., 2019; Jaeger et al., 2019; Monsees et al., 2019; Mupambwa et al., 2019; Pérez-Urrestarazu et al., 2019; Ghamkhar et al., 2020). While studies indicate that aquaponics and hydroponics have higher yields and a lower environmental footprint than conventional agriculture (medium confidence), aquaculture and heated greenhouse production (Pantanella, 2018; Romeo et al., 2018), aquaponic production may need to be coupled or decoupled or have double-recirculation systems to meet the different requirements of farmed fish and crop species (Pantanella, 2018; Suhl et al., 2018; Mupambwa et al., 2019). Aquaponics and hydroponics are a promising adaptation option for urban agriculture, with benefits including a protected growing environment from climate extremes, reduced GHG emissions related to food transportation, reduced food waste, rainwater harvesting and use of food waste (medium agreement , limited evidence) (Goddek et al., 2015; Al-Kodmany, 2018; Clinton et al., 2018; Weidner and Yang, 2020). Such systems show promise for reducing food production environmental footprints and increasing food security, particularly in arid or water-stressed environments (Doyle et al., 2018; Mupambwa et al., 2019). Barriers to aquaponics and hydroponics adoption include market acceptance of cultured fish species and desirability of plant crops, lack of expertise, legal constraints or high investment costs and financial feasibility (Bosma et al., 2017; Al-Kodmany, 2018; Datta et al., 2018; Pantanella, 2018; El-Essawy et al., 2019; Martin and Molin, 2019; Pérez-Urrestarazu et al., 2019; Specht et al., 2019). There ishigh confidence (high agreement , medium evidence) that a major barrier to hydroponic and aquaponics adoption is the requirement for skilled operators (Goddek et al., 2015; Bosma et al., 2017; Datta et al., 2018; McHunu et al., 2018; Pantanella, 2018), which could be mitigated by decoupling systems and disciplines (Pantanella, 2018). As yet, these systems are not widely implemented and information on their climate change impacts is limited.
5.10.4.4 Transitions in and between mixed systems as adaptation strategy
Transitions in and between the different elements of integrated agricultural systems can be an effective adaptation option (medium confidence). Havlik et al. (2014) projected that, by 2030, market-driven autonomous transitions towards more efficient production systems would increase ruminant meat and milk productivity by up to 20% and decrease emissions by 736 MtCO2 e y −1, most of this arising through avoided emissions from the conversion of 162 Mha of natural land. Weindl et al. (2015) assessed the implications of several climate projections on land use change to 2045 and found that shifts in livestock production towards mixed crop–livestock systems would represent a resource- and cost-efficient adaptation option, reducing global agricultural adaptation costs and abating deforestation by about 76 million ha globally. Both studies suggest that public policy support for transitioning livestock production systems to increase their efficiency could be an important lever for reducing adaptation costs and contributing to emissions reductions. This policy support could include modified regulatory and certification frameworks that incentivise livestock producers to adapt and mitigate (Weindl et al., 2015).
Recent reviews have summarised literature on production system transitions, driven at least partly by a changing climate or changing climate variability, that sometimes involves substantial shifts in enterprises and land configurations. These reviews found several cases of transitions affecting pastoral and mixed systems, with a range of responses including intensification, diversification and sedentarisation as well as the abandonment of agriculture (see Section 5.1 4.3.1, Vermeulen et al., 2018; Thornton et al., 2019). The consequences of these system transitions have been mixed; in some cases, the household-level outcomes have been beneficial, while in others not. Policy environments, defined in terms of multi-level governance structures and institutions, are critical enablers of change. The vulnerability of many crop–livestock keepers to climate change is particularly affected by property and grazing rights (high confidence). Identifying the winners and losers from changes in land ownership and the use of communal lands in the coming decades is a key challenge for the research agenda, particularly as climate change impacts in the marginal lands intensify (Reid et al., 2014).
5.11 The Supply Chain from Post-harvest to Food
The food system is more than just the production of food. It includes domestic and international transportation, storage, processing, market infrastructure and institutions that make up value chains, as well as the food environment in which consumers make food purchasing decisions (HLPE, 2017a). Climate change impacts along the value chain alter availability, access and stability of food security. Nutrition-dense foods tend to be more perishable and are thus more vulnerable to limitations of food storage and transportation infrastructure (Ickowitz et al., 2019). Climate-change-related damage to food in storage (e.g., electricity failures and loss of cold storage) and transportation infrastructure (e.g., extreme weather events damaging roads and other infrastructure) could significantly decrease availability and increase the cost of highly perishable, nutritious foods such as fruits, vegetables, fish, meat and dairy.
This discussion of the post-harvest food system (i.e., after production or catch) focuses on three key elements—food safety, storage, and domestic and international transactions—that could see significant climate change impacts, either directly or indirectly. Higher temperatures and humidity can increase post-harvest loss from pests and diseases, increase occurrence of food-borne diseases and contamination, and raise the cost of refrigeration and other forms of preservation. Extreme weather events can cause disruptions to food transport networks and storage infrastructure. Changes in regional weather can cause production centres to shift locations, potentially requiring changes in storage and processing locations. Prices to producers and consumers will change, although directions and magnitudes are determined by local conditions and policies.
Food loss is the harvest not used by industry or for food. Food waste is the subset of food loss that is potentially recoverable for food use. As a product moves in the post-harvest chain to end users, post-harvest food loss from climate change can occur from improper handling to damage from microorganisms, insects, rodents or birds. Post-harvest losses in quality can be the result of stresses and damage to a plant or animal before harvest, including from climate change (Hodges et al., 2011; Medina et al., 2015a). Food waste caused by climate change may occur at both retail units and homes because fresh ingredients and freshly prepared foods are vulnerable to quality reduction and spoilage from exposure to higher temperatures and humidity. Food waste also contributes to climate change by utilising resources that emit GHGs (Galford et al., 2020).
Box 5.8: Climate Adaptation and Maladaptation in Cocoa and Coffee Production
Coffee and cocoa are important crops in low-latitude regions where agriculture is projected to be heavily impacted by climate change. Both crops are at risk from climate change impacts by 2050 (Baca et al., 2014; Ovalle-Rivera et al., 2015; Chemura et al., 2016; Schroth et al., 2016; Bacon et al., 2017; Schreyer et al., 2018; de Sousa et al., 2019; Lahive et al., 2019; Pham et al., 2019; Cilas and Bastide, 2020). Chocolate and coffee are notable among foods in that their carbon footprint ranges from negative to high, as these industries include both low-input agroforestry systems that have many co-benefits, and high-input monoculture systems where crops are grown without shade, in some cases on sites that have been deforested (Poore and Nemecek, 2019). While the coffee industry in many countries has already transitioned from agroforestry to full-sun production (Jha et al., 2014), the cocoa industry is at a turning point, with many growers deciding whether to move to the potentially more productive ‘full-sun system’, despite a general view that the agroforestry system is more resilient to climate change impacts (Rajab et al., 2016; Schroth et al., 2016; Farrell et al., 2018; Niether et al., 2020).
Shade-grown cocoa and coffee agroforestry systems provide an array of ecosystem services, including regulating pests and diseases, maintaining soil fertility, maintaining biodiversity and carbon sequestration (high confidence) (Jha et al., 2014; Rajab et al., 2016; Cerda et al., 2017; Pham et al., 2019). For example, a comparison of Indonesian cocoa stands found that total carbon stocks above and below ground were five times higher in multi-shade agroforestry stands compared with monoculture stands (57 compared with 11 Mg C ha −1), and total NPP was twice as high (18 compared with 9 Mg C ha −1 yr −1). The extra carbon sequestration was achieved without any notable difference in cocoa yield (Rajab et al., 2016). At higher levels of shade, there can be negative impacts on the yield of the understory crop, but careful management of shade trees allows for both crops to thrive (Andreotti et al., 2018; Blaser et al., 2018; Niether et al., 2020).
Cocoa grown under shade in some situations may be more resilient to climate change (Schwendenmann et al., 2010; Schroth et al., 2016). Schwendenmann et al. (2010) implemented drought experimentally in the field and found shade trees increased drought resilience. Shade trees insulate the understory crop from the warming and drying sun (Schroth et al., 2016). On the other hand, full-sun cocoa systems may be more climate resilient in some cases (Abdulai et al., 2018), as interactions between understory trees and shade trees are complex; in addition to shade effects, evapotranspiration and root interactions must be considered (Niether et al., 2017; Wartenberg et al., 2020). Moving to a full-sun system may also involve additional inputs in irrigation, fertilizer and labour. Neither (2020) reviewed the literature comparing the two cocoa production systems and concluded that the agroforestry system was superior in terms of climate adaptation.
The choice of cropping system will have wide-reaching consequences for climate vulnerability and climate justice. Coffee and cocoa are often a main source of income for small-scale producers who are among the most vulnerable to climate hazards (Bacon et al., 2014; Schroth et al., 2016). Most of their produce is exported by large corporations and sold to relatively better-off consumers. In the context of climate justice, underlying structural inequities (socioeconomic, ethnicity, gender, caste), marginality and poverty help to shape the vulnerabilities of small-scale farmers to climate hazards (Beckford and Rhiney, 2016; Schreyer et al., 2018). Climate change may compound their vulnerability, if for example the loss of pollination services leads to a reduction in productivity (Avelino et al., 2015). Adaptation needs to consider the inequities associated with the commodity chain, and the adaptative capacity of producers as they seek to move into the more advanced processing stages of the commodity chain to realise higher returns from their exports (Ovalle-Rivera et al., 2015). Blue Mountain Coffee is a ‘specialty’ coffee associated with a protected area forest ecosystem that attracts a high price premium owing to its distinct flavour and aroma. The livelihoods of coffee farmers in this region are characterised by multiple socioeconomic, environmental and institutional stressors related to climate change, pests, plant diseases and production costs. Some coping strategies employed by these coffee farmers have increased their susceptibility to future climate impacts (Guido et al., 2019). Davis (2017) showed that these coffee farmers’ food security challenges could be alleviated by improved marketing of fruit tree products under shade coffee farming systems. Adaptation measures in such systems need to consider co-benefits and negative trade-offs, especially in vulnerable communities, to avoid widening further the inequities, rural livelihood loss, migration and marginalisation, and ensure progress towards the SDGs (high confidence).
5.11.1 Current and Future Climate Change Impacts on Food Safety
Emerging food safety risks from climate change include those posed by toxigenic fungi, plant- and marine-based bacterial pathogens, HABs and increased use of chemicals (plant protection products, veterinary drugs) potentially leaving residues in food (European Food Safety Authority Panel on Plant Protection Products and their Residues et al., 2017; Deeb et al., 2018; Mbow et al., 2019; FAO et al., 2020).
Mycotoxins, produced by toxigenic fungi found on many crops, contaminate food and feed and cause a wide range of adverse impacts to human and animal health. Climate change can affect the growth and geographical expansion of these fungi (high confidence) (Wild et al., 2015; Battilani, 2016; FAO and WHO, 2016; Watson et al., 2016b; Alshannaq and Yu, 2017; Chen et al., 2018a; Avery et al., 2019; Milicevic et al., 2019; Van der Fels-Klerx et al., 2019; FAO, 2020a; FAO et al., 2020).
Aspergillus flavus is a fungus that infects a range of crops and can reduce grain quality. Several strains also produce aflatoxin, a particularly problematic mycotoxin . Increasing CO2 and drought stress has little effect on growth of Aspergillus but significantly increases the production of aflatoxin (Medina et al., 2015b).
In Europe, one estimate is that the risk of aflatoxin contamination will increase in maize in a +2°C temperature scenario in Europe, with nearly 40% of Europe exceeding the current legal limits (Battilani and Toscano, 2016). In Malawi, maize aflatoxin levels above European Union (EU) legal thresholds are possible for most of the country by mid-21st century (Warnatzsch and Reay, 2020). The occurrence of toxin-producing fungi will increase and expand from tropical and subtropical areas into new regions and where appropriate capacity for surveillance and risk management is lacking (medium confidence) (Miller, 2016). The increase in toxigenic fungi in crops, and consequent contamination of staple foods with mycotoxins, will increase the risks of human and animal exposure (high confidence) (Botana and Sainz, 2015; Rose and Wu, 2015 ; Battilani, 2016; Avery et al., 2019; Bosch et al., 2019; Milicevic et al., 2019; Moretti et al., 2019; Van der Fels-Klerx et al., 2019; FAO, 2020a).
In aquatic systems, toxins produced during HABs also cause food safety problems (high confidence) (Botana, 2016; Estevez et al., 2019; Section 5.8). Increased poleward expansion of Vibrio in coastal mid- to high-latitude areas has been observed (Baker-Austin et al., 2017). Vibrio-related mortalities from finfish consumption are expected to rise with climate change (water temperature, salinity, oxygen and pH) (medium confidence) (Mohamad et al., 2019a; Mohamad et al., 2019b). For shellfish species, oxygen deficits (Mohamad et al., 2019b), sea level rise (Deeb et al., 2018) and temperature (Green et al., 2019) will be most important for food safety.
Food safety is also anticipated to worsen from increased contaminant bioaccumulation under climate-induced warming (high confidence) (Sections 3.5.8, 3.5.9, 5.8, 5.9, Bindoff et al., 2019;), with changes in pathogen, parasite, fungi and virus abundance and virulence (Bondad-Reantaso et al., 2018). Coastal communities who depend on fisheries for livelihoods and nutrition are especially vulnerable (Hilmi et al., 2014; Golden et al., 2016; Bindoff et al., 2019).
Occurrence of bacterial pathogens such as Salmonella and Campylobacter will increase with rising temperatures (high confidence). Foodborne pathogen risks will increase through multiple mechanisms, though in general the impacts of climate change on different pathogens are uncertain (Akil et al., 2014; Hellberg and Chu, 2016; Lake and Barker, 2018). Even species within a genus can be affected differently. For example, higher CO2 levels depress the growth rate of F. graminearum, an economically important pathogen on barley but have little effect on F. verticillioides, which is the most reported fungal species infecting maize.
Increases in rainfall intensity will have some effect on the transport of heavy metals by enhancing runoff from soil and increasing the leaching of heavy metals into water systems, with magnitudes dependent on local conditions (high confidence) (Joris et al., 2014; Wijngaard et al., 2017). Methyl mercury (MeHg) is highly neurotoxic and nephrotoxic and bioaccumulates and biomagnifies through the food web via dietary uptake (fish, seafood, mammals) (Fort et al., 2016). Ocean warming facilitates methylation of mercury, and the subsequent uptake of methyl mercury in fish and mammals has been found to increase by 3–5% for each 1°C rise in water temperature (Booth and Zeller, 2005; FAO, 2020a). A changing climate will release mercury from snow and ice, raising the amount of mercury in aquatic ecosystems, although its importance relative to industrial sources is unknown (Morrissey et al., 2005).
Increased frequency of inland floods has been associated with contamination of food with toxic and fat-soluble persistent organic pollutants (POPs), polychlorinated biphenyls (PCBs) and dioxins (Lake et al., 2014; Tirado, 2015; Alava et al., 2017). Exposure to POPs can lead to serious health effects, including certain cancers, birth defects and impairments to the immune, reproductive and neurological systems.
Climate change–contaminant interactions may alter the bioaccumulation and biomagnification of POPs and PCBs as well as MeHg (Alava et al., 2017). Of particular concern is the pollution risk influenced by climate change in Arctic ecosystems and the bioamplification of POPs and MeHg in seafoods resulting in long-term contamination of traditional foods in Indigenous communities (Tirado, 2015; Alava et al., 2017).
The high risk associated with emerging zoonoses (animal diseases that can infect humans) and alterations in the distribution, survival and transmission of vectors and associated pathogens and parasites could lead to an increased use of veterinary drugs and more rapid development of microbial resistance (European Food Safety Authority et al., 2020; FAO, 2020a) and higher veterinary drug residues in food of animal origin, potentially posing health issues for humans (Beyene et al., 2015; FAO et al., 2018; European Food Safety Authority et al., 2020). These outcomes will depend, at least in part, on the extent of changes in current regulatory systems for veterinary drugs. Pre-harvest stress on animals can increase the contamination of meat products with zoonoses. Climate change may also increase rodent populations and rodent-born zoonoses (Naicker, 2011). Extreme weather events that cause flooding, such as hurricanes or extreme rain events, increase the chance of inundating areas that contain waste from animal farms where antibiotics are used for production, increasing the spread of antibiotic-resistant bacteria into the surrounding environment (FAO, 2020a).
5.11.2 Current and Future Climate Change Impacts on Food Loss in Storage, Distribution and Processing
The potential for climate-change-based food losses exists in all parts of the food system—post-harvest storage, distribution and processing—with the potential for impacts in one part of the system to be passed on to other elements (Davis et al., 2021). Storing a product destined for food use makes it available in times other than immediately after harvest, which is especially important for products with a pronounced seasonal availability or that are not available from other regions with different seasons. Storage of fresh products (meat, fish, fruits and vegetables) even with the best cold storage technology results in some quality loss relatively quickly. Higher temperatures increase the cost of maintaining quality. One estimate is that an increase in outdoor temperature from 17°C to 25°C increases cold storage power consumption by about 11% (James and James, 2010). Post-harvest storage of roots and cereals is subject to physical and quality losses from damage by mice, rats and birds and by microorganisms such as the toxigenic fungi discussed above, all of which are expected to increase in warmer and more humid conditions.
The higher temperatures and humidity will generally raise storage costs and lower the quantity and quality of stored product, reducing producer incomes and raising consumer prices (high agreement , medium evidence) (Mbow et al., 2019). For example, in the US state of Michigan, climate change will shorten the period of reliably cold local storage of potato by 11–17 days and 14–20 days further south by mid-century and by 15–29 days and 31–35 days, respectively, by late century. These changes would increase future demand for ventilation and/or refrigeration immediately after harvest and again in spring and early summer (Winkler et al., 2018).
Insects are a main source of food loss. Climate change can alter insect damage in at least two ways: increases in reproductive rate from temperature increases and changes in pheromone effectiveness (high confidence). Increasing temperature up to about 40°C raises the rates of insect food digestion and reproduction (Deutsch et al., 2018), but temperatures above that level are fatal for many insects (Neven, 2000). Most insects rely on pheromones to facilitate reproduction. Higher temperatures, but also increases in atmospheric CO2 and O3 levels, can affect this process. Insect species that rely on long-range chemical signals (such as ladybirds, aphids, bark beetles and fruit flies) will be most impacted, because these signals suffer from longer exposure to processes that reduce pheromone effectiveness (Medina et al., 2015b; Moses et al., 2015; Boullis et al., 2016; Verheecke-Vaessen et al., 2019).
There are several potential pathways for climate change impacts on processing that would negatively affect quality and appearance, but with limited research to date. For example, some studies have indicated that recent increases in temperature have decreased the appearance and milling quality of rice in the USA and East Asia, owing to increased occurrence of chalky grains (Lyman et al., 2013; Morita et al., 2016; Masutomi et al., 2019; Ishigooka et al., 2021). Impacts on quality of perennial crops and annual fruits and vegetables are discussed above (Section 5.4.3 and Box 5.2).
5.11.3 Current and Projected Impacts on Transportation and Distribution: Domestic and International Trade
Regional differences in resource availability are a key underlying driver of domestic and international trade. Climate change can change resource availability, in terms of both quantity and quality, altering trade flows, prices and incomes of producers. Climate change can also affect food access, and its stability can be affected through climate-change-driven disruption of infrastructure (FAO et al., 2018; Mbow et al., 2019). Extreme events are expected to become more common as climate change progresses. Recent examples illustrate the potential for trade disruptions. In March 2019, Cyclone Idai affected 1.7 million people in Mozambique and 920,000 in neighbouring Malawi, according to United Nations (UN) officials. The World Food Program reported that satellite imagery of flooding in central Mozambique showed an ‘inland ocean’ the size of Luxembourg with potentially large impacts on distribution of existing supplies, and uncertain effects on future food production and availability. The extreme rainfall events in the US state of Iowa in spring 2019 destroyed large numbers of well-built grain silos. In addition, major road and bridge damage required rebuilding.
Trade plays a sizeable role in global food supplies. More than 1 billion people relied on international food trade in the early 21st century (Fader et al., 2013; Pradhan, 2014). Domestic and international trade flows can be dramatically affected by climate change impacts (medium evidence, high confidence) (Nelson et al., 2014; Pradhan, 2014; Wiebe et al., 2015). Since the impacts of climate change will not be uniform, profitable locations for exports production will change. In addition, the effects of increasing local weather variability caused by climate change means increasing variability of food availability for domestic use and international trade. Finally, extreme events driven by climate change can disrupt transportation along the food value chain. Countries more at risk of natural hazards that disrupt transportation and distribution, and with less extensive routes, are more vulnerable to climate change impacts. A global multi-hazard risk assessment (Koks et al., 2019) suggests surface and river flooding, which are projected to increase in a warmer climate, are the main hazards for road and railway infrastructure, increasingly disrupting international and domestic transportation of agricultural commodities.
Climate change impacts will increase most global prices relative to early 21st century levels, with varying effects on the cost of food imports (high confidence) (Nelson et al., 2014; Wiebe et al., 2015; Fujimori et al., 2018; Lee et al., 2018). For example, analysis using results from one study (using CMIP5 data for RCP8.5 and SSP2) found that net food importing countries in the early 21st century would see expenditures on food imports decrease by USD 36 billion in mid-century in real terms with climate change over a no climate change scenario. (Table 5.13).
Table 5.13 | Net exports of agricultural products, by net exporting and net importing countries, 2010 and 2050 (billion constant parity US dollars), based on analysis in Beach et al. (2019).
2010 | 2050 | |
Net importers in 2010 | ||
No climate change | –301 | −838 |
Climate change | −301 | −802 |
Global economic models with a focus on agriculture provide a perspective on the range of potential changes in market outcomes because of climate change. In one study comparing several SSPs to a future with no climate change to one with impacts from RCP8.5, 2050 yields with climate changes impacts are 17% smaller on average than those without climate change. Adaptation by farmers reduce that to an 11% decline. The change in 2050 prices of all crops and regions after climate change impacts and farm-level adaptation is a mean 20% increase (Nelson et al., 2014). Substantial differences arise from both the heterogeneous impacts of climate change over crops and geography and the diversity of modelling approaches in the GCM and crop models. A later study with more socioeconomic scenarios and fewer models got roughly similar results (Wiebe et al., 2015), as did a modelling study focused on food security in South Asian countries (Cai et al., 2016).
Most climate scenario modelling to date does not incorporate increasing variability nor the use of storage, a critical tool to manage variability. Two recent studies are exceptions. In one, climate change generally reduces mean yields and increases their variability in the Midwestern USA and causes modest increases in price volatility (Thompson et al., 2018). A second study (Chen and Villoria, 2019) focuses on maize net importers across Africa, Asia and Latin America during 2000–2015. A 1% increase in the ratio of imports to total consumption reduces domestic price variability by 0.29%. A 1% increase in stocks at the beginning of the season is correlated with a 0.22% reduction in the coefficient of variation.
5.11.4 Adaptation in the Post-harvest Supply Chain
The SRCCL (Mbow et al., 2019) findings on adaptation support targeting food value chains and intervention types to the needs of specific locations. Furthermore, adaptation choices will need to be dynamic as climate change impacts are expected to worsen over time.
As discussed above and in Section 6.2.5, climate change is expected to cause increasingly severe effects on infrastructure needed for food security: roads and harbours for transport, water storage facilities for irrigation and storage facilities able to withstand climate-related damage. Three categories of adaptation could be considered: adoption of technologies already in use elsewhere, including Indigenous and local knowledge, or available or near ready that become profitable as impacts become more severe; development of new technologies; and taking advantage of changing comparative advantage across regions. Specific examples of post-harvest technical adaptation options that are already available but could be more widely adopted include solar driers, cold storage facilities and transport and use of ultrasonic humidification of selected fruits and vegetables, a technology that has been shown in Europe to reduce losses in each post-harvest stage by 20% or more (Fabbri et al., 2018). Hermetic storage containers using community-based farmer research networks to scale out (Singano et al., 2020; Wenndt et al., 2021) also show promise. Another innovation is to introduce Aspergillus fungi that do not produce aflatoxins in biocontrol formulations, as is being undertaken in the Aflasafe project in Kenya (Bandyopadhyay et al., 2016).
International trade changes are a potentially important adaptation mechanism for both the short-term effects of climate variability and long-term changes in comparative advantage with globally substantial benefits but that are distributed unevenly (Mosnier et al., 2014; Baldos and Hertel, 2015; Fuss et al., 2015; Costinot et al., 2016; Hertel and Baldos, 2016; Gouel and Laborde, 2021). One estimate is that, with a reduction in tariffs as well as institutional and infrastructural barriers, the negative impacts of climate change globally would be reduced by 64%, with hunger-affected import-dependent regions seeing the greatest benefit. However, in hunger-affected export-oriented regions, partial trade integration might lead to increased exports at the expense of domestic food availability (Janssens et al., 2020). It is possible for policy changes that result in increased trade flows to also increase the potential for maladaptation, for example by encouraging conversion of environmentally sensitive areas to agriculture (Fuchs et al., 2020; 5.13.3).
As discussed in Section 5.4, climate change is expected to increase variability in yields. As long as the variability is not correlated across regions, trade flows within a year can partially compensate, with in-period exports from countries less affected to those that are. Alterations in trade flow patterns to accommodate these impacts will reduce the negative effects so long as this variability is not correlated across regions (UK, 2015; Janetos et al., 2017).
In terms of food safety impacts, Lake and Barker (2018) highlight a range of approaches to enhance preparedness for more serious foodborne disease effects from climate change: adoption of novel surveillance methods to speed up detection and improve intervention in foodborne outbreaks; genotype-based approaches to surveillance of food pathogens to enhance spatiotemporal resolution in tracing and tracking of illness; improving integration of plant, animal and human surveillance systems under the rubric of One Health, increased commitment to cross-border and global information initiatives; and improved clarity regarding the governance of complex societal issues such as the conflict between food safety and food waste and strong user-centric (social) communications strategies to engage diverse stakeholder groups.
The range of potential adaptation approaches from production to transportation to reduce food loss and waste is captured in Figure 5.17 (Galford et al., 2020).
Figure 5.17 | Examples of food loss and waste (FLW) interventions at five stages in the food value change (Galford et al. , 2020).
The importance of reducing food loss and waste due to climate change is widely recognised, but literature on cost-effective reductions is sparse, particularly in low-income countries (Parfitt et al., 2010). A list of farm and post-harvest methods to reduce food loss (Sheahan and Barrett, 2017) includes potential farm interventions such as varietal choice, education in harvest and post-harvest handling, hermetic storage technologies (see above), chemical sprays and integrated pest management techniques in storage. The evidence on their effectiveness, especially in the face of increased climate change impacts, is limited.
5.12 Food Security, Consumption and Nutrition
5.12.1 Introduction
Food security and nutrition are key desired outcomes of food systems. Climate change is already contributing to reduced food security and nutrition and will continue to do so (high confidence) (Sections 5.4, 5.5, 5.8, 5.9, 5.10). Climate change impacts affect all four dimensions of food security: availability, access, utilisation and stability (Table 5.14), through both direct and indirect pathways.
Table 5.14 | Impacts from climate change drivers on the four dimensions of food security. Adapted from Table 5.1 in SRCCL.
CIDs and mechanism for food security impacts | Examples of regions and groups most affected | References |
Food security dimension: Availability | ||
Increased heat and drought reduce crop and animal productivity and soil fertility and increase land degradation for some regions and crops. | Countries in which a large proportion relies on agriculture for livelihoods. Food production systems that rely on rainfed agriculture and pastoral rangeland. Urban populations and the poor. | FAO et al. (2018), Dury et al. (2019), Mbow et al. (2019), Section 5.4 and 5.5). |
Extreme heat affects crop productivity. Combined with high humidity reduces agricultural labour capacity and animal productivity. | Countries and sectors that rely extensively on outdoor manual agricultural labour and experience high temperatures and humidity | Zander et al. (2015), Kjellstrom et al. (2016), Ioannou et al. (2017), Mitchell et al. (2017), FAO et al. (2018), Flouris et al. (2018), Kjellstrom et al. (2018), Levi et al. (2018). |
Increasing temperatures and precipitation changes increase and shift crop and livestock pests and diseases | East African pastoral groups who experienced increased livestock morbidity and mortality from RVF in El Niño years. | Bebber (2015), FAO et al. (2018), Mbow et al. (2019), Sections 5.4.1.3 and 5.5.1.3 |
Increasing temperatures and drought stress has led to higher post-harvest losses due to mycotoxins. | Tropical and subtropical regions with limited food safety surveillance | |
Rising ocean temperatures, marine heatwaves and ocean acidity has reduced availability of fish in coastal communities. | Coastal people and coastal areas of tropical countries with high dependence on fisheries, e.g., West African coastal communities | Hilmi et al. (2014), Golden et al. (2016), Bindoff et al. (2019), Section 5.8 and 5.9 |
Increased number and intensity of extreme events such as cyclones lead to reduced food production and distribution from crop damage, increased pest incidence and transportation disruption. | Delta regions where there are high populations and are often important food production regions, e.g., Cyclone Nargis in Myanmar estimated to reduce crop production by 19%, production declined for subsequent 3 years. | |
Increased atmospheric CO2concentrations increase total plant biomass and plant sugar content, which can increase crops as well as pests and weeds. High CO2 also reduces transpiration during drought, which can increase plant drought resistance. | All regions are anticipated to have increased atmospheric CO2 concentrations, but due to impacts of other CIDs (e.g., drought, heat stress, pests), the impacts on crop growth, forage and subsequent food availability are mixed. | Iizumi et al. (2018); Canadell et al. (2021), Ranasinghe et al. (2021), Cross-Chapter Box MOVING PLATE this chapter) |
Food security dimension: Access | ||
Increased drought and flood events and increased pests and disease from rising temperatures lead to loss of agricultural income due to reduced yields, and higher costs of production inputs such as water. Reduced ability to purchase food leads to lower dietary diversity and consumption levels. | Low-income smallholder farmers and pastoralists in Ethiopia, Mali, Niger, Malawi, Zambia and Tanzania. | Saronga et al. (2016), Giannini et al. (2017), FAO et al. (2018) Mbow et al. (2019) Omori et al. (2020) |
Increase in number and intensity of extreme weather events (e.g., droughts, floods) lead to increased food prices, which often leads to lower dietary diversity as well as lower consumption levels. | Low-income consumers. Women and girls. | FAO et al. (2018), Mbow et al. (2019), Ilboudo Nébié et al. (2021) |
Extreme events (e.g., floods) disrupt food storage and transport networks, reducing access and availability of food supplies. | Countries dependent on food imports, e.g., Small Island Developing States. Poor households living in flash flood and saline zones in Bangladesh who rely on monocropped rice. Women and children may experience greater impacts from extreme events. | Toufique and Belton (2014), FAO et al. (2018), Hickey and Unwin (2020), Algur et al. (2021) |
Food security dimension: Utilisation (food quality and safety) | ||
Increased temperatures reduce food safety caused by microorganisms, including increased mycotoxins in food and feed. | Countries with limited food safety surveillance systems. | |
Climate change extreme events make fruits and vegetables relatively unaffordable compared with less-nutrient-dense foods. | Urban low-income households and rural households who purchase the majority of their food. Children in regions such as West Africa, with lower access to diverse food types as a result of climate impact drivers, e.g., drought. | An et al. (2018), Algur et al. (2021), Baker and Anttila-Hughes (2020), Niles et al. (2021) |
Rising air temperature, ocean warming and high CO2conditions increase risk of food poisoning and pollutant contamination of food through increased prevalence of pathogens, HAB and increased contaminant bioaccumulation and threaten human health. | Low-income tropical countries where current ability to reduce and monitor mycotoxin contamination is limited. Coastal Indigenous Peoples and other poor populations in coastal areas of tropical countries with high dependence on fisheries, e.g., west African coastal communities | Golden et al. (2016), Bindoff et al. (2019), Sections 5.7, 5.8, 5.9, 5.11 |
Increased atmospheric CO2concentrations reduce nutritional quality of grains, some fruits and vegetables. | Low-income households who have limited access to range of diverse foods. | |
Rising ocean temperatures, marine heatwaves and ocean acidity reduce fish populations, which reduces consumption of fish high in iron, zinc, omega-3 fatty acids and vitamins in areas where fish populations decline. | Coastal areas of tropical countries; coastal Indigenous Peoples and other groups who rely on fisheries. | Golden et al. (2016); Bindoff et al., 2019; Section 5.7, 5.8, 5.9 |
Food security dimension: Stability | ||
Increased frequency and severity of extreme events (e.g., droughts and heatwaves) lead to greater instability of supply through production losses and disruption to food transport. | Landlocked countries; low-income countries reliant on imports; low-income households in areas prone to floods. | Toufique and Belton (2014), FAO et al. (2018), Algur et al. (2021), Section 5.11 |
Increased drought and flood events and increased pests and disease from rising temperatures lead to unstable incomes from agriculture and fisheries. | Small-scale producers (crops and livestock) and fishers | Ruiz Meza, (2015), FAO et al. (2018), Sections 5.8, 5.9 |
Climate change extreme events increase food prices due to climate shocks. | Low-income countries reliant on imports; urban low-income households and rural households who purchase the majority of their food. | Bene et al. (2015), Peri (2017), Mbow et al. (2019), Section 5.11 |
Increased drought and flood events and increased pests and disease from rising temperatures cause widespread crop failure. Rising ocean temperatures, marine heatwaves and ocean acidity lead to dramatic decline in fisheries, contributing to migration and conflict. | Coastal communities in West Africa, Southeast Asia and other tropical countries highly dependent on fisheries. | Golden et al. (2016), Bindoff et al. (2019) Mbow et al. (2019) |
Reduced frost days and snow days will increase stability of food security in some temperate regions since there will be less loss of food crops to frost damage and a longer growing season. However, they also raise pest and disease risks due to increased range and overwintering. | Australia, most Asian regions, Europe, Central and South America and North America. The benefits of yield gains at high latitudes may be tempered by greater risks of pests and pathogen damages. | Jones and Barbetti (2012), IPPC Secretariat (2021), Ranasinghe et al. (2021) |
Global food security improved dramatically in the 20th century even as global population increased from 2 to 6 billion. While some may assume that global food security is primarily provided by large-scale producers, research since AR5 has shown the sizeable role of small and mid-sized food producers in Asia, Africa and Latin America contributing to global food security and nutrition, while being highly vulnerable to climate change impacts on food security (Samberg et al., 2016; Herrero et al., 2017; FAO et al., 2018; Ricciardi et al., 2018). In 2019, more than 750 million people in the world, almost 1 in 10 people, suffered from severe food insecurity, a figure which has risen since 2014 in every region except North America and Europe
(FAO et al., 2020). Overnutrition, a result of high-calorie unbalanced diets, is also rising, with over 2 billion adults overweight or obese (FAO et al., 2018; Swinburn et al., 2019; FAO et al., 2020; Venkatesh Mannar et al., 2020; WHO, 2021). Many low- and middle-income countries now have both high under- and overnutrition rates (FAO et al., 2018).
There are multiple drivers of food security, including changing dietary patterns, urbanisation and population growth (HLPE, 2017b; FAO et al., 2018; Swinburn et al., 2019). Vulnerability to climate change impacts on food insecurity and malnutrition is worsened by other underlying causes, including poverty, multiple forms of inequity (e.g., gender, racial, income), low access to water and sanitation, macroeconomic shocks and conflict (Smith and Haddad, 2015; Clay et al., 2018; FAO et al., 2018; Cook et al., 2019). Climate change frequently acts to compound these drivers of food insecurity (Table 5.14).
The coronavirus disease 2019 (COVID-19) pandemic has increased vulnerability to food insecurity and malnutrition of particular groups and sectors in the food system, including low-income households, farmworkers, food service workers, informal food market sellers and low-income countries dependent on food imports (Cross-Chapter Box COVID in Chapter 7). Climate change will compound pandemic vulnerabilities in the food system (high agreement , low evidence) (HLPE, 2020; UNDRR (United Nations Office for Disaster Risk Reduction, Regional Office for Asia and Pacific), 2020; WFP-FSIN, 2020). The pandemic may also increase coordination among sectors and a willingness to address food system weaknesses made visible by the impacts of COVID-19 (Blay-Palmer et al., 2020; Cohen, 2020; Ramos et al., 2020).
Ecosystem services, the provisioning, supporting and regulating mechanisms we all depend on for food security and nutrition, are also undermined by climate change impacts (Section 5.4.3). Even in the absence of climate change, our current food system threatens to exceed planetary, regional or local boundaries of long-term sustainable development (Campbell et al., 2017). Climate change will make efforts to reduce this threat more difficult to achieve (medium confidence), though many solutions to enhancing food security are also potential climate change adaptation responses (Sections 5.4, 5.6, 5.8, 5.10, 5.14).
5.12.2 Mechanisms for Climate Change Impacts on Food Security
Climate change is increasing the number of people experiencing food insecurity through greater incidence and severity of climatic impact drivers (CIDs), (Seneviratne et al., 2021) such as extreme heat, drought and floods. Increasing CO2 concentrations have positive effects on food and forage crops by enhancing photosynthesis and alleviating drought stresses (5.4.3.1, 5.5.3.1) but have negative effects on nutrient concentrations in food crops. Ocean acidification is also caused by increasing CO2, causing negative impacts on aquatic systems. Tropospheric ozone concentrations already hinder crop production (Section 5.4.1.4). Several CIDs increase the number of people experiencing food insecurity (high confidence) (SROCC 2019, FAO et al., 2018; Mbow et al., 2019; Baker and Anttila-Hughes, 2020; Table 5.12).
Vulnerability to climate impacts on food security and nutrition varies by region and group. Countries that experience CIDs such as extreme heat, severe drought or floods and have a large proportion of the population dependent on rainfed agriculture or livestock for their livelihoods and food supply have experienced rising food insecurity due to climate change impacts (FAO et al., 2018; Cooper et al., 2019; Mbow et al., 2019). Children in Sub-Saharan Africa are particularly at risk of undernutrition and mortality from increasing temperatures (Belesova et al., 2019; Baker and Anttila-Hughes, 2020). An additional estimated 5.9 million children became underweight because of rising temperatures in 51 countries affected by ENSO intensity in 2015–2016 (Anttila-Hughes et al., 2021). Low-income urban households and marginalised groups such as landless and ethnic minorities are at risk of increased food insecurity due in part to climate change extreme events such as extended drought, floods or cyclones that interrupt supply chains and impact livelihoods (Rodriguez-Llanes et al., 2016; FAO et al., 2018; Algur et al., 2021). A systematic review in India found that women often experience greater workloads and stress during drought events (Algur et al., 2021).
In the subsequent sections, the four dimensions of food security will be discussed in relation to observed and projected impacts and vulnerabilities (Table 5.14).
5.12.3 Observed Impacts
5.12.3.1 Impacts on food availability
All food production systems (crops, livestock, marine, fish, mixed, aquaculture) have been undermined by climate change and are expected to experience larger impacts in the future as described in earlier sections (see Sections 5.4.1, 5.5, 5.8, 5.9, 5.10). In addition, sudden production losses from extreme climate events can reduce food security (FAO et al., 2018; Cottrell et al., 2019; FAO et al., 2020; Anttila-Hughes et al., 2021). For example, a 2007 drought-induced crop failure in southern Africa led to severe food insecurity in Lesotho because of the land-locked country’s dependence on imports from South Africa that aggravated food availability and access under conditions of declining food production and land degradation (Verschuur et al., 2021). Pest and disease outbreaks in both crops and livestock due to climate change (Sections 5.4.1, 5.5.1) have also impacted food availability and access (see Box 5.8 Desert Locust case study). Loss in labour productivity from climate-change-related heat stress is a growing problem.
Climate change affects agricultural labour productivity through increased intensity and frequency of heat stress events, with those performing physical labour in high humidity and ambient temperatures most vulnerable to heat stress (high confidence) (Hsiang et al.; FAO et al., 2018; Kjellström et al., 2019; Antonelli et al., 2020; Shayegh et al., 2020). Labour capacity, supply and productivity loss in moderate outdoor work due to heat stress is estimated between 2% and 14%, depending on the location and indicator (Ioannou et al., 2017; Kjellstrom et al., 2018), with an overall estimate of 5.3% loss in productivity for outdoor work between 2000 and 2015 (medium confidence) (Watts et al., 2018) but as high as 14% in low-income tropical countries (Antonelli et al., 2020; Shayegh et al., 2020). Highly vulnerable occupation groups affected by heat stress include farmers, farmworkers and livestock keepers working outdoors in low-income tropical countries (high confidence) (Zander et al., 2015; Kjellstrom et al., 2016; Flouris et al., 2018; Kjellstrom et al., 2018; Levi et al., 2018). Farmworkers and small-scale food producers in high- and middle-income countries involved in outdoor labour are also affected by heat stress (Zander et al., 2015; Gosling et al., 2018; Szewczyk et al., 2018; Watts et al., 2021). There is also evidence that heat stress is affecting labour supply through variation in nutrition intake (Antonelli et al., 2020).
5.12.3.2 Impacts on food access (physical, economic and socio-cultural) and vulnerabilities
Increased extreme events (e.g., droughts, floods and tropical storms; Seneviratne et al., 2021) due to climate change are key drivers of recent rises in food insecurity rates and severe food crises in some regions (high confidence) (Section 5.4.1, Yeni and Alpas, 2017; FAO et al., 2018; Cooper et al., 2019; Baker and Anttila-Hughes, 2020; Bogdanova et al., 2021; Ilboudo Nébié et al., 2021). Extreme weather events reduce physical and economic access to food, increase food prices, and compound underlying conditions of food insecurity and malnutrition such as low access to diverse healthy foods and safe water (FAO et al., 2018; Niles et al., 2021). Increased incidence of severe drought conditions since 2005 is contributing to food insecurity in affected regions, including Africa, Asia and the Pacific (Chapter 7, Phalkey et al., 2015; FAO et al., 2018; Cooper et al., 2019; Ilboudo Nébié et al., 2021; Verschuur et al., 2021;). In Arctic western Siberia, high temperatures, melting ice and forest and tundra fires have degraded reindeer pastures; Indigenous Peoples have reduced traditional diets and increased purchased food with increases in hypertension and related health impacts (Bogdanova et al., 2021).
There is growing evidence that anthropogenic climate warming has already intensified climate extreme events induced by large-scale SST oscillations such as ENSO (Herring et al., 2018; Seneviratne et al., 2021). For example, the 2015–2016 El Niño, the strongest in the past 145 years, induced severe droughts in Southeast Asia and eastern and southern Africa, some intensified by anthropogenic warming (Funk et al., 2018). As a result, 20.5 million people faced acute food insecurity in 2016 (FSIN, 2017) and an estimated additional 5.9 million children became underweight (Anttila-Hughes et al., 2021).
Weather extreme events increased food prices and food price volatility (Peri, 2017), thereby worsening food insecurity (Shiferaw et al., 2014; Bene et al., 2015; Miyan, 2015; FAO et al., 2018; Ilboudo Nébié et al., 2021). Rising food prices can affect conflict, political instability and migration (Bush and Martiniello, 2017), but the relationship between climate change, political instability and conflict is often mediated by other underlying factors such as poor governance (Chapter 7.2.7, Mach et al., 2019; Selby, 2019).
Low-income urban and rural households who are net food buyers are particularly affected by food price increases, with reduction in consumption of diverse food groups (high confidence) (Green et al., 2013; Villasante et al., 2015; FAO et al., 2018). Depending on the context, particular groups, including women, ethnic and religious minorities, will be more vulnerable to worsening food insecurity from climate change impacts (Clay et al., 2018; Jantarasami et al., 2018; Nature climate change Editorials, 2019; Algur et al., 2021 and see Cross-Chapter Box GENDER in Chapter 18). Indigenous Peoples are often more vulnerable to climate change, due to conditions of poverty, limited resources, discrimination and marginalisation (high confidence) (Smith and Rhiney, 2016; Vinyeta et al., 2016; Jantarasami et al., 2018). Indigenous Peoples may experience loss of culturally significant foods and declining traditional ecological knowledge (Dounias and Ichikawa, 2017; Ross and Mason, 2020; 5.7).
5.12.3.3 Impacts on food utilisation and vulnerabilities
Food utilisation refers to the way the body most effectively uses food, and includes food preparation, food quality and intra-household distribution. Food utilisation is affected by climate change in several ways: food safety, dietary diversity and food quality (Aberman and Tirado, 2014).
Climate change have increased food safety risks (high confidence), including foodborne zoonotic animal diseases (5.5), and marine toxins from HABs (Sections 5.8, 5.9) and mycotoxins (Section 5.11). Other foodborne and waterborne infectious diseases such as cholera are further covered in Chapter 7.
Weather variability and extreme events (Seneviratne et al., 2021) have reduced availability and access to diverse foods to sell and to purchase in rural markets, thereby reducing access to affordable, diverse foods for both rural small-scale producers and net consumers, particularly for landlocked and low-income countries (high confidence) (Pant et al., 2014; Villasante et al., 2015; Alston and Akhter, 2016; FAO et al., 2018; Park et al., 2019; Niles et al., 2021) and otherwise marginalised communities (Algur et al., 2021). One study of 87 countries and 150 extreme events estimated that low-income food deficit and landlocked countries had reduced nutrient supply ranging from −1.6 to −7.6% of average supply, a significant portion of a healthy child’s average dietary intake (Park et al., 2019).
Rural children in low-income countries are at particular risk of undernutrition from climate change impacts, due to a combination of factors: potential reduction in food quantity and quality from heat impacts; greater exposure from outdoor play and agricultural activities; and increased likelihood of heat exhaustion and vector-borne and diarrheal diseases (Oppenheimer and Anttila-Hughes, 2016). A study of child growth data in 30 countries in Africa between 1993 and 2012 found that increased temperature was significantly related to children’s wasting (Baker and Anttila-Hughes, 2020). Another study examined 30 years of climate data and child dietary diversity outcomes in 19 countries, and found that higher-than-average annual temperatures correlated with declines in child diet diversity at levels equal to or greater than other factors which often are the focus of policy, such as market access or education (Niles et al., 2021).
5.12.3.4 Impacts on food stability
Climate change has already changed the start and duration of the growing season and increased variability of rainfall in some places, with impacts on food intake and nutritional status and income for low-income and small-scale producers (medium evidence, high agreement , (FAO et al., 2018; Cooper et al., 2019). Evidence to date suggests that climate change has negative impacts on the stability of food supply over the medium to long term, thereby affecting food stability (Myers et al., 2017b). Increasing number and intensity of adverse weather events, driven by climate change (Seneviratne et al., 2021), are important factors decreasing food stability, through reduced availability, increased local price volatility, reduced livelihoods for food producers and disruption to food transport (Toufique and Belton, 2014; Verma et al., 2014; Ruiz Meza, 2015; Clay et al., 2018; FAO et al., 2018; Mbow et al., 2019).
Box 5.9: Desert Locust Case Study: Climate as Compounding Effect on Food Security
At the end of 2019, desert locust swarms infested Eastern Africa and caused widespread damage to crops and pastures, threatening food security and livelihoods (Kimathi et al., 2020; Salih et al., 2020). The FAO estimates that over 200,000 ha of crop and pastureland were damaged, rendering 2 million people in the region acutely food insecure (IGAD, 2020). The desert locust infestation was facilitated by two tropical cyclones that created desert lakes in a usually dry region of Saudi Arabia. Moist soils, warm temperatures and ample vegetation provided a suitable environment for desert locust breeding and migration to Yemen and Somalia, where the pest remained uncontrolled due to conflict and spread to neighbouring countries. A series of political and socioeconomic weaknesses such as armed conflict, limited financial resources and lack of early actions compounded the impact of the current invasion and made it the most damaging in 70 years (Meynard et al., 2020; Salih et al., 2020).
Although desert locusts have been here for centuries, this recent outbreak can be linked to a unique feature of the positive IOD event, in part caused by long-term trends in SSTs (Wang et al., 2020a). The warming of the western Indian Ocean has increased frequency and intensity of severe weather, including tropical cyclones (Roxy et al., 2014; Murakami H, 2017; Roxy et al., 2017). Under a 1.5°C warmer climate, extreme positive IODs are anticipated to occur twice as often, which could also increase the occurrence of pest outbreaks (Cai et al., 2018).
Climate change increases the need for robust adaptation measures, such as transnational early-warning systems, biological control mechanisms, crop diversification and further technological innovations in areas of sound and light stimulants, remote sensing, and modelling for tracking and forecasting of movement (Maeno and Ould Babah Ebbe, 2018; Peng et al., 2020). The desert locust outbreak and the role of the Indian Ocean warming show that the impacts of climate change can increase unpredictable events. Extreme weather events act as a compounding effect, exacerbated further by weak governance systems, political instability, limited financial resources and poor early-warning systems (Meynard et al., 2020).
5.12.4 Projected Impacts on Food Security
5.12.4.1 Food availability and access
Climate change will have negative effects on food security and nutrition in 2050 (high agreement , medium evidence) (Amjath-Babu et al., 2016; Springmann et al., 2016; Lloyd et al., 2018; Richardson et al., 2018; see Chapter 7; Hasegawa et al., 2021a). How many people are affected will depend considerably on non-climatic drivers of food security (van Dijk et al., 2021), but modelling studies agreed that climate change would increase the risk of food insecurity. For example, one study comparing an RCP8.5 scenario with one that has zero climate impacts estimates 65 million additional people (10% increase) will experience food insecurity due to climate change impacts in 2050 (modelling results in Nelson et al., 2018). Another study accounting for climate extreme events estimates that, by 2050, the number of people at risk of hunger will increase by 20% and 11% under high- and low-emission scenarios, respectively, owing to a once-per-100-year extreme climate event (Hasegawa et al., 2021a). Sub-Saharan Africa and South Asia in this study were projected to be at the greatest risk, with triple the amount of South Asia’s current food reserves needed to offset such an extreme event. Models suggest that food security and malnutrition impacts will be much more severe from 2050 onwards relative to pre-2050, but the scale and extent of the impacts will strongly depend on the GHG emission scenario (FAO, 2018a; Richardson et al., 2018). Due to CIDs and non-climate drivers of food insecurity, Sub Saharan Africa is projected to be the hardest hit, followed by South Asia and Central and South America, but contingent on adaptation level (Richardson et al., 2018; Hasegawa et al., 2021a).
Without adaptive measures, heat stress impacts on agricultural labour will increase with climate change (high confidence) (Im et al., 2017; Levy and Roelofs, 2019; Hertel and de Lima, 2020). Climate-change-related heat stress will reduce outdoor physical work capacity on a global scale. Depending on GHG concentrations, some regions will experience losses of 200–250 outdoor workdays per year at century’s end. Using results from one study reporting experimental procedures to assess loss of work capacity (Foster et al., 2021), regions hardest hit in an SSP5-8.5 scenario include much of South Asia, tropical Sub-Saharan Africa and parts of Central and South America (Figure 5.18). de Lima et al. (2021) projected that negative impacts of warming on crop yields and labour capacity would affect crop production and cost for workers and labour-saving mechanisation, raising food price by 5% at +3° from the baseline period (1986–2005) globally, with significant implications for vulnerable regions (sub-Saharan Africa and Southeast Asia). Large uncertainties, however, exist around population diversity and adaptive capacity (Vanos et al., 2019). Agricultural labour productivity impacts of heat attributed to climate change are expected to be worse in low- and middle-income countries (Kjellstrom et al., 2016). Adaptation options needed to protect agricultural worker productivity outdoors and reduce occupational heat illnesses and deaths include cooled working environments, improved surveillance systems and education on the need to monitor (high confidence) (Xiang et al., 2016; Quiller et al., 2017; Flouris et al., 2018; Day et al., 2019; Vanos et al., 2019). Currently available options, however, are more difficult to achieve in lower-income economies (Kjellstrom et al., 2016; Im et al., 2017).
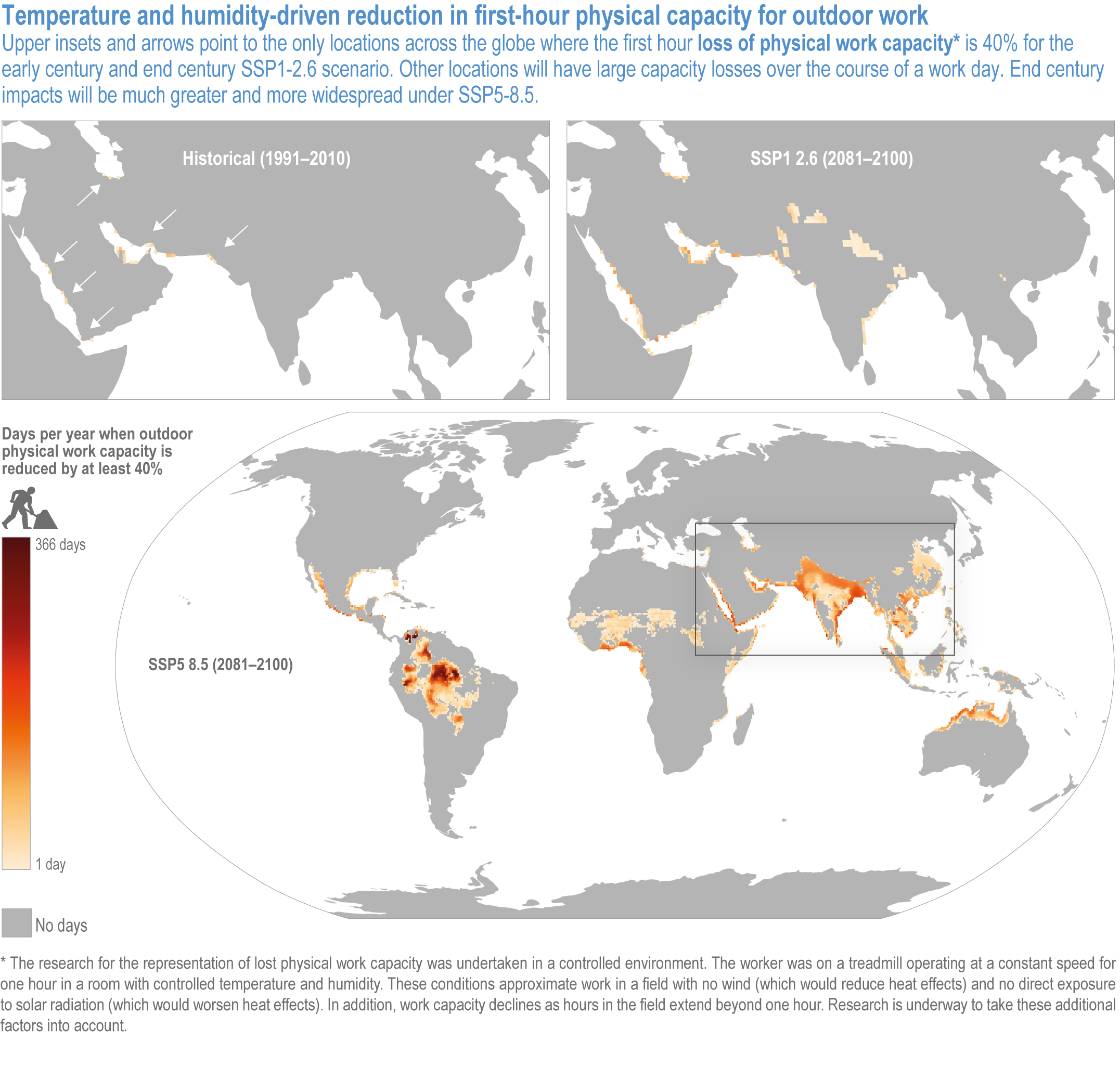
Figure 5.18 | The number of days per year where physical work capacity (PWC) is less than 60% based on average daily air temperature and relative humidity (Foster et al. , 2021). PWC is defined as the maximum physical work output that can be reasonably expected from an individual performing moderate-to-heavy work in a ‘cool’ reference environment of 15°C. Values plotted are from the early (A) and end of century (B) for SSP5-8.5 using ensemble means from the ISI-MIP CMIP6 data set. See SM5.4 for details.
Under higher-emission scenarios, food availability will be further reduced after 2050, due to the potential for widespread crop failure and decline in livestock and fisheries stocks (Mbow et al., 2014; Kelley et al., 2017; Challinor et al., 2018; Hendrix, 2018; Bindoff et al., 2019). At +3°C from the preindustrial era, all food production sectors will experience greater, and pronounced, losses due to climate change compared with +1.5°C or +2°C (see Sections 5.2, 5.4.3, 5.8.3 and 5.9.3).
Food insecurity from food price spikes due to reduced agricultural production associated with climate impact drivers such as drought can lead to both domestic and international conflict, including political instability (Abbott et al., 2017; Bush and Martiniello, 2017; WEF, 2017; D’Odorico et al., 2018; de Amorim et al., 2018;Chapter 7.2.7). While climate change impacts, including drought impacts on food security, are important risk factors for conflict, other key drivers are often more influential, including low socioeconomic development, limited state capacity, weak governance, intergroup inequities and recent histories of conflict (medium confidence) (Mach et al., 2019; Selby, 2019; Chapter 7.2.7). The interaction between extreme weather events, conflict and human migration may increase vulnerability of particular communities of low-income countries (WEF, 2017; D’Odorico et al., 2018; de Amorim et al., 2018; Chapter 7). Further research is needed to better understand how increased drought risk under future climate change might affect food prices and water availability (Abbott et al., 2017).
5.12.4.2 Projected Impacts on Food Safety and Quality
Increasing levels of CO2 directly contribute to reduced food quality by reducing levels of protein, iron, zinc and some vitamins, varying by crop species and cultivars (high confidence) (Section 5.4.3, Myers et al., 2014; Smith and Haddad, 2015; Bisbis et al., 2018; Scheelbeek et al., 2018; Weyant et al., 2018; Zhu et al., 2018a). Higher levels of CO2 are predicted to lead to 5–10% reductions in a wide range of minerals and nutrients (Loladze, 2014). Climate warming will also reduce food quality of seafood, by changing the LC-PUFA content in phytoplankton (Section 5.8; Hixson and Arts, 2016).
5.12.4.3 Reaching Sustainable Development Goal 2
Current projections indicate that it is highly likely that the UN SDG2 (‘Zero Hunger’) by 2030 will not be achieved, with climate impacts on one of several drivers of food security and nutrition preventing this goal, including in Africa, Small Island States and South Asia (high confidence) (FAO et al., 2018; Otekunrin et al., 2019; Singh et al., 2019; Atukunda et al., 2021; Kumar et al., 2021; Vogliano et al., 2021). Integrated policy strategies that consider synergies and trade-offs between different food system components would strengthen the likelihood of meeting SDG2 goals (Dyngeland et al., 2020; Lipper et al., 2020; Vogliano et al., 2021) (Grosso et al., 2020). Adaptation options which address climate risks for food security and nutrition are discussed below.
Box 5.10: Food Safety Interactions with Food Security and Malnutrition
Climate change significantly increases the future food safety risks (high confidence) (Sections 5.8.2, 5.8.3, 5.11.1, Box 5.9). Increasing temperatures and drought stress are expected to lead to greater aflatoxin contamination of food crops. Aflatoxins, a major foodborne hazard, contaminate staple crops and are associated with various health risks, including stunting in children and cancer (Koshiol et al., 2017). In LICs, children with high exposure to aflatoxins were found to be more likely to suffer from micronutrient (zinc and vitamin A) deficiencies (Watson et al., 2016b). Climate change is expected to cause decreases in micro- and macronutrient content of foods, leading to an increased burden of infectious diseases, diarrhea and anaemia, with an estimated 10% increase in disability-adjusted life years (DALYs) by 2050 associated with undernutrition and micronutrient deficiencies (Aberman and Tirado, 2014; Smith and Myers, 2018; Weyant et al., 2018; Zhu et al., 2018a; Ebi and Loladze, 2019; FAO, 2020a; Sulser et al., 2021b).
Children in low-income countries will be at greater risk of undernutrition from these multiple climate change impacts, including lower food availability, quality and safety and increased risk of diarrheal disease (high confidence) (Aberman and Tirado, 2014). One study of 30 countries in Africa estimated that, by 2100, increased temperatures under RCP8.5 could increase children’s wasting by 37% in western Africa and 25% in southern Africa (Baker and Anttila-Hughes, 2020).
The combination of climate change and the presence of arsenic in paddy rice fields is expected to increase the toxic heavy metal content of rice and reduce production by 2100, threatening food security and food safety mainly in low-income countries where rice is the main staple (Neumann et al., 2017; Muehe et al., 2019; Farhat et al., 2021).
Table 5.17 | Examples of adaptation responses to drought and floods by food security level and time frame. Adapted from Ilboudo Nébié et al. (2021) Table 4, with information from Bahadur et al. (2015); Costella et al. (2017); Gros et al. (2019); Ulrichs et al. (2019); Medina Hidalgo et al. (2020); Bacon et al. (2021); and Verschuur et al. (2021).
Food insecurity level and time frame of adaptation | ||||
Adaptation response to drought or floods | Acute, short term | Moderate, medium term | Chronic, long term | Resilience type |
Forecast-based financing (provides unconditional cash in advance of extreme event) | X | Anticipatory: people and systems are better prepared for climate shock by reduced exposure or vulnerability. | ||
Early-warning systems/climate services and education for disaster preparation | X | X | X | |
Social protection programmes with regular provisions which allow for asset building, e.g., savings, building of informal networks, purchase of livestock | X | X | ||
Humanitarian food aid and malnutrition treatment | X | X | Absorptive capacity: people or systems cope with climate-related shocks or systems while and immediately after they occur. | |
Home-grown nutrition-sensitive school feeding programmes | X | X | ||
Social protection programmes with short-term targeted response, e.g., short-term cash transfers, food assistance for asset building such as wells | X | |||
Weather index insurance program | X | X | X | |
Regional grain banks run by farmer associations | X | X | Adaptive capacity: can adjust to long-term climate risks and disasters, reduce vulnerability to future shocks. | |
Savings, credit and local food procurement support for smallholder farmers | X | X | ||
Agroecosystem diversification, other agroecological practices to strengthen ecosystem services in long term (see Box 5.10) | X | X | ||
Rainwater evacuation infrastructure combined with flood management and waste collection and urban gardening | X | X | ||
Drought- or flood-resistant crop varieties | X | X | ||
Expand trade partners beyond climactically connected partners | X | X | ||
Gender transformative or responsive agriculture programmes | X | X | ||
5.12.5 Adaptation Options for Food Security and Nutrition
Since AR5, there has been increased research on adaptation options that address climate risks for food security and nutrition. In this section, cultivar improvements, urban and peri-urban agriculture, changing dietary patterns, integrated multi-sectoral approaches and rights-based approaches are assessed for their potential as an adaptation option that addresses food security and nutrition. Feasibility and effectiveness assessment of several options is in Section 5.1 4.
5.12.5.1 Potential, barriers and challenges for genetically modified crops to address food security and nutrition
While biotechnology can be used as an adaptation strategy (Section 5.4.4.3), there is low confidence that genetically modified (GM) crops can increase food security and nutrition in smallholder farming systems relative to alternative agronomic strategies (National Academies of Sciences Engineering and Medicine, 2016; Qaim, 2016). Some underline their potential in building resilience to changing climatic conditions, in the form of enhanced drought/heat tolerance, pest/disease protection and/or reduced land usage, thus serving to bolster food security and nutrition (Sainger et al., 2015; Muzhinji and Ntuli, 2021). Others suggest that the empirical evidence supporting GM crops as a climate-resilience strategy remains thin (Leonelli, 2018). Technical and social barriers and potential solutions are summarised in Table 5.15.
Table 5.15 | Barriers, challenges and potential solutions for GM crops.
Barriers and challenges | Examples and potential solutions to barriers |
Major challenges as a food security and nutrition adaptation include the introgression of GM traits into host varieties (Dowd-Uribe, 2014), and confusion around proper growing practices that can accelerate resistance (Iversen et al., 2014; Fischer et al., 2015). The combination of the kinds of traits and restrictions that come from the predominant intellectual property rights instruments used in their commercialisation, and concentration of plant and animal breeding industry (Bonny, 2017) mean that benefits from released GM crops tend to be captured disproportionately by farmers with more land, wealth and education (Afidchao et al., 2014; Ali and Rahut, 2018; Azadi et al., 2018) but also increase debt levels for growers (Dowd-Uribe, 2014; Leguizamón, 2014). Underlying gender inequities also play a critical role in shaping food security and nutrition outcomes associated with the introduction of GM crops, in part due to unequal control over income and agricultural decision making; in some cases, women reported decreased workload and enhanced decision-making power (Gouse et al., 2016), while in others the introduction of GM crops could increase workload and devalue womens’ role as seed savers (Carro-Ripalda and Astier, 2014; Addison and Schnurr, 2016). Major hurdles for GM crops include translating promising research results into real-world farming systems and consumer trust in the food product. Experimental programmes have been dogged by issues, including complications with the introgression of GM traits into high-performing varieties (Dowd-Uribe and Schnurr, 2016; Stone and Glover, 2017), strict management regimes that clash with the realities of smallholder agricultural systems (Iversen et al., 2014; Whitfield et al., 2015), and a lack of attention to farmer decision making (Schnurr, 2019). | One case study is the Water Efficient Maize for Africa (WEMA) programme, a public–private partnership that transplants a cold shock protein B, known as Droughtgard, into maize in order to mitigate yield losses from drought. Proponents suggest that this GM venture, which will be distributed free to smallholder farmers, represents the best strategy for ensuring stable yields in the face of climatic change across Africa (Kyetere et al., 2019). Critics argue that WEMA maize is not a good fit with the smallholder farming systems it is designed to benefit, with particular concerns around how farmers will access the extra inputs, credit and labour that WEMA maize requires to be successful (Schnurr, 2019). Emergent genome-edited crops are considered a more precise, accessible and accelerated means of targeting stressors that matter to poor farmers, but evidence is limited (Kole et al., 2015; Haque et al., 2018; Zaidi et al., 2019). A more iterative and flexible adaptation approach beyond just genomic improvement to tackle the multiplicity of factors limiting smallholder production is anticipated to increase the likelihood that these promising technologies can enhance food security and nutrition (medium confidence) (Giller et al., 2017; Stone, 2017; Montenegro de Wit, 2019). To address food security and nutrition, future breeding needs to move from just enhancing agronomic traits of a single crop to improving multiple traits of multiple crops suited to local conditions that will increase climate resilience of farming systems. To make breeding technologies scale-neutral, the policy structure needs to support and protect smallholders (medium confidence). |
5.12.5.2 Urban and peri-urban agriculture, vertical and horizontal
Urban areas have more than half of the global population and consume about 70% of the total food supply (FAO, 2019b). The urban population is projected to grow further to about 70% of the global population by 2050 (UN, 2018). Direct evidence supporting climate resilience of urban and peri-urban agriculture (UPA) is limited and contextual, but there is medium confidence of multi-functional benefits from UPA, depending on regions and types of UPA (Artmann and Sartison, 2018; Kareem et al., 2020). UPA takes different forms of production, and can be broadly classified into four categories, depending on operating characteristics and capital inputs (Table 5.16) (Goldstein et al., 2016). Controlled environments can protect crops, livestock and fish from extreme weather events or pest and disease outbreak (Mohareb et al., 2017). Innovative indoor farming such as vertical farming can be highly productive with minimal water and nutrient supply but can be capital intensive with high energy demand (O’Sullivan et al., 2019), and those with aquaponics can be water demanding (Love et al., 2015). Currently, commodities are often limited to crops with short growing seasons such as leafy vegetables. Vertically grown crops are more expensive than field-grown produce and, thus, not accessible for low-income urban dwellers (Al-Kodmany, 2018). Community and institutional unconditioned (outdoor) farms and gardens are better positioned to provide increased access to healthy food to those who need it (Eigenbrod and Gruda, 2015; Goodman and Minner, 2019).
Table 5.16 | Urban agriculture classifications based on operating characteristics and capital inputs (Goldstein et al., 2016; O’Sullivan et al., 2019), and a summary of literature search on positive and negative aspects.
Summary of urban and peri-urban agriculture and evidence for improved food security and nutrition | ||
Urban agriculture has two components: vertical (e.g., grown on or in buildings) and horizontal (grown on land within urban boundaries, in backyards and marginal spaces). The horizontal component of urban and peri-urban agriculture (UPA) has gained attention because of multiple functions that could improve food systems and ecosystem services under climate change (Revi et al., 2014; Artmann and Sartison, 2018; FAO, 2019b; Mbow et al., 2019; Chapter 6). UPA cannot fully feed urban dwellers within its boundaries but can make an important contribution to local food security and nutrition (medium confidence) (Martellozzo et al., 2014; Badami and Ramankutty, 2015; Algert et al., 2016; Mohareb et al., 2017; Clinton et al., 2018; Kriewald et al., 2019). UPA is also expected to play important roles in ecosystem functions in addition to alleviating food shocks caused by natural disasters and reducing food mileage. | ||
Categoriesand Description | Synergies | Trade-offs |
Ground-based Unconditioned |
|
|
Traditional, peri-urban field farms, market gardens, community farms, community gardens, home gardens. | ||
Building-integrated Unconditioned | ||
Rooftop gardens, balcony agriculture, and green wall, but production quantity is small. | ||
Ground-based Conditioned |
|
|
Horticultural farms using glasshouses or polyhouses. Often exist on the city fringes. Aquaponics that grow fish in aquaculture systems and re-use nutrient-rich wastewater. One of the few options that provide proteins in urban farms. | ||
Building integrated Conditioned | ||
Rooftop glasshouses, fully indoor, artificially lit plant factories. Recent advancements include production using vertical stacks to produce more food per land area. Indoor aquaculture is also included. | ||
Many UPA farmers are migrant workers or other socially marginalised racial and ethnic groups and often limited by access to land (Lawanson et al., 2014; Horst et al., 2017). There is high agreement that proactive policies for urban design accounting for food–energy nexus and social inclusion including addressing questions of governance and rights to green urban spaces are necessary to enhance food provisioning and to gain multiple functions of UPA (Lwasa et al., 2014; Horst et al., 2017; Mohareb et al., 2017; Siegner et al., 2018; O’Sullivan et al., 2019; Titz and Chiotha, 2019; Halvey et al., 2020).
5.12.6 Changing Dietary Patterns
Dietary change in regions with excess consumption of calories and animal-sourced foods to a higher share of plant-based foods with greater dietary diversity and reduced consumption of animal-sourced foods and unhealthy foods (as defined by scientific panels such as EAT-Lancet) has both mitigation and adaptation benefits along with reduced mortality from diet related non-communicable diseases, health, biodiversity and other environmental co-benefits (high confidence) (Springmann et al., 2016; Springmann et al., 2018; Branca et al., 2019; Henry et al., 2019; Searchinger et al., 2019; Swinburn et al., 2019; Willett et al., 2019; Rosenzweig et al., 2020; Chapter 7.4.2.1.3 and WGIII Chapter 12). Reducing food waste, especially of environment- and climate-costly foods would further extend these benefits (Rosenzweig et al., 2020 and see Section 5.11).
Dietary behaviour is complex: shaped by the broader food system (HLPE, 2017a), the food environment (Herforth and Ahmed, 2015; Turner et al., 2018) and socio-cultural factors (Fischler, 1988). Since most food-related decisions are made at a subconscious level (Marteau et al., 2012), achieving dietary change for personal health reasons has proven difficult; it seems unlikely that dietary change for climate will be achieved without careful attention to the factors that shape dietary choice and behaviour. Food environments, defined as ‘the physical, economic, political and socio-cultural context in which consumers engage with the food system to make their decisions about acquiring, preparing and consuming food’ (HLPE, 2017a): 28), include food availability, accessibility, price/ affordability, food characteristics, desirability, convenience and marketing.
There are a range of options to change dietary patterns, but more research is needed in this area, adjusted to the regional, socioeconomic and cultural context. Studies of policy instruments to change diets include changes in subsidies, taxes, marketing regulation and efforts to change the retail physical environment. Subsidies directed at staple foods and animal sourced foods could be shifted towards diversified production of plant-based foods in order to change the relative price of foods and, thus, dietary choice (Franck et al., 2013; Harris et al., 2021). Taxes on animal-sourced foods that are climate-costly and unhealthy, as defined by scientific panels such as the EAT-Lancet report, could similarly impact relative price (Mbow et al., 2019; Willett et al., 2019). Regulation of marketing could change desirability of climate-unfriendly and unhealthy foods (Willett et al., 2019). Many of the same strategies used to increase sales by conventional food marketing efforts hold potential to change the desirability and people’s preferences for plant foods which are strongly shaped by social–cultural norms. Studies have shown that changes to the number, placing or prevalence of vegetarian options on a menu (Bacon and Krpan, 2018; Kurz, 2018; Garnett et al., 2019; Gravert and Kurz, 2019), the relative price of vegetarian options (Garnett et al., 2021) and the ‘access’ (order and distance) to vegetarian options in the retail physical environment (Garnett et al., 2020) can all increase consumption of plant-based foods and decrease meat consumption (Bianchi et al., 2018). Studies on food environment ‘nudging’ methods found that making the vegetarian meal option the default during conference registration or on a meal plan significantly reduced meat consumption (Campbell-Arvai et al., 2012; Hansen et al., 2019b). Studies simply educating people about the negative health and environmental/climate outcomes of meat consumption have been found to have very little impact (Byerly et al., 2018). More research is needed to understand the potential for motivational crowding in shaping pro-climate dietary choice, as has been demonstrated in development (Agrawal et al., 2015) and conservation interventions (Rode et al., 2015).
5.12.7 Integrated Multisectoral Food Security and Nutrition Adaptation Options
Integrated multi-sectoral strategies that incorporate social protection are effective adaptation responses (high confidence) (Gros et al., 2019; Ulrichs et al., 2019; Medina Hidalgo et al., 2020; Daron et al., 2021; Ilboudo Nébié et al., 2021; Verschuur et al., 2021; 7.4.2, Cross-Chapter Box-GENDER in Chapter 18). Social protection programmes, such as cash transfers, weather index insurance and asset-building activities such as well construction, can support short-term responses to acute food insecurity in response to extreme events but can also build adaptive capacity longer term (Table 5.16, Costella et al., 2017; Ulrichs et al., 2019). An assessment of an adaptive social protection programme in the Sahel found that tailored seasonal forecasting can improve responsiveness to climate-related extreme events, but investment in capacity building and dialogue between forecasters, community groups and humanitarian organisations is needed (Daron et al., 2021). Forecast-based financing, which automatically disperses funds when threshold forecasts are reached for an extreme event (Coughlan de Perez et al., 2016), used in Bangladesh prior to a 2017 flood event allowed low-income, flood-prone communities to access better-quality food in the short term without accruing debt (Gros et al., 2019).
Differentiated responses based on food security level and climate risk can be effective. A study of drought impacts on food security in Senegal between 1997 and 2016 recommended different adaptation strategies based on whether the region was a higher risk of acute short-term food insecurity and/or faced higher risk of drought (Table 5.16; Ilboudo Nébié et al., 2021). Given identified linkages between higher temperatures and extreme events with declines in child dietary diversity, safeguarding diverse diets is one important adaptation priority (Niles et al., 2021). Humanitarian responses are appropriate for short-term acute hunger, while in the medium term, home-grown school feeding programmes with diverse foods can support child nutrition and learning, and with local procurement can also increase income and food security of smallholder farmers (Ilboudo Nébié et al., 2021). Farmer associations can manage regional staple food storehouses, in which farmers store their harvest and receive credit, and can sell their harvest later in the season and pay back the credit with interest, strengthening local supplies and farmer income (Ilboudo Nébié et al., 2021).
A study in Lesotho examined the extent to which climate change increased the likelihood of an acute drought in 2007, and a related food crisis (Verschuur et al., 2021). Given land degradation, reliance on rainfed agriculture and food imports from neighbouring South Africa, the study recommended crop diversification, increased use of drought tolerant crop varieties and expanded trade partners in the medium to long term, to both strengthen regional food production and reduce risk of crop failure and the likelihood of climate-induced drought from trade partners reducing food imports (Verschuur et al., 2021). A longitudinal study of smallholder coffee farmers in Nicaragua found that crop diversification, alongside crop management and varietal improvement, would help farmers strengthen food security long term in the face of climate hazards such as drought and coffee leaf rust (Bacon et al., 2021). Another medium- to long-term adaptation response is to address systemic gender, land tenure and other social inequities as part of an inclusive approach (Bezner Kerr et al., 2019; Khatri-Chhetri et al., 2020; Bacon et al., 2021). This long-term strategy could be part of a human-rights-based approach (HRBA, 5.12.8)
5.12.8 Incorporating Human Rights-Based Approaches into Food Systems
A human rights-based approach (HRBA), endorsed by the UN, is one strategy for addressing core inequities that are key drivers for food insecurity and malnutrition of particular groups such as low-income consumers, children, women, small-scale producers and different regions of the world (FAO, 2013; Claeys and Delgado Pugley, 2017; Caron et al., 2018; Le Mouël et al., 2018; Springmann et al., 2018; Tramel, 2018; HLPE, 2019; Willett et al., 2019). Climate change impacts, mitigation and adaptation approaches can also worsen inequities (Eastin, 2018; Borras et al., 2020). HRBA includes core principles of participation, accountability, non-discrimination, transparency, human rights, empowerment and rule of law, which can be integrated into policymaking and implementation as part of transforming the food system (FAO, 2013; Caron et al., 2018; Toussaint and Martínez Blanco, 2020). The right to well-being can serve as the overarching umbrella of HRBA to addressing climate change within food systems and includes a right to health, right to food, cultural rights, the rights of the child and the right to healthy environment (Swinburn et al., 2019). An HRBA has a specific focus on those groups who are vulnerable due to poverty, discrimination and historical inequities and involves meaningful participation of vulnerable groups in governance, design and implementation of adaptation and mitigation strategies, including gender responsiveness and integration of Indigenous Peoples’ knowledge (UNHRC 2017; Caron et al., 2018; Mills, 2018). There can be conflicts and trade-offs, such as between addressing land rights or traditional fishing grounds, the right to food, and addressing climate justice concerns (Mills, 2018; Borras et al., 2020; Section 5.13). Adaptation strategies that incorporate HRBA include legislation, programmes that address gender inequities in agriculture, agroecology, recognition of rights to land, fishing areas and other natural resources, protection of culturally significant seeds, and community-based adaptation that explicitly involves marginalised groups in governance (Mills, 2018; Tramel, 2018; Huyer et al., 2019; Borras et al., 2020; Section 5.1 4).
5.13 Climate-Change-Triggered Competition, Trade-offs and Nexus Interactions in Land and Ocean
This section presents information about the impacts generated by competition and trade-offs in food systems and discusses opportunities and challenges associated with the use of the Nexus framework.
5.13.1 Impacts of Global Land Deals on Land Use, Vulnerable Groups and Adaptation to Climate Change
Land deals, also known as large-scale land acquisitions (LSLAs), describe recent changes in access to land globally (Borras et al., 2011). Since 2000, at least 160 million hectares have been under negotiation (Land Matrix, 2021). Land deals surged after the 2007–2008 food price crisis and farmland investment boom (Fairbairn, 2014), with a diverse range of drivers (Arezki et al., 2015; Zoomers and Otsuki, 2017; Conigliani et al., 2018) including land-based climate change interventions (Dunlap and Fairhead, 2014; Davis et al., 2015a; Hunsberger et al., 2017; Franco and Borras, 2019). Examples are the expansion of biofuel crops (e.g., Yengoh and Armah, 2016; Aha and Ayitey, 2017), afforestation and reforestation (A/R) projects (Olwig et al., 2016; Richards and Lyons, 2016; Scheidel and Work, 2018), REDD+ (Bayrak and Marafa, 2016; Ingalls et al., 2018), conservation areas (Lunstrum, 2016; Schleicher et al., 2019), renewable energy installations (e.g., Sovacool, 2021) or natural disaster management (e.g. Uson, 2017).
Land deals raise important social justice questions (Franco et al., 2017; Hunsberger et al., 2017; Borras and Franco, 2018b; Borras et al., 2020; Sekine, 2021) (high confidence). Specific impacts of land deals vary according to their purpose, location, actors, land use history and procedural aspects. However, multi-case analyses identify severe adverse impacts (Table 5.18). LSLAs are a significant driver of tropical forest loss (Davis et al., 2020), increasing emissions through deforestation (Liao et al., 2021) and industrialisation of agriculture (Rosa et al., 2021). LSLAs entail large water appropriations (Breu et al., 2016; Chiarelli et al., 2016; Adams et al., 2019), affecting local populations’ access to water and food security (Dell’Angelo et al., 2018; Veldwisch et al., 2018). By increasing exported crops, and limiting local populations’ access to land, LSLAs produce food security risks (Marselis et al., 2017; Müller et al., 2021b). Negative livelihoods impacts arise through enclosure of assets, elite capture (Oberlack et al., 2016), crowding out of small farmers (Nolte and Ostermeier, 2017) and reducing local populations’ access to commons (Dell’Angelo et al., 2016; Giger et al., 2019). Indigenous People are affected, facing high levels of violence in land acquisition conflicts (Dell’Angelo et al., 2021). The social burdens of land deals tend to be gendered (e.g., Fonjong et al., 2016; Nyantakyi-Frimpong and Bezner Kerr, 2017; Atuoye et al., 2021).
Table 5.18 | Adverse social and ecological risks and impacts of agricultural land deals on land use and vulnerable groups.
Land use dimensions | Impacts and implications | References (2014 to present) |
Forestry | Direct and indirect land use change provoked by LSLAs accelerates deforestation of tropical forests globally (medium confidence). | Multi-case analyses Case study examples |
Energy use and access | Expected land use changes provoked by agricultural LSLAs have high fossil-energy footprints. LSLAs may adversely affect local populations’ access to energy resources (medium confidence). | Multi-case analyses |
Carbon emissions | LSLAs have high carbon footprints resulting from deforestation and industrialisation of agriculture (medium confidence). | Multi-case analyses Case study examples Johansson et al. (2020) |
Water use and access | LSLAs frequently involve water appropriations, which may affect access to water, traditional agriculture and the human right to food of local populations (medium confidence). | Multi-case analyses Dell’Angelo et al. (2018) Case study examples |
Food security and nutrition | LSLAs pose food security risks by re-orienting crop production to nutrient-poor crops predominantly destined for export, and/or excluding local populations from agricultural land (high confidence). | Multi-case analyses Cristina Rulli and D’Odorico (2014) Mechiche-Alami et al. (2021) Conceptual studies Case study examples |
Livelihoods | LSLAs often lead to adverse livelihood impacts and increased livelihood vulnerability of local populations (high confidence). | Multi-case analyses Davis et al. (2014) Conceptual studies Case study examples |
Indigenous People and commons | LSLAs often have adverse impacts on Indigenous peoples and lands, including land encroachment, dispossession, and displacement. Land deals frequently target common land and may increase the vulnerability of customary, traditional, and Indigenous systems common property, while reducing their adaptive capacity (high confidence). | Multi-case analyses Conceptual studies Haller et al. (2020) Case study examples Haller (2019) Hak et al. (2018) |
Gender | Impacts and implications of land deals are sometimes experienced in different ways by different genders (high confidence). | Case study examples Yengoh et al. (2015) Nyantakyi-Frimpong and Bezner Kerr (2017) Porsani et al. (2019) |
Impacts on other climate change mitigation and adaptation initiatives | LSLAs may undermine mitigation and adaptation initiatives and other land uses relevant for climate change mitigation and adaptation (high confidence). | Multi-case analyses Case study examples Borras et al. (2020) |
Other environmental impacts | LSLAs are projected to provoke global environmental change; LSLAs are a potential driver of slope instability; LSLAs affect natural habitats such as tiger landscapes; LSLAs jeopardize biodiversity (low confidence). | Chiarelli et al.(2021) Debonne et al.(2019) |
Local populations can experience declining access to livelihood resources and deteriorating food security, increasing gendered vulnerabilities (Yengoh et al., 2015; Faye and Ribot, 2017; Atuoye et al., 2021). Vulnerable groups displaced by land deals may face higher exposure to climate change (Dell’Angelo et al., 2017). LSLAs affecting common-pool resources governed by Indigenous institutions jeopardise the resilience and adaptive capacity of local socio-ecological systems (Dell’Angelo et al., 2016; D’Odorico et al., 2017; Hak et al., 2018; Haller, 2019; Haller et al., 2020). Growing land tenure insecurity may force farmers to engage in unsustainable farming and forestry practices (Aha and Ayitey, 2017; Gabay and Alam, 2017) and hinder agroecological innovations to manage climate risks (Nyantakyi-Frimpong, 2020b). Social justice concerns and vulnerability of local populations can be addressed by promoting land redistribution and recognition, particularly for customary lands of Indigenous and ethnic minorities, and land restitution to those who were forcibly displaced (Franco et al., 2015; Borras and Franco, 2018a).
5.13.2 Trade-offs Generated by Agricultural Intensification and Expansion
Agricultural intensification seeks to increase agricultural productivity per input unit, reducing the pressure on land use and generating positive impacts in GHG emissions (Mbow et al., 2019), but valuing the final effect requires common metrics in terms of carbon capture or emission reductions (Searchinger et al., 2018). It has been suggested to address multiple SDGs (SDG2, SDG13, SDG15) but only occasionally leads to simultaneous positive ecosystem service and well-being outcomes (Rasmussen et al., 2018). When the process relies only on increasing input use, there is a risk of generating adverse outcomes that may override positive effects, such as CO2 emissions, (McGill et al., 2018), NOx emissions (Hickman et al., 2017), soil salinisation and groundwater depletion (Doody et al., 2015; Daliakopoulos et al., 2016; Fragaszy and Closas, 2016; Foster et al., 2018; Flörke et al., 2019). Agricultural intensification could meet short-term food security and livelihood goals, but reduces biological and landscape diversity, and ecosystem services (high confidence) (Campbell et al., 2017; Balmford et al., 2018; Springmann et al., 2018; Ickowitz et al., 2019; Mbow et al., 2019). Agricultural intensification can also affect livelihoods of small-scale producers, compromising food security. It can increase low-waged casual farm work, increasing gender and income inequity (Bigler et al., 2017; Clay and King, 2019; Table 5.18).
Land available for provisioning ecosystem services is declining in many places because of agricultural expansion, bioenergy crops and reforestation for mitigation (Kongsager, 2018), with adverse climate impacts (Froese and Schilling, 2019). Cropland expansion can deteriorate biodiversity (Delzeit et al., 2017), water quality (Ayala et al., 2016) and carbon storage (Goldstein et al., 2012) and increase water demands (Yokohata et al., 2020).
A systems-based perspective on land use is needed to address climate change impacts on nutrition security and ecosystem services (Springmann et al., 2018; IPCC, 2019b; Willett et al., 2019). Land sparing sets aside some land for conservation purposes and intensifies production on farmland (Balmford et al., 2018; Benton et al., 2018; IPCC, 2019b), with potential to offset GHG emissions (Lamb et al., 2016). Alternatively, ‘land sharing’ approach employs principles such as minimising fossil-fuel-based inputs, maximising synergies, and addressing climate change mitigation and adaptation as well as biodiversity (Kremen and Miles, 2012; Kremen, 2015; Kremen and Merenlender, 2018; HLPE, 2019; Section 5.1 4, Box on Agroecology). Community-managed initiatives can address biodiversity and ecosystem conservation, livelihoods, food provisioning and other ecosystem services (Kremen and Merenlender, 2018; HLPE, 2019).
The concept of sustainable intensification has emerged, looking for enhancements in environmental outcomes, while maintaining or increasing agricultural systems performance. There is a potential to find synergies between agricultural production and landscape systems if systems are designed to operate within planetary boundaries (Rockström et al., 2017; Liao and Brown, 2018; Pretty, 2018; Pretty et al., 2018).
5.13.3 Competition between Food Systems in Land and Ocean
Livestock and aquaculture feeds utilise crops such as soyabean and maize, with food conversion efficiencies similar in chicken and Atlantic salmon, and higher in pigs and cattle (Troell et al., 2014; Fry et al., 2018b; Fry et al., 2018a). Use of wild fish meal and oil has been decreasing, partly due to concerns regarding vulnerable small pelagic fish stocks (Bindoff et al., 2019). The instability of wild fish stocks has increased terrestrial crop feed components (Troell et al., 2014; Blanchard et al., 2017; FAO, 2017; Cottrell et al., 2018). The use of wild fish in fish feeds that may have been directly consumed may put low-income households at risk of food insecurity (Troell et al., 2014). An increasing demand for aquaculture products intensifies competition for feed supplies (medium confidence) (Troell et al., 2014; Blanchard et al., 2017). Increases in demands for animal protein and shifts to pescatarian diets will increase the existing competition for land resources, particularly in low- and medium-income countries, with negative impacts on food security (Makkar, 2018), but may be mitigated by dietary changes, novel feeds and food waste usage for aquatic systems (Berners-Lee et al., 2018; Hua et al., 2019; Cottrell et al., 2020).
Competition over use of major aquaculture feed crops (Fry et al., 2016) with terrestrial livestock (Troell et al., 2014), and fish use by terrestrial livestock, will also place pressure on fish and crop resources (medium confidence) (Cottrell et al., 2018). Increases in feed prices will affect fish and meat prices (Troell et al., 2014), and changes in agriculture will be needed to satisfy aquaculture demands (Blanchard et al., 2017). Aquaculture and livestock dietary components may also compromise crops and forage fish that provide essential nutrients for low-income households increasing nutritional insecurity, in regions of sub-Saharan Africa, Asia and Latin America (Troell et al., 2014). Waste fish products can supplement fish meal and oil to reduce competition for feed, as well as reducing use of fish that could go to human consumption (medium confidence) (Little et al., 2016; Shepherd et al., 2017; Dave and Routray, 2018; Naylor et al., 2021). Use of algae, bacteria, yeast and insect diets could replace fishmeal for aquaculture (Cohen et al., 2018; Hua et al., 2019; Cottrell et al., 2020), not affecting nutritional profiles (Campanaro et al., 2019), and fish could be reared on waste by-products of other food production systems (Bava et al., 2019). Complete fish oil substitutions with microalgae may be possible without compromising omega-3 contents, but energy usage in diet production should be considered Cottrell et al. (2020). Substitutions of plant-based and alternative feeds may decrease food conversion efficiencies (Cottrell et al., 2020), affect omega-3 content of farmed seafood (Fry et al., 2016; Shepherd et al., 2017), be problematic for the fish themselves (Little et al., 2016; Naylor et al., 2021) and lead to reduced productivity (Shepherd et al., 2017).
Competition will be heightened by other climate impacts, such as changes in water availability. Water usage is relatively high in animal production (Abraham et al., 2014; Sultana et al., 2014; de Miguel et al., 2015; Palhares and Pezzopane, 2015; Weindl et al., 2017). In some areas, increased demand for plant-based animal feeds will be affected by sea level rise and competing usage of available freshwater with other users, and ecosystem needs (Karttunen et al., 2017).
5.13.3.1 Agricultural and river runoff
Flooding on agricultural land will enhance nutrient runoff, creating eutrophication and increasing harmful phytoplankton blooms, affecting fisheries and aquaculture, human health and ecosystem biodiversity. Changes in precipitation, monsoons, runoff and flood potential combine with deforestation and poor sewage treatment, resulting in larger volumes of nutrients and freshwater reaching coastal ecosystems (Jin et al., 2018; Nasonova et al., 2018; Tamm et al., 2018). Rising surface temperatures, ocean acidification and eutrophication will increase pathogenic Vibrio bacterial loads in marine organisms, with potential transfer to humans (Hernroth and Baden, 2018). Shallow and microtidal estuaries will be more vulnerable to changing river runoffs and saltwater intrusions, eutrophication and hypoxia (high confidence) (IPCC, 2019c).
5.13.4 Maladaptation Responses and Sustainable Solutions
Maladaptation can result in three types of outcomes (Juhola et al., 2016): (1)rebounding vulnerability: short term adaptations that decrease adaptive capacity and hinder future choices; (2)shifting vulnerability: larger-scale adaptation actions that produce spill-over effects in other locations; (3)eroding sustainable development : adaptation strategies which increase emissions and deteriorate environmental conditions and/or social and economic values (Tables 5.20 and 5.21).
Existing climate policies do not adequately consider trade-offs, adaptive limits, cumulative costs and potential risks of maladaptation (robust evidence, medium agreement ) (Dovie, 2017; Holsman et al., 2019; IPCC, 2019b; Work et al., 2019; Thomas, 2020: Table 5.19). Government policies are seldom coordinated across scales and often focused on regional short-term risks (medium evidence, medium agreement ) (Dovie, 2017; Holsman et al., 2019; Rahman and Hickey, 2019; Butler et al., 2020). Past development trajectories and dominant political economic structures may narrow adaptation pathways, be restrictive and increase the vulnerability of particular groups (Paprocki, 2018; Quan et al., 2019; Rahman and Hickey, 2019; Work et al., 2019).
Table 5.19 | Case studies of trade-offs and negative outcomes associated with agricultural intensification on biodiversity and ecosystem services.
Ecosystem service | Trade-offs/negative outcomes | References |
Provisioning: water quality | Negative impacts on ephemeral wetlands | Dalu et al. (2017) |
Provisioning: water availability | Contribution to water scarcity | |
Supporting: soil | Increasing erosion risk | |
Regulating: climate | Reduced SOC sequestration | |
Regulating: pest control | Reduced level of biological control of pests; reduced number of insectivorous birds | |
Cultural: recreational | Reduction of river wildlife | |
Biodiversity | Reduced global biodiversity | Newbold et al. (2015), Egli et al. (2018), Beckmann et al. (2019) |
Biodiversity | Reduction of taxonomic diversity | Jeliazkov et al., (2016), Kehoe et al. (2017), Banerjee et al. (2019) |
Biodiversity | Negative impacts on mean population stability |
Case Studies of Maladaptation
Large-Scale Irrigation Project in Navarre, Spain
Many small-scale producers could not afford the irrigation investment and had to sell or rent their land to those who joined the irrigation project. Many large-scale farmers using irrigation switched to corn and forage and dropped crops with high labour costs. Water costs are now paid to a private company, and small-scale farmers lost access to communal water rights. The project increased inequity and land concentration and lowered crop diversity, with small-scale producers more vulnerable to climate change. Large-scale intensive farmers are more exposed to crop price volatility than to climate vulnerability but have greater access to subsidies and water rights (Albizua et al., 2019).
Constraining Adaptation: Previous Agricultural Development Pathways in India
Government policies in colonial and postcolonial India invested in infrastructure, export production and synthetic input use (Gupta, 1998; Davis, 2001), setting the stage for current development trajectories and closing out other adaptive options. Although such policies increased national food production, they failed to address high levels of malnutrition, worsening regional inequities, degraded natural resources and an agrarian debt crisis (Singh, 2000; Gupta et al., 2016; Gajjar et al., 2019). Agricultural livelihoods are increasingly considered unviable, with lower adaptive capacity of farmers, high debt levels (Gupta et al., 2016), and Indigenous and local knowledge loss and denigration (Kumar, 2016) alongside lower crop diversification (Srivastava et al., 2016). Government institutions aimed at infrastructure often lack adaptive capacity needed to address rural livelihoods (Singh et al., 2017; Gajjar et al., 2019).
Adaptation options that consider adverse effects for different groups reduce the risk increasing vulnerability, negatively affecting socioeconomic factors to deal with climate impacts, or impeding efforts to implement SDGs (high confidence) (Juhola et al., 2016; Antwi-Agyei et al., 2018; Paprocki and Huq, 2018; Holsman et al., 2019; IPCC, 2019b; Stringer et al., 2020). Adaptation methods considering historical roots of current vulnerabilities can identify viable solutions, which are difficult to undertake because of path dependencies (high confidence) (Ribot, 2014; Albizua et al., 2019; Gajjar et al., 2019; Paprocki, 2019; Thomas, 2020). Planning techniques that model outcomes for different groups from different adaptation options could be put in place to diminish maladaptation risks (Rodríguez et al., 2019).
Inclusive planning initiatives such as community-based anticipatory adaptation combined with ‘two-way learning’ that considers future scenarios and different adaptation pathways can prevent maladaptation (high confidence) (Dovie, 2017; Bezner Kerr et al., 2019; Neset et al., 2019a; Rahman and Hickey, 2019; Work et al., 2019; Butler et al., 2020; Nunn et al., 2020; Piggott-McKellar et al., 2020; Westoby et al., 2020; Table 5.20). Promising policy management tools combine temporal scales and mitigation–adaptation interactions and consider political dynamics, socioeconomic impacts and trade-offs for vulnerable groups, long-term support for policy leaders, efforts to establish livelihood ‘niches’ and ongoing participatory evaluation (Dovie, 2017; Holsman et al., 2019; Rahman and Hickey, 2019; Work et al., 2019; Butler et al., 2020). A focus on the most disadvantaged groups can help small-scale producers at higher risk to prevent maladaptation (FAO, 2018c). Governance mechanisms have emerged that consider food security, socio-cultural factors, and land and water rights, using participatory, inclusive ‘two-way learning’ methods that involve vulnerable people alongside government (IPCC, 2018; Holsman et al., 2019; IPCC, 2019b; Rahman and Hickey, 2019; Butler et al., 2020).
Table 5.20 | Summary of the emerging literature on potential risks of maladaptation.
Description of adaptationstrategy | Potential negative impacts | Maladaptation typology (1 = rebounding vulnerability, 2 = shifting, 3 = eroding SDGs) and confidence level | Regions andcountriesaffected | Groups affected | References |
Agricultural intensification to increase productivity, in places with heavy rainfall events or rising pest/disease incidence | Increases GHG emissions, water pollution, possible insect resistance and costs to farmers; possibly increases inequities. May constrain adaptation policy options for development pathways due to lock-ins and trade-offs which entrench inequities. | 1, 2, 3 Robust evidence, medium agreement | USA, Africa, Asia (India, China), Europe | Farmers, pastoralists/nearby communities who rely on water; small-scale farmers who cannot afford inputs; policymakers. | Gajjar et al. (2019), Guodaar et al. (2019), Houser and Stuart (2019), Neset et al. (2019b), Quan et al. (2019), Young and Ismail (2019) |
Livelihood diversification into charcoal production | Increases GHG emissions and deforestation rates | 1, 3 medium | Africa (Northern Ghana), South America (Peru) | Small-scale food producers; Indigenous communities | Antwi-Agyei et al. (2018), Zavaleta et al. (2018), Young and Ismail (2019) |
Irrigation projects or programmes that are either large-scale and/or rely on groundwater | Reduces long-term potential for hydropower and groundwater availability, can increase salinisation and cost of water. Can increase cost of farming and debt levels of farmers, squeezing out small-scale producers. Can reduce water availability for aquaculture. | 1, 2, 3 high | Central China, India, Mediterranean areas, Europe, USA | Food producers who rely on irrigation; consumers who rely on hydropower or groundwater; small-scale diversified producers who cannot afford irrigation; aquaculture. | Doody et al. (2015), Herbert et al. (2015), Barik et al. (2016), Daliakopoulos et al. (2016) Fragaszy and Closas (2016) Dalin et al. (2017), Foster et al. (2018) Hanaček and Rodríguez-Labajos (2018), Albizua et al. (2019), Flörke et al. (2019) Gajjar et al. (2019), Zhang et al.(2019a) |
Investment in improved cultivars or shift to different crops | May displace local varieties; reduces diversity if too much policy/extension emphasis falls on a few varieties; may increase risk of crop loss from pests, disease or drought if reliant on a few varieties; may increase fertilizer use; may lead to loss of Indigenous or local knowledge. | 1, 3 medium | South America (Bolivia, Pacific Islands, Asia | Small-scale food producers; Indigenous communities | Mcleod et al. (2018), Meldrum et al. (2018), Neset et al. (2019b) Rahman and Hickey (2019) |
Migration | Can increase the workload of people left behind (often women), worsen rural livelihoods and food insecurity; can lead to worsened living conditions, food security and poverty in precarious urban conditions; may increase vulnerability to flooding in urban locations. May affect mental health by disrupting existing social ties. | 1, 3 high | Asia, Africa, Central and South America | Small-scale low-income food producers or rural workers; women | Bettini et al. (2017), Paprocki (2018), Chen et al. (2019), Jacobson et al. (2019), Michael et al. (2019), Young and Ismail (2019), Singh and Basu (2020), Torres and Casey (2017) |
Coastal sea walls, embankments, canals, riverbed draining and dikes to reduce flood risk | Can degrade coastal mangroves, deplete open freshwater fisheries, cause sedimentation of rivers, reduce fish diversity and increase flooding risk for particular vulnerable groups; may divert funds from other more sustainable measures. | 1, 2, 3 high | Asia, South Pacific Islands, West Africa | Coastal communities dependent on mangroves and fisheries; low-income rural households with seasonal dependence on inland fisheries | Dovie (2017), Owusu-Daaku (2018), Freduah et al. (2019), IPCC (2019c), Rahman and Hickey (2019), Nunn et al. (2020) Seddon et al. (2020), Thomas (2020) |
River regulation for hydropower | May have negative impacts on inland fisheries. | 2, 3 | Global | Small-scale inland fisheries and low-income rural households with seasonal dependence on inland fisheries | |
Government policies to manage coastal fisheries which promote overcapitalisation of fisheries, including index insurance | Government confiscation of fishing nets to prevent rapid decline of fish population can worsen livelihoods for small-scale fishers; subsidies of pre-mixed fuel to allow fishers to stay out longer due to shifting fish populations may increase total number of fishers and total fish catch; insurance payments may benefit larger-scale fishing fleets and push out small-scale fishers. | 1, 3 medium | West Africa | Coastal small-scale fishery communities | FAO (2018b), Freduah et al. (2019), Holsman et al. (2019), Sainsbury et al. (2019) |
Consultative stakeholder systems in fisheries or flood management | May encourage inertia in the system due to a few powerful stakeholders participating in the consultative process. | 2 low | North America, Asia | Coastal fisheries | |
Climate services | May reinforce existing inequities if climate services are attuned to powerful stakeholders in industry, services are privatised, there are limited ways to get input from vulnerable groups and planning budgets that use climate services are constrained. | 1, 2, 3 medium | North America | Coastal fisheries, farming | |
Nature-based solutions mitigation and adaptation strategies such as reforestation or afforestation | Can displace local communities’ access to land for food production and other ecosystem services, have negative impacts on Indigenous rights, reduce biodiversity and may not reduce GHG as much as conserving natural forests and wetlands or agroecological systems such as agroforestry or other means to increase soil C. | 2, 3 medium | Africa, Asia, and South America, e.g., Indonesia, Amazon, west-central Africa | Indigenous communities; small-scale producers and forest-dependent communities | Lunstrum et al. (2016), Work et al. (2019), Seddon et al. (2020), Cross-Working Group Box BIOECONOMY this chapter) |
Social safety nets provide funds which increase consumption of processed, purchased food and erode Indigenous knowledge | Decline in Indigenous knowledge of and collective approaches to seasonal adaptation strategies in hunting, fishing and food production; shift in dietary patterns to more processed and non-local foods; reduction in farming. Reduced capacity to respond to hazards through dispersed settlement, e.g., hunting, fishing, wild food collection. Increased population density increases deforestation and vulnerability. | 1, 3 low | South America (Amazonian region of Peru), Africa (South Africa) | Indigenous communities | |
Community-based adaptation strategies | Local gender and other social inequities can lead to ‘elite capture’ that reinforces inequity; power dynamics between the funding agency and local participants can make local community involvement tokenistic. There may be inadequate attention to socio-cultural preferences and structural factors which foster maladaptation such as inappropriate crops or animals used. | 1, 3 high | Pacific Islands, Africa, Asia | Small-scale food producers; Indigenous communities, other vulnerable groups such as women and low-caste groups | McNamara and Buggy (2017) Jamero et al. (2018), Singh (2018) Bezner Kerr et al. (2019) Piggott-McKellar et al. (2020), Westoby et al. (2020) |
Digital agriculture for increased precision and efficient use of fertilizers, pesticides, water | Could lead to net job losses, particularly for those with lower levels of education; increased surveillance and employer scrutiny of lower-skilled workers in fields, greenhouses and processing plants and warehouses; separate workers from employees and companies who collect data. Overall increased racial, income inequities and unequal working conditions. | 2, 3 low | North America, South America, Europe, Asia, parts of Africa | Farmworkers; small-scale food producers who cannot afford digital technologies; rural communities | |
Increased credit access for livelihood diversification | High interest rates, tight return policies could increase debt loads for low-income households, which could rebound vulnerability. Household may invest in livelihood strategies which are vulnerable to climate change impacts, or which increase GHG. | 1, 3 low | Asia (Bangladesh) | Low-income landless people or small-scale producers | Rahman et al. (2018) |
Aquaculture | Large-scale coastal aquaculture can increase soil salinisation and reduce land available for other food production and increase migration. | 2, 3 low | Asia (Bangladesh) | Small-scale mixed systems including rice production and other rural livelihoods |
Table 5.21 | Strategies to avoid maladaptation (adapted from Magnan, 2014; Lim-Camacho et al., 2015; Sovacool et al., 2015; FAO, 2018b; Paprocki and Huq, 2018; Sainsbury et al., 2019).
Type of maladaptation | Strategies |
Environmental |
|
Socio-cultural |
|
Political-economic |
|
5.13.5 Climate Change and Climate Response Impacts on Indigenous People
Indigenous people and ethnic minorities, many of them having special cultural associations to local foods, are particularly vulnerable to climate change owing to changes in the availability of wild foods, crop failure and food production losses or increased food prices (Norton-Smith et al., 2016; Otto et al., 2017).
Changes in sea level rise or coastal erosion can reduce ecosystem services to a point where either subsidies are used to enable human populations to remain in their place of attachment, or ultimately to displace coastal residents, thereby removing connections to places of intrinsic value. For example, the United Houma Nation in Louisiana is experiencing coastal land loss, sea level rise and strong Gulf hurricanes, which leads to the relocation of some tribes causing loss of Houma identity (Sullivan and Rosenberg, 2018). Another example is the relocation of Alaska Native communities due to climate change (Hamilton et al., 2016)
Expansion of agriculture can bring distress to Indigenous communities because of environmental deterioration and the stress associated with relocation or displacement (Otto et al., 2017). A/R programmes can also bring inequities to Indigenous communities (Godden and Tehan, 2016) and even violent displacement with tragic results (Celentano et al., 2017). A/R programmes can negatively affect a range of substantial and procedural Indigenous Peoples’ rights entrenched in international human rights law (Table 5.22) and their potential for climate change adaptation (high confidence).
Table 5.22 | Indigenous rights recognised in international human rights law negatively affected by A/R projects.
Negative impacts of monoculture plantations (and other A/R projects) | Indigenous Peoples’ rights affected | Degree of certainty | References |
Local community not informed, not adequately consulted, not provided means for meaningful participation in project design, implementation, and monitoring (with specific attention to women and poor households); disruption or non-recognition of local or traditional institutions; elite capture; no access to third-party grievance mechanisms. | Right to self-determination; consultation and free, prior and informed consent (FPIC); participation | Medium evidence, high agreement | Aggarwal (2014), Maraseni et al. (2014), Ravikumar et al. (2015), Bayrak and Marafa (2016), Loaiza et al. (2016), Vijge et al. (2016), Pye et al. (2017), Ryngaert (2017), Wolde et al. (2016), Brancalion and Chazdon (2017), Seddon et al. (2020) |
Evictions and displacement; dispossession; livelihood precarity; and criminalisation of forest-dwelling people. | Right not to be forcibly removed | Medium evidence, high agreement | Mingorría (2014), Richards and Lyons (2016), Witasari (2016), Corbera et al. (2017), Pye et al. (2017), Sarmiento Barletti et al. (2020), Brancalion and Chazdon (2017) |
Loss, transfer or acquisition of land. A/R projects involve changes in land use for medium to long term and often lack consideration for local dynamics including land tenure and competition with agriculture or conservation. | Rights to land and territory | Limited evidence, high agreement | Aggarwal (2014), Robinson et al. (2014), Bayrak and Marafa (2016), Pye et al. (2017), Bond et al. (2019) |
A/R projects exacerbate conflicts, accentuate uneven power relations, increase existing inequities within communities, exclude the poor and deepen structural injustices, including racism and stigmatisation. | Rights to land and territory | Limited evidence, low agreement | |
Forest expansion intensifies already acute land shortages for growing food and forces villagers to take their animals for grazing to new areas as a result of forests being fenced off. | Rights to land and territory (with implications for food security) | Limited evidence, high agreement | Lyons et al. (2014), Wolde et al. (2016), Brancalion and Chazdon (2017), Mousseau and Teare (2019) |
Decreased stream flows and water yields; exacerbated water scarcity. | Right to water | Robust evidence, high agreement | Veldman et al. (2015), Aitken and Bemmels (2016), Brancalion and Chazdon (2017), Pye et al. (2017), Bond et al. (2019), Seddon et al. (2020) |
Pollution of lakes with agrochemicals; heavy chemical use, including the spread of pesticides, herbicides and fertilizers by aircraft and other means causing runoff into rivers. | Right to a healthy environment | Medium evidence, high agreement | Richards and Lyons (2016), Johansson and Isgren (20179, Pye et al. (2017) |
Encroachment on other ecosystems with devastating impacts on biodiversity; pressures on ecologically sensitive ecosystems such as wetlands; reduction in seed-dispersing animals; planted tree species becoming invasive, introducing pests and diseases. | Right to a healthy environment, right to food | Medium evidence, high agreement | Richards and Lyons (2016), Holmes et al. (2017), Seddon et al. (2020), Ennos et al. (2019) |
Loss of habitat, degradation of savannas, native grasslands (grassy biomes) or mangroves wrongly characterised as degraded land suitable for afforestation. | Right to a healthy environment, right to food | Robust evidence, high agreement | Veldman et al: (2015), Cormier-Salem and Panfili (2016), Brancalion and Chazdon (2017), Bond et al. (2019), Seddon et al. (2020) |
Direct negative health impacts; loss of traditional medicine. | Right to health | Limited evidence, medium agreement | |
A/R projects affect burial sites as, for many communities, the forest is also the resting place for deceased ancestors. | Right to cultural identity and to main and control their traditional knowledge | Limited evidence, high agreement | Lyons et al. (2014), Gabriel and Mangahas (2017), Mousseau and Teare (2019) |
Loss of traditional or Indigenous ecological knowledge and forest management practices | Right to cultural identity and traditional knowledge | Limited evidence, medium agreement | |
Increased labour burden. Benefit sharing by direct cash transfer or in-kind modalities tends to not compensate lost income opportunities. Some projects bring employment opportunities, but these are short term and limited and rarely viable if the opportunity cost of land and labour is considered. Poor farmers may drop out to regain access to their land for uses that provide cash returns in the shorter term. | Right to an adequate standard of living; right to decent work; right to benefit sharing | Medium evidence, medium agreement | Boyd et al. (2007), Aggarwal (2014), Cagalanan (2016), Witasari (2016), Corbera et al. (2017), Pye et al. (2017) |
A significant proportion of land targeted for A/R projects is inhabited and used by Indigenous Peoples and local communities (Cagalanan, 2016). Indigenous Peoples have rights to and/or manage at least 37.9 million km 2 of land and influence land management across at least 28.1% of the land area (Garnett et al., 2018). At least a quarter of the global land area is traditionally owned, managed, used or occupied by Indigenous Peoples, overlapping with 35–40% of the area that is formally protected (Garnett et al., 2018; Brondizio et al., 2019). In many cases, A/R is implemented in areas where tenure rights are insecure and Indigenous Peoples’ rights are at risk of being disregarded (Naughton-Treves and Wendland, 2014; Kohler and Brondizio, 2017; Garnett et al., 2018) (medium evidence, high agreement ). Many projects are also found in areas where complex socio-political contexts challenge management (Jurjonas and Seekamp, 2019). It is anticipated that A/R projects will create huge pressures on existing land uses and generate further land use conflicts (Aggarwal, 2014; Robinson et al., 2014; Paul et al., 2016; Brancalion and Chazdon, 2017; Pye et al., 2017; Bond et al., 2019). In addition, many afforestation projects are conducted in regions that are not bio-climatically suitable, leading to the degradation of ecosystems that are key to local livelihoods (Veldman et al., 2015; Robinson et al., 2016b).
Until 2010, most A/F projects had technical, carbon-related goals and did not consider issues of livelihoods, community involvement or broader ecosystem impacts (Wolde et al., 2016). New strategies such as nature-based solutions (Seddon et al., 2020) and forest and landscape restoration (Brancalion and Chazdon, 2017) integrate a larger set of social and environmental objectives. Indigenous Peoples enjoy a range of co-benefits of A/F initiatives such as improved habitat, fire management or protection from climatic shocks such as drought (Robinson et al., 2016b; Seddon et al., 2020), provided they are able to manage carbon funds collectively, meet the monitoring and reporting requirements, and protect forests from illicit uses and natural disasters (Wolde et al., 2016).
Policies and safeguards attached to specific A/R initiatives determine their impact (high confidence) (Talor, 2015; West, 2016; Brancalion and Chazdon, 2017). In countries where there is a great level of devolution of rights to Indigenous Peoples, there is a risk that the A/R agenda will lead to recentralisation (limited evidence, medium agreement ) (Bayrak and Marafa, 2016). Some A/R initiatives specify the need to respect the rights of Indigenous Peoples and local communities and protect biodiversity (medium evidence, high agreement ) (Seddon et al., 2020).
Local communities’ ability to participate in project design, implementation and monitoring is directly linked to the autonomy and independence of local institutions (Pye et al., 2017), their ability to formulate by-laws (Wolde et al., 2016) and handle funds in a transparent way (medium evidence, high agreement ) (Witasari, 2016). It is further dependent on cohesion in the community (Cagalanan, 2016), the existence of clear rules delineating community membership and the presence of elders and community members with relevant local knowledge (Robinson et al., 2016b), and gender and out-migration dynamics affecting participation structures (robust evidence, medium agreement ) (Cormier-Salem and Panfili, 2016; Witasari, 2016; Wolde et al., 2016; Jurjonas and Seekamp, 2019).
5.13.6 Increased Presence of Financial Actors in the Agrifood System
Financial actors, markets, institutions and incentives have gained importance in agricultural commodities and farmland markets in the past two decades (Clapp and Isakson, 2018; Fairbairn, 2020). New types of investment vehicles such as commodity index funds that track prices of commodities and farmland have emerged, and the use of older vehicles such as forward and futures markets has increased (Schmidt and Pearson, 2016; Clapp and Isakson, 2018). These trends are connected to climate change as financial investments are influenced by the likelihood that climate change will increase commodity and farmland price variability (medium confidence) (Cotula, 2012; Isakson, 2014; Tadesse et al.).
Financial investors pool their investments through intermediaries, alongside other dynamic forces in the global economy, making unambiguous assessments of their effect difficult (Clapp, 2014; Clapp, 2017). However, assessment of the broader trends at the interface of financial investment, food system dynamics and climate change shows potential connections.
Climate-induced variability in food production has the potential to introduce a new level of uncertainty into food and farmland markets, encouraging financial investment into products to capitalise on price volatility and to hedge risks. The new financial instruments enable investors to speculate more easily on the direction of food and land prices, especially when they are volatile (Ouma, 2014; Baines, 2017).
5.13.7 Climate Change Interactions with other Drivers—Food–Water–Health–Energy–Security Nexus
Linkages between food security and nutrition with water and energy as well as other important socio-environmental issues are increasingly being described within a nexus framework (see also Chapters 3, 4, 6 and 7), with food systems frequently located at the centre of nexus concepts (Caron et al., 2018).
Climate change will affect the food–energy–water (FEW) nexus, commonly in the form of risk multiplier (high confidence) (e.g., Conway et al., 2015; Barik et al., 2016; Keairns et al., 2016; Abbott et al., 2017; Ebhuoma and Simatele, 2017; Caron et al., 2018; D’Odorico et al., 2018; de Amorim et al., 2018; Mpandeli et al., 2018; Nhamo et al., 2018; Soto Golcher and Visseren-Hamakers, 2018; Yang et al., 2018; Amjath-Babu et al., 2019; Froese and Schilling, 2019; Mercure et al., 2019; Momblanch et al., 2019; Pastor et al., 2019; Xu et al., 2019). Xu et al. (2019) modelled the irrigation demand for large-scale maize production in Northeast China and concluded that increasing droughts under future climate change would lead to extreme shortage of irrigation water without adaptive measures. Barik et al. (2016) described how the growing demand for food in India has led to more irrigation with a reduction in groundwater levels in some regions.
Increasing demands for food, energy and water can lead to domestic and international conflict, including political instability and migration, often in the context of drought (high confidence) (Abbott et al., 2017; Bush and Martiniello, 2017; WEF, 2017; D’Odorico et al., 2018; de Amorim et al., 2018). de Amorim et al. (2018) conclude that the FEW nexus is susceptible to many global risks, including extreme weather events and human migrations, and predominantly endangers vulnerable communities of less developed countries. There is emerging evidence that food and water insecurity enhances social conflicts, including protests and violent riots, at least partially, by accelerating existing grievances (Heslin, 2021; Koren et al., 2021). Closer coordination at global, regional and national levels could be recommended to manage these risks.
Meeting growing demands for food, water and energy under a changing climate requires technical solutions and behavioural change as well as greater coordination across multilateral institutions and governance. Supply-side solutions focus on enhancing production, reducing food waste and loss or lowering water demand through both technological approaches (e.g., breeding, improved irrigation) and agroecological approaches, such as agroforestry, underutilised and more adapted crops, and transition towards a circular economy (Alexander et al., 2015; Obersteiner et al., 2016; D’Odorico et al., 2018; Nhamo et al., 2018; Soto Golcher and Visseren-Hamakers, 2018). Demand-side solutions focus primarily on changes in consumer behaviour towards healthier diets with lower carbon footprints, particularly reduction of meat consumption (Alexander et al., 2015; Obersteiner et al., 2016). Improving the coordination of multilateral organisations could result in improved cross-boundary management of natural resources, particularly related to water (Conway et al., 2015; Nhamo et al., 2018; Soto Golcher and Visseren-Hamakers, 2018).
As relationships between individual subsystems are systemic, integrated solutions would result in better outcomes across the FEW nexus (strong agreement ). Obersteiner et al. (2016) concluded that single-sector policies can create strong trade-offs with other policy targets and SDGs, whereas strategies that reduce pressure on food production systems diminish trade-offs between FEW nexus components. This suggests that achieving multiple SDGs will require balancing societal demands in the context of finite natural resources (Jägermeyr et al., 2017; Amjath-Babu et al., 2019; Momblanch et al., 2019).
Despite concluding that integrated solutions addressing the systemic connections between the FEW nexus would improve development and environmental outcomes, there are limitations of integrating multiple frameworks, both in terms of describing the complexities and in finding solutions (Leck et al., 2015; Weitz et al., 2017; Wichelns, 2017; Shannak et al., 2018). Leck et al. (2015) and Weitz et al. (2017) indicate that evidence of successful implementation and improved outcomes based on the application of nexus concepts is rare.
5.14 Implementation Pathways to Adaptation and Co-benefits
5.14.1 State of Adaptation of Food, Feed, Fibre and Other Ecosystem Products
Since AR5, several adaptation reviews have been done (Ford et al., 2015; Lesnikowski et al., 2016). In a review of 1159 peer-reviewed sources, Berrang-Ford et al. (2021b) found that observed adaptations in food, fibre and other ecosystem products have consisted mainly of changes in autonomous behaviour changes, such as changing planting time, followed by technological/infrastructure and ecosystem-based adaptation approaches, the majority of which have occurred in Africa and Asia (Figures 5.20 and 5.21, Table 5.22). Several adaptation options addressed multiple SDGs (e.g., 2, 6, 8, 12) (Figure 5.21).
Figure 5.19 | State of adaptation by region and type of response (based on 1159 peer-reviewed references that addressed adaptation in food, fibre and other ecosystem products sector; source: Global Adaptation Mapping Initiative (GAMI) database (Berrang-Ford et al. , 2021a). The bars indicate the amount of evidence for the category x region.
Assessment of adaptation options was done for 15 potential options for land and ecosystem transitions (SM5.7, Figure 5.22a). Several adaptation options have high to medium feasibility, with robust evidence, high agreement about the adaptive capacity resilience building potential of options in relation to climate change impact drivers (high confidence). Policy and planning and production shifts have limited evidence for feasibility. Most options are technically and physically feasible, with generally high political and social acceptability and environmental feasibility, but have limited evidence for institutional feasibility. Most adaptation options have medium to high microeconomic feasibility (high confidence) but limited evidence for macroeconomic viability.
Figure 5.22 | Assessment of 11 feasibility indicators (six categories), five effectiveness indicators and maladaptation of adaptation options based on 287 peer-reviewed papers. See SM5.7 for methods and data. Scores ranging from 1 (low) to 3 (high) were obtained by averaging five or more papers for each option and indicator. Blank cells were not assessed because of insufficient literature.
Among five effectiveness indicators (SM5.7, Figure 5.22b), most options have robust evidence of reduced risk vulnerability to climate change, with low scores for local governance, substitution of plant or animal type, community forest management, livelihood diversification and climate services. Higher-scored options to reduce risk included increasing biodiversity (at landscape and field level), community seed banks, conventional breeding (plant and animals), mixed systems and agroecological approaches (medium confidence), suggesting multiple co-benefits of these options. Most options have high scores for enhancing social well-being and economic and environmental benefits (medium confidence) but limited evidence for strengthening institutions for most options. There were low scores for potential maladaptation (medium confidence).
Table 5.23 | State of adaptation in food, fibre and other ecosystem products by actors and vulnerabe groups (source: GAMI database; Berrang-Ford et al., 2021a)).
Actors | N (%) | Vulnerable groups | Planned, N (%) | Implemented, N (%) |
International or multi-national governance institutions | 72 (6%) | Women | 134 (12%) | 118 (10%) |
National government | 264 (23%) | Youth | 22 (2%) | 24 (2%) |
Local government | 267 (23%) | Elderly | 31 (3%) | 28 (2%) |
Sub-national government | 89 (8%) | Low income | 201 (17%) | 258 (22%) |
Private sector corporations | 56 (5%) | Disabled | 2 (0%) | 3 (0%) |
Private sector SMEs | 80 (7%) | Migrants | 12 (1%) | 18 (2%) |
Civil Society — international/multi-national/national | 117 (10%) | Indigenous | 95 (8%) | 85 (7%) |
Civil Society — sub-national or local | 257 (22%) | Ethnic minorities | 32 (3%) | 32 (3%) |
Individuals or households | 1087 (94%) |
Figure 5.20 | Observed adaptation across regions in food, fibre and other ecosystem products based on the GAMI database (Berrang-Ford et al. , 2021a). The bars indicate the number of evidence for the options x region.
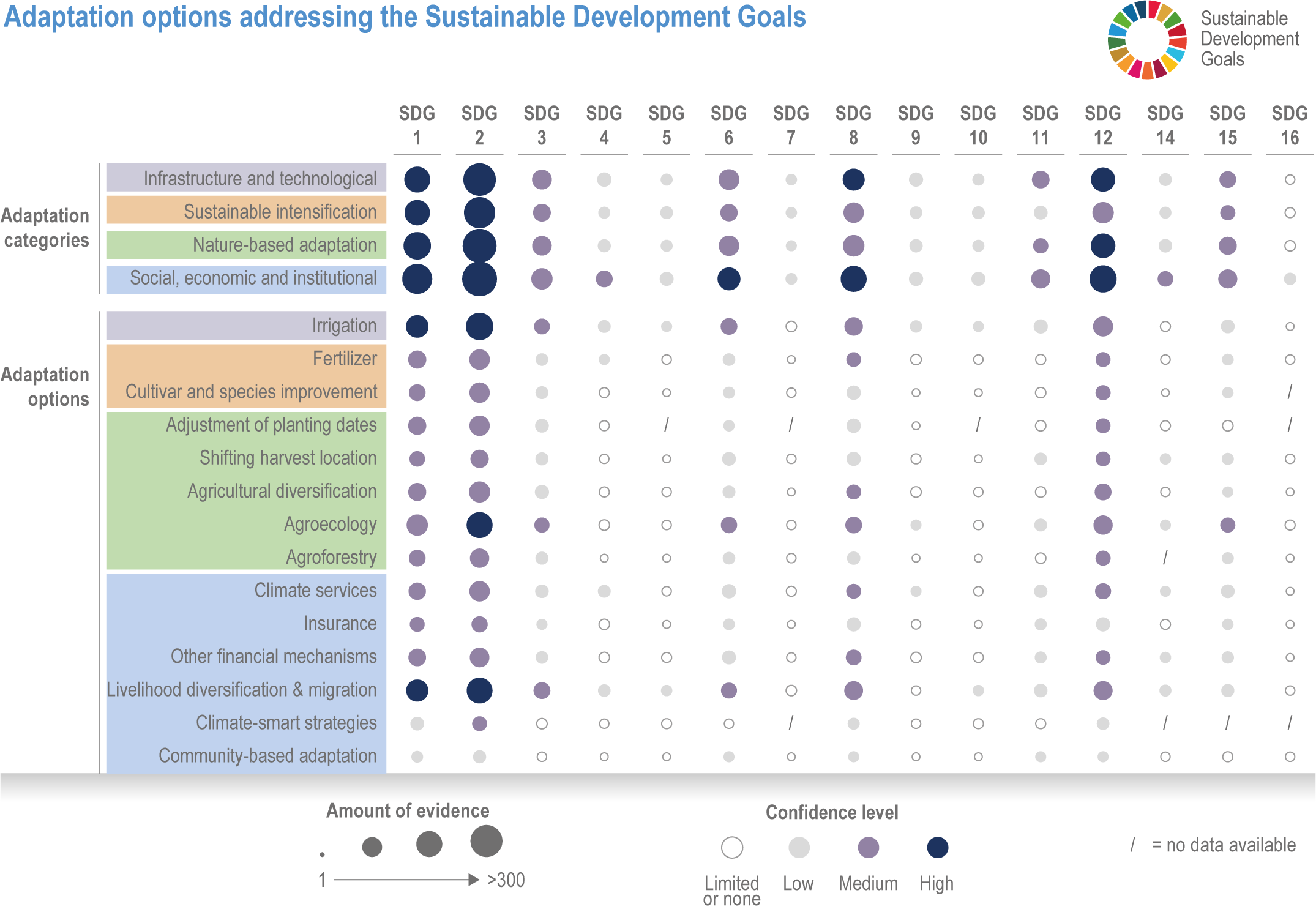
Figure 5.21 | How different response types address the SDGs based on GAMI.
5.14.1.1 Nature-based solutions or ecosystem-based adaptation
There is growing evidence that nature-based solutions (NBS), which emphasise ecological approaches and biodiversity conservation (Chapter 1), have high potential to transform land and aquatic systems into climate-resilient systems (medium evidence, high agreement ) (Albert et al., 2017; Brugère et al., 2019; Galappaththi et al., 2020b; Snapp et al., 2021; Cross-Working Group Box BIOECO; Cross-Chapter Box NATURAL in Chapter 2).
5.14.1.2 Climate services
Climate services, understood as the production, translation, communication and use of climate information in decision-making processes, can contribute to adaptation efforts in agricultural systems (medium agreement , low evidence). Climate services can support decision makers in agriculture by providing tailored information that can inform the implementation of specific adaptation options (Vaughan, 2018; Buontempo et al., 2019; Dobardzic et al., 2019; Hank et al., 2019).
For some high- and medium-income countries, evidence suggests that climate services have been underutilised (Mase and Prokopy, 2014), with limited evidence in these countries of the impact of climate services on yields, income, and food security and nutrition. In low-income countries, use of climate services can increase yields and incomes and promote changes in farmers’ practices (low confidence) (Roudier et al., 2014; Roudier et al., 2016; Tarchiani et al., 2017; Ouedraogo et al., 2018). There is low confidence that climate services are delivering on their potential, whether they are being accessed by the vulnerable, and how these services are contributing to food security and nutrition (Ouedraogo et al., 2018; Vaughan et al., 2019).
Improved design and delivery of climate services can enhance effectiveness (medium confidence). Ways to enhance the impact of climate services include integrating information from multiple sources at different scales (Bouroncle et al., 2019), participatory collection and analysis of climate information (Loboguerrero AM, 2018; Tesfaye et al., 2019; Rossa, 2020), and making forecast information available in local languages and as verbal communications for farmers who cannot read (Nkiaka et al., 2019).
In countries with limited climate data, crowd sourcing (outsourcing data collection to the public) (Minet et al., 2017) and digital tools present an opportunity for addressing climate risk (medium confidence) (Osgood et al., 2018; Thornton, 2018; Partey et al., 2020; Sotelo et al., 2020). Bundling additional services such as market information with climate information may be effective at plugging information gaps (low confidence) (Chatuphale and Armstrong, 2018; Tsan et al., 2019; Tesfaye et al., 2019)
There may be inequality in access to climate services; their use may tend to benefit large-scale operations and disadvantage small- and medium-scale farmers and others who face issues of access due to social and economic inequity; also some groups such as pastoralists have not yet benefitted from climate services (high confidence) (Furman et al., 2014; Muema et al., 2018; Awazi et al., 2019; Nyantakyi-Frimpong, 2019; Paudyal et al., 2019; Vaughan et al., 2019; Nidumolu et al., 2020; Partey et al., 2020). Other challenges include technology ignorance, data privacy and security, data access permissions, software and system compatibility, and understanding how to use and derive value from accessed data (Chatuphale and Armstrong, 2018; Drewry et al., 2019). More work is needed to understand the factors that prevent farmers and fishers from benefitting from this new information. Recent assessments suggest that access to, and value of, climate and weather information can be enhanced by the development of digital tools (including radio, text messages, etc.) appropriate to the specific needs of different vulnerable groups, as well as by including these groups in their development and building their capacity (medium confidence) (Camacho and Conover, 2019; Gumucio et al., 2020; Sultan et al., 2020).
5.14.1.3 Insurance as a climate impact risk management tool
Insurance is a financial adaptation strategy increasingly used in agriculture and aquaculture. A relatively new approach to agricultural insurance risk is the use of financial derivative products, such as index-based agricultural insurance (IBAI), marketed by financial institutions to farmers to help them deal with weather-related production risks (Isakson, 2015; Jensen and Barrett, 2017). The basic idea is to rely on easily observed weather indices, such as precipitation or temperature, that co-vary with farm production. Insurance payments are received when the metric trigger for a region is reached, eliminating the need to collect farm-specific information. Proponents of index insurance argue that it can resolve the information costs and incentive problems inherent in rural financial markets, such as adverse selection, and allow provision of insurance coverage at a fraction of the costs of loss-based polices (Jensen and Barrett, 2017). Buyers of index policies do not have to prove their ownership of assets with weather-related losses. This lowers transactions costs and makes it more affordable to insure small plots of land.
The creation of index insurance requires significant prior research and extensive data that may not be available or sufficient in lower-income countries, including identifying the most appropriate farm and climate variables to include and financial and regulatory support from the public sector (Economic Commission for Latin America and the Caribbean and Central American Agricultural Council of the Central American Integration System, 2013; Economic Commission for Latin America and the Caribbean and System, 2014). Some insurance providers bundle it with other services, such as fertilizer use or seeds that may not be useful to particular farmers and can increase their overall capital costs (Isakson, 2015). Although proponents see IBAI as a way to mitigate farmers’ risks associated with more variable weather patterns (Greatrex et al., 2015), critics argue that derivative-based insurance products tend to benefit wealthier farmers and fail in assisting the poorest and most marginalised farmers (Isakson, 2015; Taylor, 2016). Thus far, there is low agreement and medium evidence regarding the adaptation potential of derivatives-based insurance products, signalling a need for further research in this area.
5.14.1.4 Community-based adaptation approaches
Community-based adaptation (CbA) strategies, which involve locally driven, place-based adaptation approaches, can help build adaptive capacity to climate change impacts, but require explicit attention to power dynamics, respect for local and Indigenous knowledge systems, adequate resources, future climatic trends and coordination at multiple levels of governance to be effective (high confidence) (Spires et al., 2014; Fernández-Giménez et al., 2015; Nagoda, 2015; Ashley et al., 2016; Berner et al., 2016; Ensor et al., 2016; Avtar et al., 2019; Lam et al., 2019; Silwal et al., 2019; McNamara et al., 2020; Piggott-McKellar et al., 2020; Rossa, 2020; Uchiyama et al., 2020). Since AR5, there is strong evidence that participation of local stakeholders in adaptation planning and implementation improves communities’ capacity to monitor and respond to climate change impacts on food, fibre and forestry systems, provided that adequate resources and local knowledge on climate change exist. Participatory monitoring of climate change impacts and participatory scenario development to develop community action plans are examples, which can help strengthen community preparation for and response to climate impacts.
Community-based monitoring of forests, coral reefs, seagrass and mangroves are examples of local natural resource assessment that can support food security and livelihoods while informing regional and national climate change planning tools (Carter et al., 2014; Gevaña et al., 2018; Avtar et al., 2019). Negotiation among many stakeholders at multiple scales, including inclusive mechanisms to address power inequities in governance structures and communities, may be needed for CbA to be effective (Avtar et al., 2019; McNamara et al., 2020). Indigenous knowledge and community-based management of fisheries and aquaculture in the Arctic and Asia (Roux et al., 2019; Chen and Cheng, 2020; Galappaththi et al., 2020a; Schott et al., 2020; Galappaththi et al., 2021) provide adaptive strategies for sustainable use. (Iticha and Husen, 2019). Community-based climate services in the Andes (managed through a collaboration of smallholder producers and an international partnership) built capacity and knowledge of climate change dynamics as well as trust in local climate institutions, providing meaningful information for regional responses to climate change impacts (Rossa, 2020). Community-based participatory scenario planning can help identify multiple climate stressors and vulnerabilities to develop effective adaptation plans (Fernández-Giménez et al., 2015; Bennett et al., 2016; Cross-Chapter Box MOVING PLATE this chapter).
An assessment of 32 different CbA initiatives in the Pacific Islands, including addressing risks to food security, found high-performing projects had six key entry points: effective methods to improve adaptive capacity, appropriate to the local context, which moved beyond narrow geographical definitions of community to consider equity of impact, and ecosystem-based approaches, jointly addressing climatic and non-livelihood pressures and consideration of future climatic trends (McNamara et al., 2020). Low-performing initiatives, in contrast, were not sustained; these overlooked future climatic trends in their initiatives, such as beehive susceptibility to climate extremes, and had dependent, unequal relationships that lacked genuine local approval or ownership and did not fit local values and context (Spires et al., 2014; McNamara et al., 2020; Piggott-McKellar et al., 2020). CbA initiatives can also suffer from not having adequate local knowledge of potential strategies to address future climatic scenarios, and may lead to maladaptation, increasing socioeconomic inequities in communities (Nagoda, 2015). Addressing inequity in power dynamics and building technical adaptive capacity of local people are some of the ways that CbA initiatives can support more resilient food systems (McNamara et al., 2020).
5.14.1.5 Local and regional food systems’ strengthening and food sovereignty
Food sovereignty brings together adaptation options based on agroecological methods, access to resources, collective and CbA (HLPE, 2019). Addressing food security and nutrition in light of climate change impacts and vulnerabilities is considered to arise from a mixture of globalised supply chains and local production, not one or the other (Blesh et al., 2019; Stringer et al., 2020). Evidence on strengthening local and regional food systems with a food sovereignty approach, in terms of access to resources (land, seeds, water), shortened food chains and CbA strategies suggest that these strategies can positively contribute to climate change adaptation in many contexts (medium confidence) (SRCCL) but can also lead to conflict especially regarding management of mobile resources such as fisheries (Section 5.8, Cross-Chapter Box MOVING PLATE this chapter). All these options can build adaptation through actions that strengthen local capacities and the power to act within food systems. Securing and recognising tenure for Indigenous Peoples (Hurlbert et al., 2019) and local communities (Oates et al., 2020) can improve their ability to adapt by increasing the incentive to invest in resilient infrastructure and sustainable land management practices. Community seed banks and networks strengthen local seed systems and realise farmers’ rights favouring access to a variety of local genetic resources, with landraces often more adapted to the local social, cultural and ecological environment and needs, and better adapted to harsh environments without external inputs (Mousseau, 2015; Bisht et al., 2018; Maharjan and Maharjan, 2018; Otieno et al., 2018; Mbow et al., 2019). This plays a key role in PPB (Section 5.4.4.5; FAO, 2019e). The integration of informal and formal seed system elements is important for the adaptive capacity of smallholder farmers (Westengen and Brysting, 2014; Westengen and Berg, 2016; FAO, 2019e).
Strengthening both local and regional food systems is a strategy to increase resilience (Schipanski et al., 2016; Palmer et al., 2017), resource use efficiency (Mu et al., 2019) and self-reliance (medium evidence, low agreement ) (Griffin et al., 2015; Chapin et al., 2016; Karg et al., 2016). Collective trademarks (Quiñones-Ruiz et al., 2015) and participatory guarantee systems (Niederle et al., 2020) are examples of innovative institutional strategies to strengthen local and regional food systems. In the urban context, the city region food system (CRFS) approach is motivated by reducing dependence on international trade and associated instability and to facilitate local decision making (Karg et al., 2016). CRFS includes a network within a regional landscape around one urban centre and surrounding peri-urban and rural regions (Blay-Palmer et al., 2018). UPA is promoted as an effective strategy to adapt to climate change in different contexts (see Section 5.12.5.3, Dubbeling, 2015; Lwasa et al., 2015). To cope with the effects of climate change, strengthening regional food systems is becoming an explicit part of urban and regional policy, being tested in many different cities worldwide (Dubbeling et al., 2017; Blay-Palmer et al., 2018; Berner et al., 2019; Sellberg et al., 2020; van der Gaast et al., 2020). Strengthening both local and regional food systems has to be balanced against limitations and trade-offs, since modelling exercises of regionalisation scenarios show urban agriculture cannot achieve food security in areas with rapid population growth (Le Mouël et al., 2018). Furthermore, international trade can compensate in cases where the regional system fails due to extreme events or other related climate shocks (Section 5.11.8).
Box 5.11: Agroecology as a Transformative Climate Change Adaptation Approach
Agroecological approaches can increase food system resilience (robust evidence, medium agreement ), while some agroecological practices such as agroforestry can provide mitigation measures (medium confidence) (Section 5.10.4.2, Table Box 5.11.1, Altieri et al., 2015; Martin and Willaume, 2016; HLPE, 2019; Bezner Kerr et al., 2021; Snapp et al., 2021). Studies testing agroecological approaches have shown robust evidence, medium agreement of increasing adaptation effectiveness through reducing risk, improving food security and yield stability, reducing input costs, and other supporting and provisioning ecosystem services (Section 5.4.4.4 Diacono et al., 2017; Pandey et al., 2017; Schulte et al., 2017; Calderón, 2018; Bezner Kerr et al., 2019; Côte et al., 2019; Rosa-Schleich et al., 2019; Bezner Kerr et al., 2021; Snapp et al., 2021). Effective locally relevant agroecological approaches involve participatory processes, co-creation of knowledge with farmers and attention to social inequities (Bezner Kerr et al., 2021; Santoso et al., 2021; Snapp et al., 2021). To address smallholder vulnerability to climate change impacts, however, additional policy support beyond agroecology will be needed that is context specific; for example, addressing farmer capacity, limited political power to access land, water, seeds and other key natural resources, structural gender inequities, policy and market disincentives that support large-scale monocultures (high confidence) (Anderson et al., 2019a; HLPE, 2019; Holt-Giménez et al., 2021; Snapp et al., 2021).
Table Box 5.11.1 | Dimensions of agroecological transitions as a transformative climate change adaptation strategy, benefits, trade-offs and constraints to implementation.
Different dimensions of agroecological transitions as a transformative climate change adaptation strategy | Links to climate change impacts, benefits, trade-offs and constraints to implementation with examples |
Environmental: Agroecology can support long-term productivity and resilience of food systems by sustaining ecosystem services such as pollination, SOC, pest and weed control, soil microbial activity, crop yield stability, water quality and biodiversity (high confidence, see Section 5.4.4.4, Cross-Working Group Box BIOECONOMY this chapter and Cross-Chapter Box NATURAL in Chapter 2). (Isbell et al., 2017; Kremen and Merenlender, 2018; LaCanne and Lundgren, 2018; Beillouin et al., 2019b; Dainese et al., 2019; Rosa-Schleich et al., 2019; Snapp et al., 2021). |
|
Socio-cultural: Effective locally relevant agroecological approaches involve participatory processes, co-creation of knowledge with farmers and attention to social inequities, in doing so building farmer capacity (HLPE, 2019; Bharucha et al., 2020; Holt-Giménez et al., 2021; Snapp et al., 2021). |
|
Food security and nutrition: Agroecological practices can increase household food security and nutrition for producer households, with more evidence in low- and medium-income countries (high confidence) (Darrouzet-Nardi, 2016; Demeke et al., 2017; Jones, 2017a; Kangmennaang et al., 2017; Pandey et al., 2017; Luna-Gonzalez and Sorensen, 2018; Bezner Kerr et al., 2019; Boedecker et al., 2019; Mulwa and Visser, 2020; Bezner Kerr et al., 2021; Santoso et al., 2021). |
|
Economic: Agroecology can support socioeconomic resilience, through reducing reliance on purchased inputs, enhancing local and regional economies (HLPE, 2019; Bharucha et al., 2020; Holt-Giménez et al., 2021). |
|
Long-term investment : Timeframes are an important consideration, as an agroecological transition involves multiple overlapping stages, of reducing chemical inputs, experimenting with and applying new agroecological practices and adjusting them, redesigning the farm, strengthening short value chains and producer networks (Gliessman, 2014; Padel et al., 2020). |
|
Policy tools: Investment in agroecological approaches that are designed for socio-ecological context, farmer-led schools, co-learning platforms, and networks of farmers, scientists, private sector and civil society can support agroecological transitions at a regional scale (high confidence) (Coe et al., 2014; Catacora-Vargas et al., 2017; Pérez-Marin et al., 2017; Mier y Terán Giménez Cacho et al., 2018; Anderson et al., 2019a; González de Molina, 2020; Lampkin et al., 2020; Padel et al., 2020; Snapp et al., 2021). Policies can provide incentives (e.g., price premiums, access to credit, extension service, taxes, regulation) to support agroecological transitions by producers (HLPE, 2019; Rosa-Schleich et al., 2019; Gerard et al., 2020; SAPEA, 2020). |
|
Other drivers of agroecological transitions can include crises (environmental, economic or social), social movements, changing socio-cultural values, addressing social inequities, and discourse (Pérez-Marin et al., 2017; Mier y Terán Giménez Cacho et al., 2018; Anderson et al., 2019a). | Further research could provide context-specific information about economic and ecological benefits of some practices and combinations, with effective policies to support their implementation (high confidence) (HLPE, 2019; Rosa-Schleich et al., 2019; Snapp et al., 2021). Institutional support to monitor the ecosystem services climate change mitigation and adaptation impact of agroecological systems can inform policy, using systematic methods and indicators (e.g., Barrios et al., 2020; Mottet et al., 2020) including annual reporting to the United Nations Framework Convention on Climate Change (UNFCCC) (Snapp et al., 2021). |
Box 5.11
Box 5.11
5.14.2 Enabling Conditions for Implementing Adaptation
5.14.2.1 Addressing social inequities in food systems
Addressing gender and other social inequities (e.g., racial, ethnicity, age, income, geographic location) in markets, governance and control over resources is a key enabling condition for climate-resilient transitions in land and aquatic ecosystems (high confidence) (Pearse, 2017; Vermeulen et al., 2018; Blesh et al., 2019; Rao et al., 2019b; Cross-Chapter Box GENDER in Chapter 18, Section 5,13,1; Tavenner et al., 2019). Adaptation strategies can have negative impacts on marginalised social groups and worsen socioeconomic inequities unless explicit efforts are made to address unequal power dynamics and differences in access to resources in agricultural, fisheries, aquaculture, livestock and forestry systems (high confidence) (Glemarec, 2017; Haji and Legesse, 2017; Nagoda and Nightingale, 2017 ; Nightingale, 2017; Rao et al., 2019b; Huyer and Partey, 2020; Mikulewicz, 2020; Taylor and Bhasme, 2020; Eriksen et al., 2021). Technical approaches to adaptation that ignore inequities can worsen them; see, for example, the case study on Climate Smart Agriculture (Box 5.12). Enabling environments support inclusive decision making, capacity building, shifts in social rules, norms and behaviours and access to resources for marginalised groups for climate change adaptation (e.g., Tschakert et al., 2016; Ziervogel, 2019; Eriksen et al., 2021; Garcia et al., 2021).
5.14.2.2 Incorporating Indigenous knowledge and local knowledge
Indigenous knowledge (IK) and local knowledge (LK), while an important component of many adaptation strategies (Reyes-García, 2014; Roue, 2018), continues to be marginalised in food systems; greater integration will increase effectiveness (high confidence) (Ford et al., 2015; Brugnach et al., 2017; Figueroa-Helland et al., 2018). Where Indigenous Peoples have access to and control over their lands and natural resources, food systems can potentially be more sustainably managed and more resilient (high confidence) (Rumbach and Foley, 2014; O’Connell-Milne, 2015; Camacho et al., 2016; Janhiainen, 2017; Kihila, 2018). For example, on Solomon Islands, community-based adaptation combining with IK-informed community mapping helped boost agricultural yields sustainably (Leon et al., 2015), and in China people living in rich plant resource regions have used their wild plants IK to complement the decrease of crop yields during extreme droughts to ensure food security (Zhang et al., 2016). These cases have led scientists and local communities to call for more practical actions to bridge local knowledge, IK and formal science (Borquez et al., 2017; Klenk et al., 2017; Mukhopadhyay, 2017; Olorunfemi, 2017; Reyes-Garcia et al., 2019). Despite this increased public and scientific recognition, IK is often not acknowledged or used.
Effective adaptation requires a more holistic approach that includes the recognition of Indigenous rights, governance systems and laws (high confidence) (Robinson et al., 2016a; Brugnach et al., 2017; Magni, 2017; McMillen et al., 2017; McNeeley, 2017; Pearce et al., 2018), and to couple IK with proactive and regionally coherent adaptation plans, actions and cooperation (Shaffer, 2014; Melvin et al., 2017; Forbis Jr. and Hayhoe, 2018; Makondo and Thomas, 2018).
Supporting Indigenous groups’ knowledge and other excluded social groups can help preserve and harness underutilised resources to enhance nutritional and economic security, with careful measures in protecting Indigenous intellectual rights and avoiding commodification exploitation (Nakashima et al., 2012; Nandal and Bhardwaj, 2014; Ghosh-Jerath et al., 2015; Ebert, 2017). In some regions, there has been a loss of IK about food systems, reducing adaptive capacity (Richards et al., 2019; Panikkar and Lemmond, 2020). Knowledge exchange between Indigenous elders and youth can support adaptive capacity (Osterhoudt, 2018; Richards et al., 2019; Zin et al., 2019). Education utilising IK and LK can help prevent maladaptation options (high confidence) (Melvin et al., 2017; Taremwa, 2017; Forbis Jr. and Hayhoe, 2018; Narayan et al., 2020). There are examples of integrating IK and LK into resource management systems and school curricula and in local institutions with existing decision-making process to strengthen their capacity to address climate change (Huaman and Valdiviezo, 2014; McNamara and Prasad, 2014; Abah et al., 2015; Mistry and Berardi, 2016; Tschakert et al., 2017; McNeeley et al., 2018; McNeeley et al., 2020). However, there are limitations of IK and LK to address future climate impacts. Therefore, it is important that science-based knowledge and other knowledge coalesce to produce solutions that are sustainable and viable in the face of projected impacts of climate change. Community-based adaptation approaches can integrate IK and LK and more formal knowledge systems, provided efforts to establish relationships of respect, trust and common understanding between different stakeholders involved (Herath et al., 2015; Camacho et al., 2016; Fidelman et al., 2017; Inaotombi and Mahanta, 2019; Lam et al., 2019).
5.14.2.3 System transformation and policy enablers
Recent literature highlights the future challenges of producing the quantities of food needed to feed a growing world population in a way that satisfies nutritional needs, benefits everyone equally and equitably, and minimises the negative impacts of food systems on the environment and the natural resource base. There is broad agreement that current trajectories towards the SDGs and countries’ commitments under the Paris Agreement are slow and that transformation of food systems is needed (medium agreement , robust evidence) (Campbell et al., 2018; Brondizio et al., 2019; Dury et al., 2019; EAT-LANCET, 2019; FAO, 2019 f; Food and Land Use Coalition, 2019; Sachs et al., 2019; Searchinger, 2019a; Searchinger T, 2019b; Loboguerrero et al., 2020; Meridian Institute, 2020; Steiner A, 2020).
Recent reviews have summarised literature on production system transformations, driven at least in part by a changing climate or changing climate variability. Such transformations may involve sometimes substantial shifts in farm and livelihood enterprises and land configurations, including intensification, diversification, sedentarisation and abandonment of agriculture (Vermeulen et al., 2018; Thornton et al., 2019). Relevant literature is summarised in Table 5.24, showing reported farmers’ perceptions of the drivers of change and the different outcomes of these changes. The consequences of these production system transitions have been mixed; in about 40% of cases, the outcomes at household level have been unequivocally beneficial. In the other cases, there were detrimental effects on livelihoods, or a mixture of positive and negative effects. The effects on nutritional security reported in these studies were limited. Different enablers of change appear critical if transitions are to have positive outcomes. Policy environments, defined in terms of multi-level governance structures and institutions, are a key driver of systems change, as well as being enablers of and barriers to adaptation responses (Xu et al., 2008; Namgay et al., 2014; Galvin et al., 2015; Schmidt and Pearson, 2016; Liao and Fei, 2017). Policies around property and grazing rights are directly linked to small-scale food producer vulnerability, and land ownership changes will pose a key challenge as climate change impacts in the marginal lands intensify (Reid et al., 2014). Collective action at multiple scales and effective governance structures are also a key enabler of transformational change, for helping community initiatives overcome economic, social and technical barriers, and to strengthen social capital and farmer knowledge (Haglund et al., 2011; Reed et al., 2017; Vermeulen et al., 2018; Fedele et al., 2019). Market development has been shown to be a critical factor for successful adaptation at scale in sub-Saharan Africa (Ouédraogo et al., 2017; Iiyama et al., 2018; Totin et al., 2018). At the same time, financing mechanisms may be a crucial enabler for different food system actors: de-risking agricultural production and food system investments for producers and input suppliers, for example, that address core market failures and compensate actors for extra short-term costs that can lead to longer-term benefits, particularly for small-scale producers and businesses with comparatively low access to technologies and services (Vermeulen et al., 2018; Millan, 2019; see Section 5.1 4.2.5).
The examples in Table 5.24 highlight the uneven impact of adaptation programmes and projects in general, due in part to differences in institutional support and failure of policies to take into account inequities (Clay and King, 2019; Nightingale et al., 2020). Focusing on transformational adaptation, Vermeulen (2018) suggested the need to expand the remit of adaptation planning to consider the multi-functionality of agriculture and a system-wide view of food production and consumption. Several authors argue that transformational change must address the personal, practical and political spheres, in view of the role of power relations and worldviews in shaping practices, food security and inequity (O’Brien, 2015; Nightingale, 2017; O’Brien, 2018; Eriksen et al., 2019; Gosnell et al., 2019). If it involves new or unfamiliar technology, transformation may also be highly disruptive, and the added vulnerabilities of food system actors at risk will need to be addressed (Herrero et al., 2020; see Box 5.5).
Table 5.24 | Agricultural and livelihood system transformations from systematic searches of the literature, which are at least partially attributable to climatic factors and that involve increased or decreased system integration, and major consequences of the change. Information in the table is from the references cited. Sources: updated from (Vermeulen et al., 2018; Thornton et al., 2019).
Underlying production system | Primary drivers of change as stated | Major processes of change as reported | Consequences of change, if reported | Reference |
Extensive grassland-based systems | ||||
Extensive grassland-based, northwest China | Government policy, climate | Sedentarisation Diversification (crops, wages) | Income decline, asset holding decline | Liao and Fei, (2017) |
Extensive grassland-based, Peruvian Andes | Multiple climatic and non-climatic drivers | Diversification (wages, livestock assets, land) Extensification | Livestock accumulation in wealthy households, asset diversification in poorer households | López-i-Gelats et al., (2015) |
Extensive grassland-based, Bhutan | Government policy, labour constraints, climate | Sedentarisation Diversification (crops) Exit | Increased risk, loss of cultural identity, improved market access, livelihood ‘lock-in’ (inability to change rapidly) | Namgay et al., (2014) |
Extensive grassland-based, Borana, Ethiopia | Increase in climate variability, resource degradation | Livestock herd diversification (more small stock and camels, fewer cattle) | Enhanced household resilience | Megersa et al., (2014) |
Extensive grassland-based, Tibetan Plateau | Government policy, climate | Sedentarisation Diversification (crops, off-farm wages, trade) | Increased food production, increased disease burden | |
Extensive grassland-based, Afar, Ethiopia | Government policy, climate | Sedentarisation Diversification (crops) | Weakened institutions and cultural practices, deteriorating natural resources | |
Extensive grassland-based, Kajiado, Kenya | Government policy, climate, population growth | Sedentarisation Diversification (crops, wages, remittances) Intensification | Nutritional status remains poor | |
Extensive grassland-based, Mongolian Altai | Government policy, climate | Sedentarisation Diversification (cashmere sales, forest products) | Fodder shortages, forest over-use, unsustainable land use system | |
Extensive grassland based, Mongolia | Increasing drought, grassland degradation | Diversification (decreases in sheep and goats, increases in cattle, decreases in grain production, increases in fruit and vegetable production) Exit from agriculture | Increased household income from off-farm employment, more diverse diets | |
Extensive grassland-based, northern Kenya | Climate change and variability | Diversification (crops, wages, migration) | Decreasing adaptive capacity, over-dependence on local knowledge for adaptation | |
Extensive systems with crops | ||||
Extensive with crops, Eastern Cape, South Africa | Multiple | Intensification (richer households) Exit and abandonment (poorer households) Livelihood diversification | Wildlife conflicts, loss of cultural identity | |
Extensive with crops, Peruvian highlands | Economic globalisation, climate change | Diversification (dairy production, wage migration) Conversion (away from staple crops to feed production) Intensification (feed production) | Reduced vulnerability to climate change, but potential loss of both agrobiodiversity and food self-sufficiency identified by the author | |
Extensive with crops, East Africa | Climate | Diversification (crops, livestock, wages) Intensification (crops, intercrops) | Increasing household vulnerability | |
Extensive with crops, Ghana | Climate variability, temperature change | Diversification (off-farm activities) | Reduced vulnerability | |
Extensive smallholder cropping, Nepal | Annual and seasonal warming. Increased precipitation with changes in patterns. | Diversification and integration (from growing buckwheat and barley to vegetables and fruit trees) | Increased household resilience due to diversification of production | |
Extensive smallholder mixed system, Niger | Droughts and famines, and land degradation | Large-scale regeneration of native trees and shrubs in the arable landscape | Increased household income, effects on household food security not yet known | |
Other mixed coastal and forest systems | ||||
Coastal rice-based, Bangladesh | Increased salinity due to reduced dry season flows from rivers in India, use of groundwater for irrigation | Diversification (from rice cultivation to aquaculture of shrimp and prawn) | Increased household income, increased engagement of women, increased human disease vulnerability | |
Smallholder cropping systems, coastal Bangladesh | Increasing frequency and severity of floods since 2008 | Diversification (re-allocation of land from crops to aquaculture) Exit (migration away from village) | Mixed impacts on household incomes and seasonal migration frequency | Fenton et al. (2017) |
Smallholder mixed cropping in forested landscapes in Indonesia | Floods, drought, crop and livestock disease | Diversification (re-allocation of land from forests to rubber plantations and rice) Intensification (agroforestry) Extensification (reforestation, forest protection) | Locally, increased household incomes in general; more widely, some trade-offs with biodiversity, water, carbon stocks | |
‘Transformation’, defined by IPCC (2019a) as ‘a change in the fundamental attributes of natural and human systems’, is defined here as a redistribution of at least a third in the primary factors of production (land, labour, capital) and/or the outputs and outcomes of production (the types and amounts of production and consumption of goods and services arising from multi-functional agricultural systems) (Vermeulen et al., 2018; Thornton et al., 2019).
5.14.2.4 Finance needs and strategies for adaptation
Current understanding of finance flows and needs for adaptation in crop agriculture, livestock, fisheries, aquaculture and forest products relies primarily on top-down projections, with limited data (UNFCCC, 2018; Buchner et al., 2019; Jachnik et al., 2019). By one estimate, in 2017/2018, agriculture, forestry and land use received 24% of public adaptation finance (totaling USD 7 billion; half via multilateral development finance institutions and one-quarter from governments) and 35% of international grants (with 71% used for adaptation) (Buchner et al., 2019). According to data from OECD (2020), finance flows for agriculture, forestry and fisheries have risen fairly linearly from ca. USD 1.46 billion in 2010 (the year the Rio marker on climate change adaptation was introduced) to ca. 5.5 billion in 2018. Over the entire tracked period, the three subsectors combined received a total of USD 29.82 billion for activities with principal and significant adaptation components. 4 However, the data set only includes climate-related development finance from bilateral, multilateral and private philanthropic sources, whereas private sector finance flows are not captured as this is notoriously difficult to track (UNEP, 2016; OECD, 2020; cross-ref to Cross-Chapter Box FINANCE in Chapter 17). Most of the funding (85%) was directed towards agriculture, with forestry (12%) and fisheries (3%) receiving significantly less, but across the subsectors, there is consistency in the sense that policy and administrative management and development receive the lion’s share of support, which is predominantly given in the form of grants (72%), while debt instruments (26%) and equity and shares in collective investment vehicles (2%) contribute less. From a regional perspective, 80% were directed to Africa (47%), Asia-Pacific (27%), and Latin America and Caribbean States (7%), whereas Eastern Europe and Western Europe and Other States received (2%) each and 17% were destined for ‘developing countries’ without regional tags. Finally, it is noteworthy that 38% of adaptation finance in agriculture, forestry and fisheries is marked as also having mitigation benefits, and roughly a quarter of funding is reported as having principal or significant gender objectives.
Whether current levels of growth in adaptation finance for agriculture, forestry and fisheries is keeping up with estimated needs cannot be assessed because of the large uncertainties that surround adaptation cost estimates (Cross-Chapter Box FINANCE in Chapter 17). There is, hence, high agreement that better assessment of adaptation costs of climate impacts requires considerably more research (Watkiss, 2015; Diaz and Moore, 2017). A recent study focusing on investments needed to offset the effects of climate change on the prevalence of hunger concludes that investments in agricultural research and development (R&D) have to increase from USD 1.62 billion to USD 2.77 billion per year between 2015 and 2050 (Sulser et al., 2021a). In addition to agricultural R&D, significant investment increases in water and infrastructure in the range of USD 12.7 billion and USD 10.8 billion are required, respectively, a considerable portion of which is relevant to the food system. In total, Sulser et al. (2021a) estimate that annual investment between USD 21.47 billion and USD 29.8 billion are needed to avoid sliding back from climate-change-related increases in the prevalence of hunger but recognise the shortcomings of their approach and acknowledge that ‘a full analysis of adaptation to climate change in agriculture would require including many other social, economic, and environmental dimensions’. For comparison, World Bank (2010) estimated global costs of USD 70–100 billion per year for agriculture, forestry and fisheries, infrastructure, water resources, health, ecosystem services, coastal zones and extreme weather events to adapt to an approximately 2°C warmer world between 2010 and 2050. While the World Bank includes more sectors, more recent publications consider the resulting figures to be significantly too low (Baarsch et al., 2015; UNEP, 2016; Rossi and Miola, 2017; Hallegatte et al., 2018; Markandya and González-Eguino, 2019; Chapagain et al., 2020; WGII Cross-Chapter Box FINANCE in Chapter 17). Therefore, despite the methodological and data challenges, further efforts are needed to better capture the economic risks of climate change and provide estimates of adaptation costs at global to national scales as well as across sectors (Watkiss, 2015; Diaz and Moore, 2017).
Financial barriers limit implementation of adaptation options in agriculture, fisheries, aquaculture and forestry (high confidence) (Shukla et al., 2019; FAO et al., 2020). Finance strategies can contribute to adaptation in these sectors in different ways (Table 5.25) and to different degrees. Standardised strategies have not yet been developed for specific adaptation needs, and in current practice, finance strategies are opportunistically deployed, with developing countries facing particular challenges due to under-developed financial mechanisms (Omari-Motsumi et al., 2019).
Table 5.25 | Potential adaptation finance strategies for categories of climate-related risks in the agriculture, fisheries, aquaculture and forestry sectors.
Finance strategies | Reduced food availability | Low food safety /dietary health | Diminished livelihoods | Declining ecosystem services |
Reduce vulnerability | Avoid staple failure: Vouchers to producers for improved production inputs | Diversify production strategies: Invest in alternative crops/species/harvest methods | Increase producer capacity: Fund technical assistance programmes | Incentivise improved management : Improved access to credit based on environmental performance |
Anticipate/minimise impacts | Minimise impact of extreme weather: Fund early-warning systems | Diversify products in supply chains: Finance processing equipment for alternative food products | Moderate food price spikes: National food reserves | Minimise resource depletion: Subsidise micro-lending for water-efficient technologies |
Steer capital towards climate resilience | Develop climate-resilient production technologies: Fund R&D for improved genetics (crops, fish, livestock) and management | Build nutrition-sensitive food systems: Finance early-stage market building for diversified food products | Increase resilience of supply chain infrastructure: Finance improved storage and transport facilities | Disincentivise low-resilience production: Screen investments based on climate risk disclosures |
Pool climate-related risks | Distribute climate-related risks: Securitise investments in production systems | De-risk diversified food supply chains: Invest in producer aggregation to improve supply chain efficiency | Insure against supply chain risks: Subsidised index insurance programmes | Detect high-risk production systems: Invest in supply chain monitoring/traceability mechanisms |
Compensate for climate-related impacts | Compensate for production losses: Financial transfers to affected producers | Avoid food shortages: Subsidise food importation | Avoid selling off productive assets: Fund social support for low-income households | Ecological restoration: Direct development aid to land rehabilitation projects |
Many types of financial instruments are employed by diverse actors (Table 5.26) guided by their mandates (e.g., development, commerce), capacity (investor, intermediary, donor) and risk appetite. Actors within a sector or local production area can coordinate their financial strategies towards common objectives (e.g., reduced supply chain loss) or participate in joint financial action such as blended finance structures that combine commercial and concessionary finance to catalyse additional private investment, enrich the pipeline of bankable projects, and test business models (FAO, 2020b).
Table 5.26 | Potential adaptation finance objectives for major actors in agriculture, fisheries, aquaculture and forestry sectors.
Actors | Potential adaptation finance objectives |
Private sector: Focused on capturing positive externalities (i.e., lower risks or costs) from adaptation investments (Woodard et al., 2019). Major considerations include fiduciary responsibilities; expected rates of return (i.e., risk-adjusted; benchmarked to comparable investments); investment characteristics (e.g., liquidity, structure, size) and contribution to investor portfolio; material business risks (e.g., supply chain reliability; stranded assets); cost control (e.g., product losses; insurance); legal compliance; and sectoral requirements (e.g., climate risk disclosure) (Havemann et al., 2020). | |
Production companies or cooperatives |
|
Financial investors and intermediaries (e.g., banks, asset managers, venture capital; non-bank financial institutions) |
|
Public sector: Encompassing nearly commercial (e.g., specialised commodity boards; bond issuances), partially subsidised (e.g., low-interest loans) and fully subsidised (e.g., R&D; grants) investments. Major considerations include avoiding negative impacts to citizens (e.g., food price spikes) and specific constituencies (e.g., catastrophic losses to producers) and maintaining/enhancing public revenues (i.e., taxes from economic activity in agriculture, fisheries, aquaculture and forestry). | |
Government agencies and multilateral institutions |
|
Expanding access to financial services and pooling climate risks can enable and incentivise climate change adaptation (medium confidence) (Shukla et al., 2019). To mobilise financial instruments (Table 5.27) towards adaptation needs, individual actors can apply an adaptation lens to existing or new activities, accounting for investment characteristics (e.g., development stage; cash flow profile), requirements (e.g., amount; risk–return) and context (e.g., regulatory landscape) (Havemann et al., 2020). Risk-layering can match financial instruments to severity and probability climate risks (Hochrainer-Stigler and Reiter, 2021).
Table 5.27 | Major types of financial instruments suitable to adaptation finance in agriculture, fisheries, aquaculture and forestry sectors (adapted from Havemann et al., 2020).
Financial instrument | Description |
Equity: Ownership stake in a company (e.g., agricultural technology company; processing company) or collective investment vehicle (e.g., agriculture fund; Timber Investment Management Organization; commodity index fund) providing returns (via dividends and/or sale of equity shares) corresponding to business-related risk (e.g., higher return for higher risk and/or lower liquidity) | |
Listed equities | Ownership of shares in a company listed in a public market |
Private equity | Ownership of shares in a company or other assets |
Junior or risk-absorbing equity | Ownership of lower-tier shares in a company (e.g., common stock) or collective investment vehicle (e.g., first-loss tranche) |
Debt : Capital provided directly or indirectly (via banks or other third-party institutions) to users with defined repayment terms (i.e., timeframe, interest rate); more likely to deliver adaptation benefits when coupled with capacity building (e.g., technical assistance, education, analytics) (Woodard et al., 2019) | |
Loan, bond, note, credit line | Direct or indirect provision of capital (e.g., operating loans; dedicated credit line for agricultural trade); concessionary loans may allow for below-market interest rates |
Soft loan | Direct interest-free loan (e.g., funds provided in advance of good/service delivery) |
Emergency loan | Lending in response to climate risks or impacts with repayment terms (e.g., return period) that consider necessary relief, recovery and reconstruction |
Catastrophe bond | Risk transfer instrument in which insurers or reinsurers provide high interest payments to investors in exchange for a payout (and repayment deferment or forgiveness) activated by specific events (e.g., extreme weather) |
Impact bond | Subsidised investment providing capital upfront or based on defined outcomes |
Subordinated loan | Concessionary capital with a junior position (i.e., accepting higher risk of non-repayment and / or lower rate of return on investment) relative to other investors |
Securitised investments | Aggregation of equity or debt to offer marketable securities to a wider pool of investors with different risk–return appetites |
Guarantees: Commercial and concessionary guarantees that provide compensation for losses due to specified risks (e.g., political risk, performance risk); more likely to deliver adaptation benefits when linked to robust underwriting standards and verification protocols (Woodard et al., 2019) | |
Credit guarantee | Compensation for specified losses incurred by agricultural lenders |
Payment, performance, surety bonds | De-risking mechanism for transactions between providers and buyers of goods/services; may be used in trade finance and other forms of intermediation |
Insurance: Policies and other financial instruments that provide compensation for losses based on defined terms and conditions. | |
Production insurance | Compensation for specified losses related to production (e.g., insurance indexed to specific weather events) or supply chains (e.g., shipping insurance) |
Market and price insurance | Compensation for specified market-related losses (e.g., price or currency fluctuation) |
Grants: Concessionary funding provided by public or philanthropic entities to support climate adaptation costs or outcomes (no expectation of repayment) | |
Direct support | Funding for provision of goods (e.g., fertilizer, seeds, nursery stock) or services (e.g., technical assistance, product storage) to producers, local companies or intermediaries (e.g., for agronomic or business management expertise); can reduce credit risk when part of blended finance arrangements |
Performance-based grants | Grants or other concessionary funding contingent on achievement of defined adaptation outcomes (with possible third-party verification requirement); may support development and testing of new approaches (i.e., design funding; challenges/prizes) |
Governmental instruments | |
Policy incentives | Public policies designed to stimulate adaptation action among targeted groups (e.g., producers, consumers, agri-businesses, financiers) including direct or indirect subsidies (e.g., producer payments, tax breaks, health insurance), procurement policies (e.g., low carbon and sustainability criteria; nutrition-sensitive school feeding programmes) and other fiscal measures (e.g., infrastructure development; funding R&D in climate-resilient practices or technologies) (Shukla et al., 2019) |
Development aid | International or domestic programmes that directly or indirectly fund adaptation actions including financial transfers (e.g., producer support or anti-poverty programmes) and subsidised credit (medium confidence) (Shukla et al., 2019) |
Planning grants | Financial support to governments for adaptation planning (e.g., via readiness programmes) |
Other instruments | |
Fintech | Data analytics and risk analysis models used to better assess borrowers’ repayment risk (e.g., due to crop failure) and reduce transaction costs (e.g., streamlined lending processes); applications may include financial inclusion (e.g., micro-financing; lending to small- and mid-size operators), alternative repayment programmes (e.g., for larger capital borrowing), insurance (e.g., more granular risk assessment) or digital strategies (e.g., crowdfunding, smallholder credit) (Agyekumhene et al., 2018) |
Payment for Ecosystem Services (PES) | Funds delivered to land and resource managers in exchange for compliance with specified sustainability practices or environmental outcomes; PES depends on willing payers (i.e., direct and indirect beneficiaries of ecosystem services such as governments, companies, conservation groups, philanthropies) |
5.14.2.5 Constraints on adaptation finance for food, feed, fibre and other ecosystem products
Flow of adaptation finance in the agriculture, fisheries, aquaculture and forestry sectors is impeded by weak measurement and benchmarking of financial and resilience outcomes (Kramer et al., 2019; Negra et al., 2020), and challenges in assessing repayment capacity of investee producers and companies (medium confidence). Immature information systems (e.g., weak analytics, fragmented standards) (Woodard et al., 2019; Negra et al., 2020) inhibit effective due diligence and impact assessment, contributing to uncertainty and low investor confidence (Havemann et al., 2020; NGFS, 2020). Improved characterisation of adaptation finance strategies (e.g., insurance, subsidies, blended finance) requires increased transaction volume (Millan et al., 2019) and analysis of financial (e.g., risk–return profile, investor demand) and resilience (e.g., reduced vulnerability) effects.
Use of climate-resilient financial strategies and instruments is limited by weak incentives, which commonly take the form of high upfront costs (Verdolini et al., 2018), high transaction and intermediation costs (Havemann et al., 2020) and relatively long pay-off time. Tenant producers may not experience benefits from adaptation investments (Woodard et al., 2019). Investors seek low-risk, liquid investments and credit-worthy counterparties (Havemann et al., 2020), yet small- and medium-sized producers and supply chain actors often lack access to formal credit. Given limited experience and weak information for adaptation finance, sub-optimal outcomes may include imbalanced allocation of public and private finance (e.g., to less vulnerable regions and producers; to lower-resilience investments; to short-term benefits) as well as inequitable division of risks and returns (e.g., within blended finance structures) (Clapp, 2017; World Bank, 2018; Attridge and Engen, 2019). Additionally, while risk-sharing finance strategies can deliver adaptation benefits, they do not inherently reduce overall risk and commonly cover only specified types of risks (Kellett and Peters, 2014; Watson et al., 2015).
Methods to strengthen adaptation finance include updating regulations and policies to support adaptation finance instruments (e.g., climate accounting standards), requiring climate-risk disclosure, improved information-sharing among public and private sector actors and devolving funding to local actors (medium confidence) (Global Commission on Adaptation, 2019; Millan et al., 2019).
Box 5.12: Is Climate-Smart Agriculture Overlooking Gender and Power Relations?
Climate-smart agriculture (CSA) is an approach that aims to increase agricultural productivity, enhance food security, adapt to climate change and, where possible, reduce GHG emissions. The effective implementation of climate-smart practices is conceptually linked to an enabling environment in which policies, institutions and finance can re-orient agricultural systems, thereby supporting development and enhancing food security in a changing climate (Lipper et al., 2014; Karttunen et al., 2017). However, the concept has received criticism based on the absence of conceptual clarity of the interrelations between productivity, food security, adaptation and mitigation (Arenas-Sanchez et al., 2019) and because of limited evidence on the efficacy of CSA for achieving adaptation and mitigation outcomes at a global scale (Arslan et al., 2015; Lamanna et al., 2016; Chandra et al., 2018). Some argue that CSA operates within an apolitical framework that tends to minimise issues concerning power, inequity and access, and is overly focused on technical approaches (Taylor, 2017; HLPE, 2019). CSA is explicitly referenced by more than 30 countries in their Intended Nationally Determined Contributions (INDCs) (Ross et al., 2016), but measuring the degree of its implementation still represents a challenge.
There is low agreement , medium evidence on the relationship between CSA and equity (Allen, 2018; Karlsson et al., 2018). CSA can potentially benefit women if they are able to take advantage of improvements in productivity, food security and adaptation decision making as a result of the implementation of CSA practices. Nevertheless, these advantages can be unequally realised given male domination in receiving information and extension services, as well as financial or resource access (Jost et al., 2016). Some (Huyer and Partey, 2020) argue that CSA may undermine gender equity (Collins, 2018), entrench and solidify power (Haapala, 2018), and result in the disproportional allocation of new labour-intensive activities to women (Jost et al., 2016). Uptake of some climate-smart technologies can further marginalise the most disadvantaged local groups (Roncoli et al., 2009; Haapala, 2018). Unequal sharing of benefits and burdens with respect to emission reduction costs among different agricultural groups has also been observed (Budiman, 2019).
In contrast, emerging research points to the potential of CSA as a supporting condition for gender equity, provided that equity and power concerns are explicitly included in the approach (Chanana-Nag and Aggarwal, 2020). Some CSA technologies and practices, such as direct seeding, green manuring and laser land levelling, can have a significant role in reducing the gender gap in labour burden for women in agriculture (Khatri-Chhetri et al., 2020). The use of participatory approaches can facilitate community-based adaptation of gender-sensitive CSA practices (Rosimo, 2018). CSA may also empower both men and women: in two villages in India, CSA adoption empowered both sexes in decision making and use and control of income (Hariharan et al., 2018).
In general CSA programmes have tended to overlook questions of inequity (medium confidence), including limited attention to social conditions that promote Business-As-Usual pathways, although this is now changing. Addressing questions of rights, social injustice, unequal power relations and inequity would help make CSA-related policy responses more effective in addressing vulnerability (Chandra et al., 2017; Clapp and Isakson, 2018; Karlsson et al., 2018; Westengen et al., 2018; Ellis and Tschakert, 2019; Eriksen et al., 2019; Westengen et al., 2019).
Box 5.13: Supporting Youth Adaptation in Food Systems
Young people are key agents in agrifood systems: both a vulnerable group, and one that can foster systemic change (high confidence) (Brooks et al., 2019; Figure X; IFAD, 2019; Flynn and Sumberg, 2021; HLPE, 2021). Food systems are the largest source of employment for young people, but do not always provide adequate livelihoods or decent working conditions (HLPE, 2021). Regions with more youthful populations—such as Sub-Saharan Africa, South Asia and Central America—are both highly vulnerable to climate change impacts and reliant on agriculture, forestry, aquaculture and fisheries for livelihoods (Brooks et al., 2019; IFAD, 2019; HLPE, 2021). Rural youth in these sectors are particularly vulnerable, often with less access to land, water, capital and other resources, shaped by family and social relations, and fewer opportunities (high confidence) (Chingala et al., 2017; Ricker-Gilbert and Chamberlin, 2018; IFAD, 2019; Yeboah et al., 2020; Flynn and Sumberg, 2021; Nhat Lam Duyen, 2021). In these vulnerable regions, climate change compounds other drivers such as poverty to increase youth out-migration to urban areas or other regions (medium confidence) (Zin et al., 2019; Weinreb et al., 2020; HLPE, 2021; Stoltz et al., 2021; Voss, 2021), which can further worsen rural economies. Young low-income rural women may be particularly marginalised and vulnerable due to systemic gender inequities in access to land, credit, employment, institutions and other resources (medium confidence) (Sah Akwen, 2017; IFAD, 2019; Flynn and Sumberg, 2021).
Youth play a critical role in all sectors of the food system (HLPE, 2021; Figure Box 5.13.1), and some are actively pursuing work and innovation in agrifood systems (medium confidence) (Sah Akwen, 2017; 2019; Yeboah et al., 2020; Flynn and Sumberg, 2021). Climate change impacts may reduce youth employment options in food systems in some regions, while they are often politically marginalised (Brooks et al., 2019; IFAD, 2019; HLPE, 2021). At the same time, due to heightened awareness about climate change, youth may be more willing to apply climate adaptation strategies (medium confidence) (Ali and Erenstein, 2017; Jiri et al., 2017; Sah Akwen, 2017; Chamberlin and Sumberg, 2021; Doherty et al., 2021). Agrifood policy implementation of adaptation strategies could increase inclusive participation of youth to meet their needs (HLPE, 2021). Inclusive investments in water management, infrastructure, agrifood science, and policies that increase youth access to land, credit, knowledge, education, skills and other crucial resources can support dignified and rewarding agrifood employment (Ahsan and Mitra, 2016; Brooks et al., 2019; HLPE, 2021). Digital technologies can support agrifood adaptations, but digital divides must be overcome to avoid worsening inequities (HLPE, 2021). Initiatives which protect and strengthen youth engagement and employment in the all points of the food system, including recognition of youth’s critical role and agency through rights-based approaches, can support sustainable food transitions (HLPE, 2021). Harnessing youth innovation and vision to address climate change alongside other SDGs such as gender inequity and rural poverty will be a crucial strategy to ensure resilient economies in food systems (high confidence) (Laube, 2016; Brooks et al., 2019; IFAD, 2019; Abay et al., 2021; HLPE, 2021).
Figure Box 5.13.1 | Youth agency, engagement and employment in food system (HLPE, 2021).
5.14.3 Climate Resilient Development Pathways
Climate resilient development pathways (CRDPs) introduced in AR5 (Denton, 2014) can briefly be described as ‘development trajectories that integrate adaptation and mitigation to realise the goal of sustainable development’ (see IPCC (2019a)) for a more extensive definition). Several characteristics were proposed in SR1.5 by which such CRDPs could be identified: consistency with principles of sustainable development; ability to deliver poverty reduction; ability to enhance social, gender, racial, ethnic and intergenerational equity; ability to deliver resilience to climate change and other shocks and stresses; and ability to protect species, biodiversity and ecosystem goods and services. There is an increasing literature, assessed in SR1.5, on adaptation pathways approaches, generally for specific regions, locations and subsectors.
Two recent examples directly related to agriculture and food are the following: sustaining agrarian livelihoods to mid-century of Nicaraguan small-scale coffee producers using analyses of suitability and coffee quality changes under an IPCC Special Report on Emissions Scenarios (SRES) A2 emissions scenario (Läderach et al., 2017); and development of participatory pathways to mid-century under RCPs 4.5 and 8.5 support regional adaptation planning in Hawke’s Bay, New Zealand for agricultural producers and rural communities (Cradock-Henry et al., 2020). CRDPs mentioned in SROCC include shifting from providing coastal defences to adapting to seawater inundation in coastal regions (Renaud et al., 2015) and retreating coastal megacities (Solecki et al., 2017). Pathway frameworks continue to be used to frame the broad-scale challenges of development and climate change, thereby linking different types of food system actor with different responses through time using a variety of approaches, top-down and participatory, qualitative and quantitative (Butler et al., 2016; Antle et al., 2017; Thornton and Comberti, 2017; Collste et al., 2019; Loboguerrero et al., 2020; Stringer et al., 2020).
While there is consensus that the concept of CRDPs is useful, there are major challenges in identifying, operationalising, monitoring and evaluating them (Lin et al., 2017; Bloemen et al., 2018). Management approaches seldom integrate across spatio-temporal scales and may be unable to address unidirectional change and extreme events (Holsman et al., 2019). The socioeconomic complexities and implications of pursuing integrated outcomes make it difficult to evaluate synergies and trade-offs associated with different actions in local contexts through time (Thornton and Comberti, 2017; Ellis and Tschakert, 2019; Holsman et al., 2019; Orchard, 2019). Case studies by Lo (2019) of transformation in a fishing town in south China and by Gajjar (2019) on undesirable path dependencies in development trajectories in urban and rural India show that overall adaptive capacity of populations may be decreased though politicisation and entrenchment of existing inequities, severely limiting the possibilities for future adaptation. A further challenge of implementation is timely detection of tipping points and abrupt exposure events in both climate and environmental systems (Lenton et al., 2019; Trisos et al., 2020), which may alter the efficacy of current and planned adaptation actions, necessitating a switch to other, more transformational strategies; in such cases, re-energising food system actors’ commitment to adaptation action may well be needed (Bloemen et al., 2018).
Integrated modelling of CRDPs will increasingly be needed to throw light on key SDG synergies and trade-offs into the future (Bleischwitz et al., 2018). In investigating possible future pressures on land under the SSPs, Doelman (2018) projected that the largest changes take place in sub-Saharan Africa in SSP3 and SSP4, mostly because of continued high population growth coupled with (projected) sluggish increases in agricultural efficiency, among other things, leading to expansion of agricultural land for crop and livestock production and reduced food security. Lassaletta (2019) evaluated global pig production in the SSPs and concluded that the future sustainability of pig systems will depend on production efficiency improvements coupled with other factors such as use of alternative feed sources and use of slurries on cropland. Such studies will be increasingly important for quantifying the potential trade-offs and synergies between different SDGs, to guide adaptation (and mitigation) action along CRDPs in the future. The current lack of widely accepted and simple-to-measure indicators for tracking progress in adaptation is a significant hurdle to overcome. There is a large literature on the desirable characteristics of future global food systems, but much less on robust analysis that explicitly addresses and evaluates the pathways towards these desired futures. Gerten (2020) estimates that 10.2 billion people can be supported within key planetary boundaries via spatially redistributed cropland and dietary changes, among other actions. There are few, if any, analyses for detailing the plausible pathways to move towards such a future in ways that are socially, economically and environmentally acceptable through time; whether such pathways could indeed be made climate-resilient is unknown. Appropriate monitoring and rapid feedback to food system actors on what is working and why will be critical to the successful operationalisation of adaptation actions within CRDPs (Bosomworth and Gaillard, 2019).
Cross-Working Group Box BIOECONOMY: Mitigation and Adaptation via the Bioeconomy
Authors: Henry Neufeldt (Denmark/Germany), Göran Berndes (Sweden), Almut Arneth (Germany), Rachel Bezner Kerr (USA/Canada), Luisa F Cabeza (Spain), Donovan Campbell (Jamaica), Jofre Carnicer Cols (Spain), Annette Cowie (Australia), Vassilis Daioglou (Greece), Joanna House (UK), Adrian Leip (Italy/Germany), Francisco Meza (Chile), Michael Morecroft (UK), Gert-Jan Nabuurs (the Netherlands), Camille Parmesan (UK/USA), Julio C Postigo (USA/Peru), Marta G. Rivera-Ferre (Spain), Raphael Slade (UK), Maria Cristina Tirado von der Pahlen (USA/Spain), Pramod K. Singh (India), Peter Smith (UK)
Summary Statement
The growing demand for biomass offers both opportunities and challenges to mitigate and adapt to climate change and natural resource constraints (high confidence). Increased technology innovation, stakeholder integration and transparent governance structures and procedures at local to global scales are key to successful bioeconomy deployment maximising benefits and managing trade-offs (high confidence).
Limited global land and biomass resources accompanied by growing demands for food, feed, fibre and fuels, together with prospects for a paradigm shift towards phasing out fossil fuels, set the frame for potentially fierce competition for land 51 and biomass to meet burgeoning demands even as climate change increasingly limits natural resource potentials (high confidence).
Sustainable agriculture and forestry, technology innovation in bio-based production within a circular economy and international cooperation and governance of global trade in products to reflect and disincentivise their environmental and social externalities can provide mitigation and adaptation via bioeconomy development that responds to the needs and perspectives of multiple stakeholders to achieve outcomes that maximise synergies while limiting trade-offs (high confidence).
Background
There is high confidence that climate change, population growth and changes in per capita consumption will increase pressures on managed as well as natural and semi-natural ecosystems, exacerbating existing risks to livelihoods, biodiversity, human and ecosystem health, infrastructure and food systems (Conijn et al., 2018; IPCC, 2018; IPCC, 2019b; Lade et al., 2020). At the same time, many global mitigation scenarios presented in Intergovernmental Panel on Climate Change (IPCC) assessment reports rely on large greenhouse gas (GHG) emissions reduction in the Agriculture, Forestry, and Other Land Use (AFOLU) sector and concurrent deployment of reforestation/afforestation and biomass use in a multitude of applications (Rogelj et al., 2018; AR6 WGIII Chapter 3 and Chapter 7; Canadell et al., 2021; Lee et al., 2021)
Given the finite availability of natural resources, there are invariably trade-offs that complicate land-based mitigation unless land productivity can be enhanced without undermining ecosystem services (e.g., Obersteiner et al., 2016; Campbell et al., 2017; Caron et al., 2018; Conijn et al., 2018; Heck et al., 2018; WRI, 2018; Smith et al., 2019c). Management intensities can often be adapted to local conditions with consideration of other functions and ecosystem services, but at a global scale the challenge remains to avoid further deforestation and degradation of intact ecosystems, in particular of biodiversity-rich systems (Cross-Chapter Box on NBS-NATURAL in Chapter 2), while meeting the growing demands. Further, increased land use competition can affect food prices and impact food security and livelihoods (To and Grafton, 2015; Chakravorty et al., 2017), with possible knock-on effects related to civil unrest (Abbott et al., 2017; D’Odorico et al., 2018).
Developing New Bio-Based Solutions while Mitigating Overall Biomass Demand Growth
Many existing bio-based products have significant mitigation potential. Increased use of wood in buildings can reduce GHG emissions from cement and steel production while providing carbon storage (Churkina et al., 2020). Substitution of fossil fuels with biomass in manufacture of cement and steel can reduce GHG emissions where these materials are difficult to replace. Dispatchable power based on biomass can provide power stability and quality as the contribution from solar and wind power increases (WGIII Chapter 6), and biofuels can contribute to reducing fossil fuel emissions in the transport and industry sectors (WGIII Chapter 10 and Chapter 11). The use of bio-based plastics, chemicals and packaging could be increased, and biorefineries can achieve high resource-use efficiency in converting biomass into food, feed, fuels and other bio-based products (Aristizábal-Marulanda and Cardona Alzate, 2019; Schmidt et al., 2019). There is also scope for substituting existing bio-based products with more benign products. For example, cellulose-based textiles can replace cotton, which requires large amounts of water, chemical fertilizers and pesticides to ensure high yields.
While increasing and diversified use of biomass can reduce the need for fossil fuels and other GHG-intensive products, unfavourable GHG balances may limit the mitigation value. Growth in biomass use may in the longer term also be constrained by the need to protect biodiversity and ecosystems’ capacity to support essential ecosystem services. Biomass use may also be constrained by water scarcity and other resource scarcities and/or challenges related to public perception and acceptance due to impacts caused by biomass production and use. Energy conservation and efficiency measures and deployment of technologies and systems that do not rely on carbon, such as carbon-free electricity supporting, inter alia, electrification of transport as well as industry processes and residential heating (IPCC, 2018; UNEP, 2019), can constrain the growth in biomass demand when countries seek to phase out fossil fuels and other GHG-intensive products while providing an acceptable standard of living. Nevertheless, demand for bio-based products may become high where full decoupling from carbon is difficult to achieve (e.g., aviation, bio-based plastics, and chemicals) or where carbon storage is an associated benefit (e.g., wood buildings, bioenergy with carbon capture and storage (BECCS), biochar for soil amendments), leading to challenging trade-offs (e.g., food security, biodiversity) that need to be managed in environmentally sustainable and socially just ways.
Changes on the demand side as well as improvements in resource-use efficiencies within the global food and other bio-based systems can also reduce pressures on the remaining land resources. For example, dietary changes towards more plant-based food (where appropriate) and reduced food waste can provide climate change mitigation along with health benefits ( WGIII Chapter 7.4 and 12.4, Willett et al., 2019) and other co-benefits with regard to food security, adaptation and land use (Mbow et al., 2019; Smith et al., 2019c; WGII Chapter 5). Advancements in the provision of novel food and feed sources (e.g., cultured meat, insects, grass-based protein feed and cellular agriculture) can also limit the pressures on finite natural resources (WGIII Chapter 12.4, Parodi et al., 2018; Zabaniotou, 2018).
Cross-Working Group Box BIOECONOMY
Box Cross-Working Group Box BIOECONOMY.1: Circular Bioeconomy
Circular economy approaches (WGIII-12.6) are commonly depicted by two cycles, where the biological cycle focuses on regeneration in the biosphere and the technical cycle focuses on reuse, refurbishment and recycling to maintain value and maximise material recovery (Mayer et al., 2019a). Biogenic carbon flows and resources are part of the biological carbon cycle, but carbon-based products can be included in, and affect, both the biological and the technical carbon cycles (Kirchherr et al., 2017; Winans et al., 2017; Velenturf et al., 2019). The integration of circular economy and bioeconomy principles has been discussed in relation to organic waste management (Teigiserova et al., 2020), societal transition and policy development (Directorate-General for Research Innovation, 2018; Bugge et al., 2019) as well as coronavirus disease 2019 (COVID-19) recovery strategies (Palahi et al., 2020). To maintain the natural resource base, circular bioeconomy emphasises sustainable land use and the return of biomass and nutrients to the biosphere when it leaves the technical cycle.
Biomass scarcity is an argument for adopting circular economy principles for the management of biomass as for non-renewable resources. This includes waste avoidance, product reuse and material recycling, which keep down resource use while maintaining product and material value. However, reuse and recycling is not always feasible, such as when biofuels are used for transport and bio-based biodegradable chemicals are used to reduce ecological impacts where losses to the environment are unavoidable. A balanced approach to management of biomass resources could take departure in the carbon cycle from a value-preservation perspective and the possible routes that can be taken for biomass and carbon, considering a carbon budget defined by the Paris Agreement, principles for sustainable land use and natural ecosystem protection.
Land Use Opportunities and Challenges in the Bioeconomy
Analyses of synergies and trade-offs between adaptation and mitigation in the agriculture and forestry sectors show that outcomes depend on context, design and implementation, so actions have to be tailored to the specific conditions to minimise adverse effects (Kongsager, 2018). This is supported in literature analysing the nexus between land, water, energy and food in the context of climate change, which consistently concludes that addressing these different domains together rather than in isolation would enhance synergies and reduce trade-offs (Obersteiner et al., 2016; D’Odorico et al., 2018; Soto Golcher and Visseren-Hamakers, 2018; Froehse and Schilling, 2019; Momblanch et al., 2019).
Nature-based solutions addressing climate change can provide opportunities for sustainable livelihoods as well as multiple ecosystem services, such as flood risk management through floodplain restoration, saltmarshes, mangroves or peat renaturation (Cross-Chapter Box NATURAL in Chapter 2; UNEP, 2021). Climate-smart agriculture can increase productivity while enhancing resilience and reducing GHG emissions inherent to production (Lipper et al., 2014; Singh and Chudasama, 2021). Similarly, climate-smart forestry considers the whole value chain and integrates climate objectives into forest sector management through multiple measures (from strict reserves to more intensively managed forests) providing mitigation and adaptation benefits (Nabuurs et al., 2018; Verkerk et al., 2020; WGIII Section 7.3)
Agroecological approaches can be integrated into a wide range of land management practices to support a sustainable bioeconomy and address equity considerations (HLPE, 2019). Relevant land use practices, such as agroforestry, intercropping, organic amendments, cover crops and rotational grazing, can provide mitigation and support adaption to climate change via food security, livelihoods, biodiversity and health co-benefits (Ponisio et al., 2015; Garibaldi et al., 2016; D’Annolfo et al., 2017; Bezner Kerr et al., 2019; Clark et al., 2019; Córdova et al., 2019; HLPE, 2019; Mbow et al., 2019; Renard and Tilman, 2019; Sinclair et al., 2019; Bharucha et al., 2020; Bezner Kerr et al., 2021;WGII Cross-Chapter Box NATURAL in Chapter 2). Strategic integration of appropriate biomass production systems into agricultural landscapes can provide biomass for bioenergy and other bio-based products while providing co-benefits such as enhanced landscape diversity, habitat quality, retention of nutrients and sediment, erosion control, climate regulation, flood regulation, pollination and biological pest and disease control (WGIII Chapter12 Box on UNCCD-LDN, Christen and Dalgaard, 2013; Asbjornsen et al., 2014; Holland et al., 2015; Ssegane et al., 2015; Dauber and Miyake, 2016; Milner et al., 2016; Ssegane and Negri, 2016; Styles et al., 2016; Zumpf et al., 2017; Cacho et al., 2018; Alam and Dwivedi, 2019; Cubins et al., 2019; HLPE, 2019; Olsson et al., 2019; Zalesny et al., 2019; Englund et al., 2020). Such approaches can help limit environmental impacts from intensive agriculture while maintaining or increasing land productivity and biomass output.

Figure Cross-Working Group Box BIOECONOMY.1 | Left: High-input intensive agriculture, aiming for high yields of a few crop species, with large fields and no semi-natural habitats. Right: Agroecological agriculture, supplying a range of ecosystem services, relying on biodiversity and crop and animal diversity instead of external inputs, and integrating plant and animal production, with smaller fields and presence of semi-natural habitats. Credit: Jacques Baudry (left); Valérie Viaud (right), published in van der Werf et al. (2020).
Transitions from conventional to new biomass production and conversion systems include challenges related to cross-sector integration and limited experience with new crops and land use practices, including needs for specialised equipment (WGII Chapter 5.10, Thornton and Herrero, 2015; HLPE, 2019). Introduction of agroecological approaches and integrated biomass/food crop production can result in lower food crop yields per hectare, particularly during transition phases, potentially causing indirect land use change, but can also support higher and more stable yields, reduce costs, and increase profitability under climate change (Muller et al., 2017; Seufert and Ramakutty, 2017; Barbieri et al., 2019; HLPE, 2019; Sinclair et al., 2019; Smith et al., 2019c; Smith et al., 2020a). Crop diversification, organic amendments and biological pest control (HLPE, 2019) can reduce input costs and risks of occupational pesticide exposure and food and water contamination (Gonzalez-Alzaga et al., 2014; European Food Safety Authority Panel on Plant Protection Products and their Residues et al., 2017; Mie et al., 2017), reduce farmers’ vulnerability to climate change (e.g., droughts and spread of pests and diseases affecting plant and animal health; Delcour et al., 2015; FAO, 2020a) and enhance provisioning and sustaining ecosystem services, such as pollination (D’Annolfo et al., 2017; Sinclair et al., 2019).
Barriers towards wider implementation include absence of policies that compensate landowners for providing enhanced ecosystem services and other environmental benefits, which can help overcome short-term losses during the transition from conventional practices before longer-term benefits can accrue. Other barriers include limited access to markets, knowledge gaps, financial, technological or labour constraints, lack of extension support and insecure land tenure (Jacobi et al., 2017; Kongsager, 2017; Hernández-Morcillo et al., 2018; Iiyama et al., 2018; HLPE, 2019). Regional-level agroecology transitions may be facilitated by co-learning platforms, farmer networks, private sector, civil society groups, regional and local administration, and other incentive structures (e.g., price premiums, access to credit, regulation) (Coe et al., 2014; Pérez-Marin et al., 2017; Mier y Terán Giménez Cacho et al., 2018; HLPE, 2019; Valencia et al., 2019; SAPEA, 2020). With the right incentives, improvements can be made with regard to profitability, making alternatives more attractive to landowners.
Governing the Solution Space
Literature analysing the synergies and trade-offs between competing demands for land suggest that solutions are highly contextualised in terms of their environmental, socioeconomic and governance-related characteristics, making it difficult to devise generic solutions (Haasnoot et al., 2020). Aspects of spatial and temporal scale can further enhance the complexity, for instance where transboundary effects across jurisdictions or upstream–downstream characteristics need to be considered, or where climate change trajectories might alter relevant biogeophysical dynamics (Postigo and Young, 2021). Nonetheless, there is broad agreement that taking the needs and perspectives of multiple stakeholders into account in a transparent process during negotiations improves the chances of achieving outcomes that maximise synergies while limiting trade-offs (Ariti et al., 2018; Metternicht, 2018; Favretto et al., 2020; Kopáček, 2021; Muscat et al., 2021). Yet differences in agency and power between stakeholders or anticipated changes in access to or control of resources can undermine negotiation results even if there is a common understanding of the overarching benefits of more integrated environmental agreements and the need for greater coordination and cooperation to avoid longer-term losses to all (Aarts and Leeuwis, 2010; Weitz et al., 2017). There is also the risk that strong local participatory processes can become disconnected from broader national plans, and thus fail to support the achievement of national targets. Thus, connection between levels is needed to ensure that ambition for transformative change is not derailed at the local level (Aarts and Leeuwis, 2010; Postigo and Young, 2021).
Decisions on land uses between biomass production for food, feed, fibre or fuel, as well as nature conservation or restoration and other uses (e.g., mining, urban infrastructure), depend on differences in perspectives and values. Because the availability of land for diverse biomass uses is invariably limited, setting priorities for land use allocations therefore first depends on making the perspectives underlying what is considered as ‘high-value’ explicit (Fischer et al., 2007; Garnett et al., 2015; de Boer and van Ittersum, 2018; Muscat et al., 2020). Decisions can then be made transparently based on societal norms, needs and the available resource base. Prioritisation of land use for the common good therefore requires societal consensus-building embedded in the socioeconomic and cultural fabric of regions, societies and communities. Integration of local decision making with national planning ensures local actions complement national development objectives.
International trade in the global economy today provides important opportunities to connect producers and consumers, effectively buffering price volatilities and potentially offering producers in low-income countries access to global markets, which can be seen as an effective adaptation measure (Baldos and Hertel, 2015; Costinot et al., 2016; Hertel and Baldos, 2016; Gouel and Laborde, 2021; WGII Chapter 5.11).. But there is also clear evidence that international trade and the global economy can enhance price volatility, lead to food price spikes and affect food security due to climate and other shocks, as seen recently due to the COVID-19 pandemic (WGII Chapter 5.12, Cottrell et al., 2019; WFP-FSIN, 2020; Verschuur et al., 2021). The continued strong demand for food and other bio-based products, mainly from high- and middle-income countries, therefore, requires better cooperation between nations and global governance of trade to more accurately reflect and disincentivise their environmental and social externalities. Trade in agricultural and extractive products driving land use change in tropical forest and savanna biomes is of major concern because of the biodiversity impacts and GHG emissions incurred in their provision (WGII Cross-Chapter Paper 7, Hosonuma et al., 2012; Forest Trends, 2014; Henders et al., 2015; Curtis et al., 2018; Pendrill et al., 2019; Seymour and Harris, 2019; Kissinger et al., 2021).
In summary, there is significant scope for optimising use of land resources to produce more biomass while reducing adverse effects (high confidence). Context-specific prioritisation, technology innovation in bio-based production, integrative policies, coordinated institutions and improved governance mechanisms to enhance synergies and minimise trade-offs can mitigate the pressure on managed as well as natural and semi-natural ecosystems (medium confidence). Yet, energy conservation and efficiency measures, and deployment of technologies and systems that do not rely on carbon-based energy and materials, are essential for mitigating biomass demand growth as countries pursue ambitious climate goals (high confidence).
Cross-Working Group Box BIOECONOMY
1 5 For lack of space, the focus is on land only, although the bioeconomy also includes sea-related bioresources.
Frequently Asked Questions
FAQ 5.1: How is climate change (already) affecting people’s ability to have enough nutritious food?
Climate change has already made feeding the world’s people more difficult. Climate-related hazards have become more common, disrupting the supply of crops, meat and fish. Rapid changes in weather patterns have put financial strain on producers, while also raising prices and limiting the choices and quality of produce available to consumers.
Most of our food comes from crops, livestock, aquaculture and fisheries. Global food supply increased dramatically in the last century, but ongoing climate change has begun to slow that growth, reducing the gains that would have been expected without climate change. Regionally, negative effects are apparent in regions closer to the equator, with some positive effects further north and south.
Climate impacts are also negatively affecting the quality of produce, from changes in micronutrient content to texture, colour and taste changes that reduce marketability. With warmer and more humid condition, many food pests thrive, food decays more quickly, and food contains more toxic compounds produced by fungi and bacteria.
Warming of the oceans has reduced potential fish catch. The increased carbon dioxide in the atmosphere has led to ocean acidification, which is already impacting the production of farmed fish and shellfish. Changes in local climate have forced producers to shift to new locations, changing what they grow or where they work (e.g., pole-ward shifting fishing grounds).
Climate hazards have increased over the past 50 years and are the major cause of sudden losses of production (food production shocks). Food shocks occur following droughts, heatwaves, floods, storms and outbreaks of climate-related pests and combine to cause multiplying impacts. Climate hazards sometimes disrupt food storage and transport, which impairs the food supply.
All of these negative impacts can lead to increased food prices, and reduced income for producers and retailers as there are fewer products to sell. Together, these impacts threaten to reduce the supply of varied, nutrient-rich foods to poor populations that already suffer ill health.
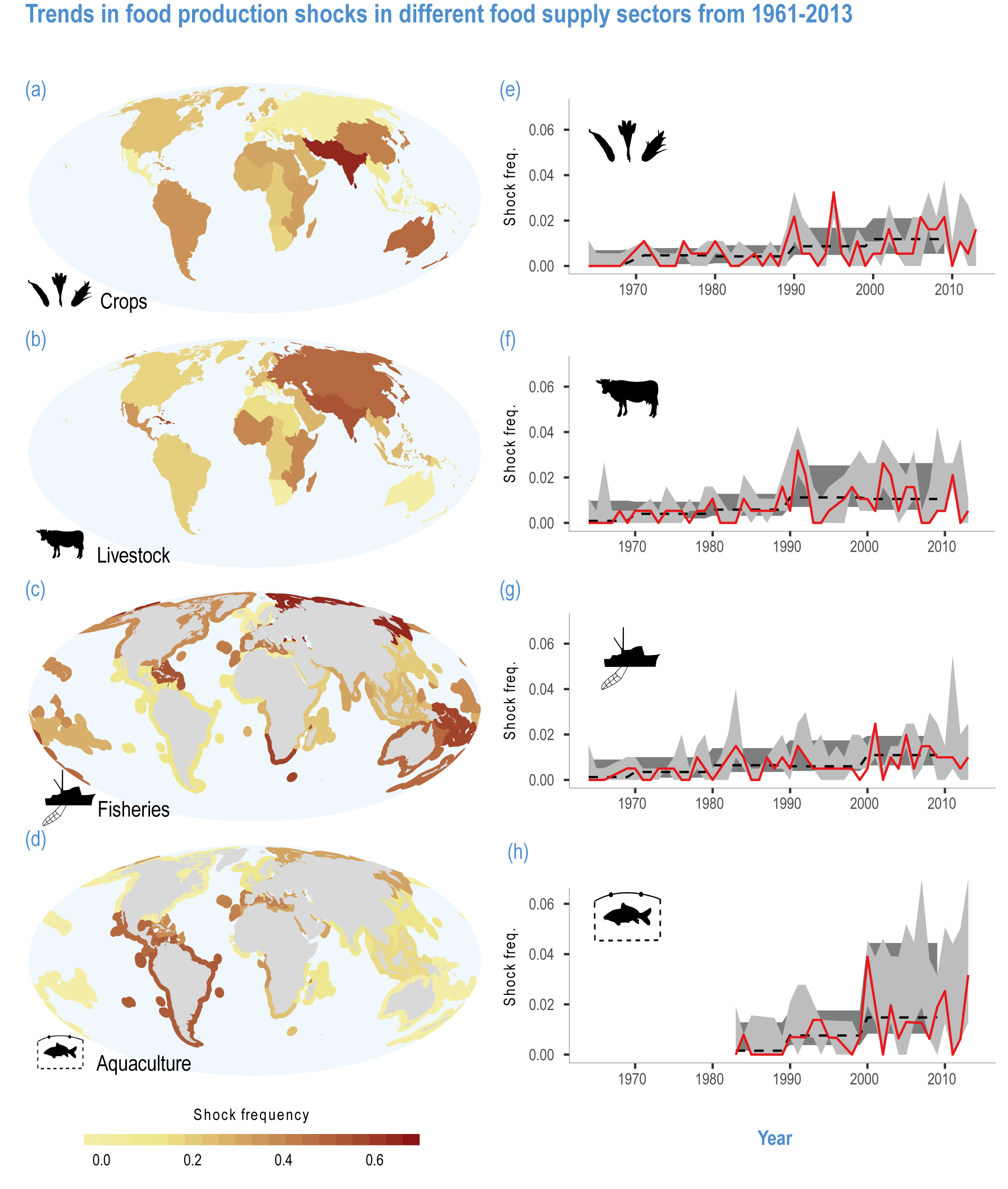
Figure FAQ5.1.1 | Trends in food production shocks in different food supply sectors from 1961 to 2013 (Cottrell et al., 2019). The red lines in the time series are the annual shock frequency, and the dashed line is the decadal mean.
FAQ 5.2: How will climate change impact food availability by mid and late century and who will suffer most?
Climate change impacts will worsen over time, with the period after mid-century seeing more rapid growth in negative impact than in the early part of this century. The impacts will be global, but people with fewer resources, and those who live in regions where impacts will worsen more rapidly, will be hurt the most.
Climate change impacts will worsen over time, but the extent depends on how rapidly greenhouse-gas emissions grow. If the current rate of emissions continues, the impacts will worsen, especially after mid-century, with rapid growth in the number and severity of extreme weather events. Yields of plants, animals and aquaculture will decline in most places, and marine and inland fisheries will suffer. Food production in some regions will become impossible, either because the crops or livestock there cannot survive in the new climatic conditions, or it is too hot and humid for farm workers to be in the fields.
After harvest, agricultural production passes through the agricultural value chain, supplying animal feeds, industrial uses and international markets, with some stored for use in the future. Each of these transitions will be affected by climate change. Food storage facilities will face more challenges in dealing with spoilage. Transportation of perishable fruits, vegetables and meats will become costlier to maintain quality. Households and food services will need to spend more on food preservation.
Low-income countries and poor people are at higher risk, as they have limited social safety nets and suffer more from rising food prices and an unstable food supply. But large famers will also be hurt. Rural communities, especially smallholder farmers, pastoralists and fishers, are extremely vulnerable because their livelihoods mainly depend on their production. The urban poor will have to spend more on food.
A flood, for example, may force low-income families out of their homes, affect their employment and reduce their access to food supplies, with prices often rising after natural disasters. Families will have less access to safe water supplies, and this combination of lower food supplies, uncertain employment, displacement from home and rising food costs will increase the number of children who are undernourished.
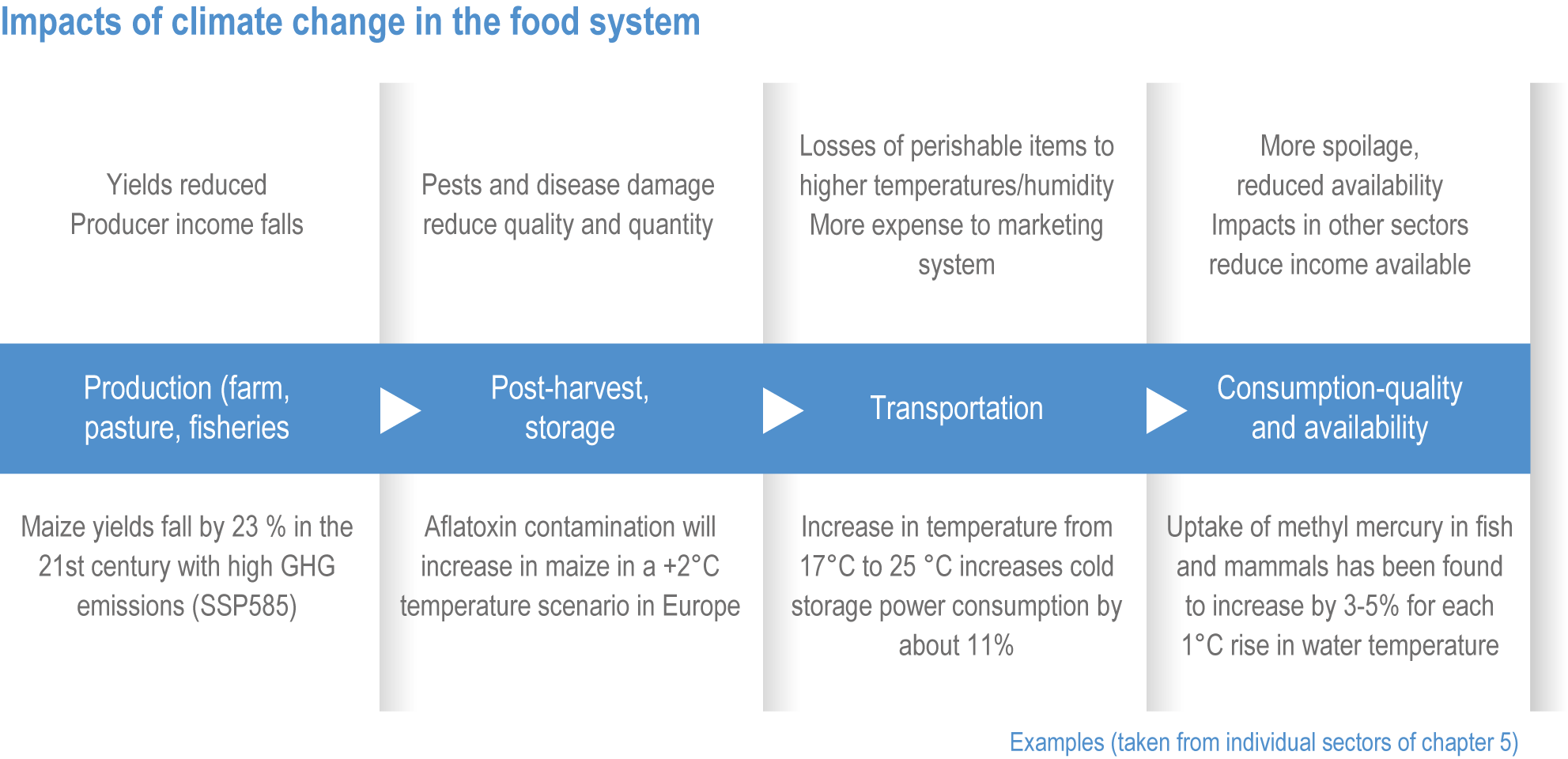
Figure FAQ5.2.1 | Impacts of climate change on the food system.
FAQ 5.3: Land is going to be an important resource for mitigating climate change: how is the increasing competition for land threatening global food security and who will be affected the most?
Climate change will affect food production. Meeting future food needs requires greater land shares unless we change what we eat and how we grow food. Additionally, large-scale land projects that aim to mitigate climate change will increase land competition. Less land will then be available for food production, increasing food insecurity. People at greater risk from land competition are smallholder farmers, Indigenous Peoples and low-income groups.
Why is land important?
Land is a limited resource on which humans and ecosystems depend on to grow plants, which capture carbon dioxide and release oxygen, and provide food, timber and other products. We also have cultural, recreational and spiritual connections to land.
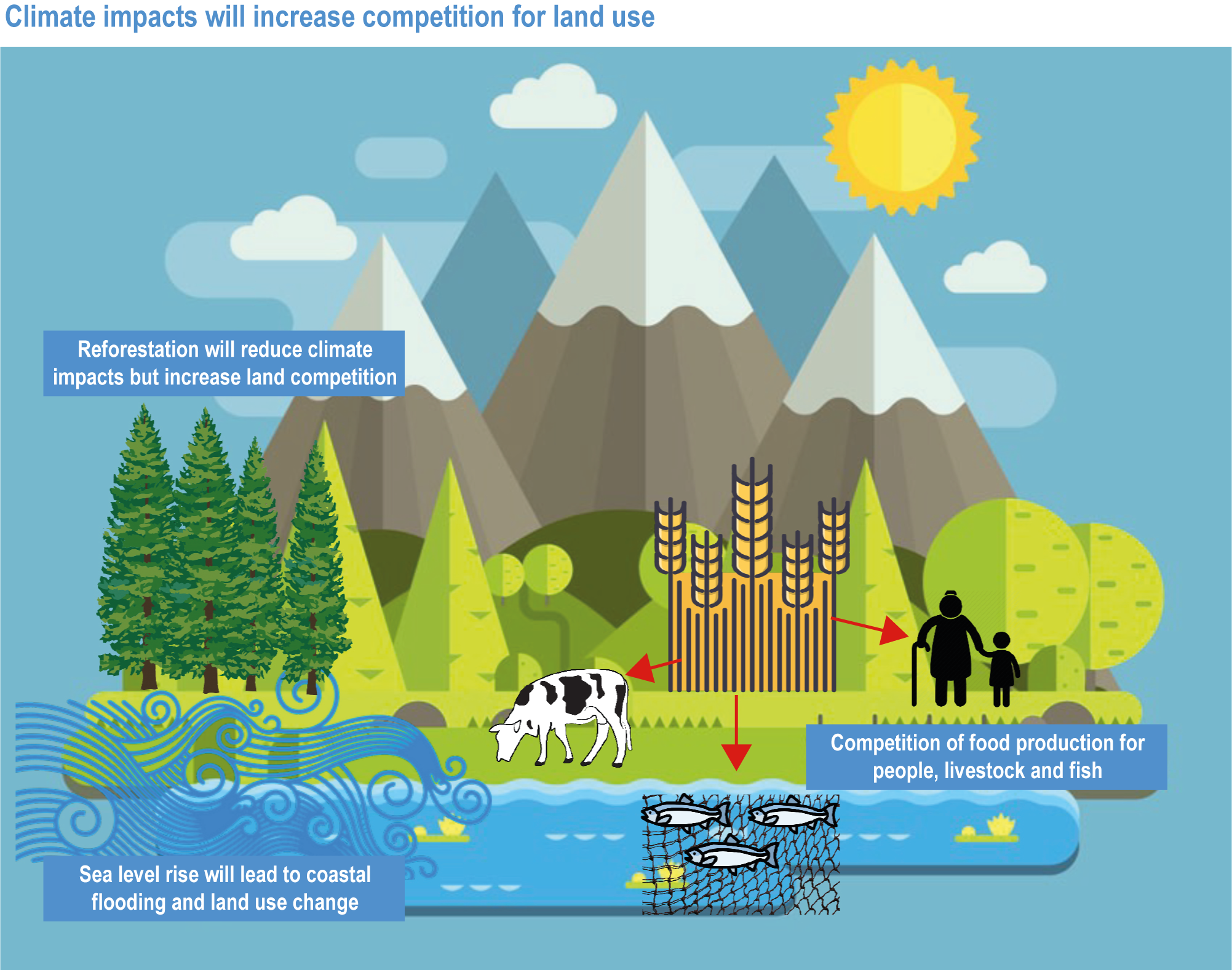
Figure FAQ5.3.1 | Climate impacts will increase competition for land use, reducing coastal land for crops and affecting food security for vulnerable groups. Adaptation methods like coastal aquaculture and mangrove reforestation reduce climate effects but may increase land competition.
Why will climate change affect land use?
Climate change results in more frequent heatwaves, extreme rainfall, drought and rising sea levels, which negatively affect crop yields. More land is thus needed to grow crops, increasing land competition with other food systems that use crops to feed their animals (e.g., livestock, fish). Where land will be flooded, humans cannot grow crops, but food production could be adapted to grow seafood instead. Extensive land allocations aiming at reducing carbon emissions, such as afforestation, reduce land availability for food. Unless carefully managed, competition for land will increase food prices and food security.
Solutions to reduce land competition and protect food security
Sustainable land management allows land to remain productive and support key functions. Other land practices include growing cover crops to improve soil quality. Governments can provide incentives to producers to grow alternative foods and use sustainable practices. Making sure that vulnerable groups (e.g., low-income communities, Indigenous people and small-scale producers) strengthen land tenure rights will help protect food security.
Food by-products used as alternative food sources and other products reduce waste and increase sustainability. Dietary changes are another important solution. People that eat high amounts of meat or unhealthy foods could reduce consumption of these foods and have more diverse diets. These dietary changes will benefit their health and reduce pressure on land. Regulated labelling, education and other policies which encourage healthy diets can support these shifts.
FAQ 5.4: What are effective adaptation strategies for improving food security in a warming world?
A variety of adaptation options exist to improve food security in a warming world. Examples of adaptation for crop production include crop management and livelihood diversification. For livestock-based systems, an example is matching number of animals with the production capacity of pastures. For fisheries, eliminating overfishing is an effective adaptation practice. For mixed cropping and nature-based systems, an appropriate adaptation is agroforestry.
Adaptation strategies to enhance food security vary from farm-level interventions to national policies and international agreements. They cover the following dimensions of food security: availability, access, utilisation (food quality and safety) and stability.
For the production of crops, adaptation strategies include field and farm-level options such as crop management, livelihood diversification and social protection such as crop insurance. The most common field management options are changes in planting schedules, crop varieties, fertilizers and irrigation. For example, farmers can shift their planting schedules in response to the early or late onset of the rainy season. Moreover, there are new crop insurance schemes that are based on changes in weather patterns.
For livestock-based systems, adaptation options include matching the number of animals with the production capacity of pastures; adjusting water management based on seasonal and spatial patterns of forage production; managing animal diet; more effective use of fodder, rotational grazing; fire management to control woody thickening of grass; using more suitable livestock breeds or species; migratory pastoralist activities; and activities to monitor and manage the spread of pests, weeds and diseases.
For ocean and inland fisheries, adaptation options are primarily concentrated in the socioeconomic dimension and governance and management. In general, eliminating overfishing could help rebuild fish stocks, reduce ecosystem impacts, and increase fishing’s adaptive capacity. Aquaculture is often viewed as an adaptation option for fisheries declines. However, there are adaptation strategies specific to aquaculture, including proper species selections at the operational level, such as the cultivation of brackish species (shrimp, crabs) in inland ponds during dry seasons and rice–freshwater finfish in wetter seasons.
For so-called mixed farming systems that produce a combination of crops, livestock, fish and trees, these systems’ inherent diversity provides a solid platform for adaptation. A good example is agroforestry, the purposeful integration of trees or shrubs with crop or livestock systems, which increases resilience against climate risks.
Overall, nature-based systems or ecosystem-based strategies in food systems, such as agroecology, can be a useful adaptation method to increase wild and cultivated food sources. Agroecological practices include agroforestry, intercropping, increasing biodiversity, crop and pasture rotation, adding organic amendments, integration of livestock into mixed systems, cover crops and minimising toxic and synthetic inputs with adverse health and environmental impacts.
FAQ 5.5: Climate change is not the only factor threatening global food security: other than climate action, what other actions are needed to end hunger and ensure access by all people to nutritious and sufficient food all year round?
Our food systems depend on many factors other than climate change, such as food production, water, land, energy and biodiversity. People’s access to healthy food can be also be affected by factors such as poverty and physical insecurity. We are all stakeholders in food systems, whether as producers or consumers, and we can all contribute to the goal of a food-secure world by the choices we make in our everyday lives.
Today more than 820 million people are hungry, and hunger is on the rise in Africa. Two billion people experience moderate or severe food shortages, and another 2 billion suffer from overnutrition, a state of obesity or being overweight from unbalanced diets, with related health impacts such as diabetes and heart disease. The changing climate is already affecting food production. These effects are worsening, affecting food production from crops, livestock, fish and forests in many places where people already do not have enough to eat. Food prices will be affected as a result, with increasing risk that poorer people will not be able to buy enough for their families. Food quality will increasingly be affected too.
Our ability to grow and consume food depends on many factors other than climate change. There are tight connections between food production, water, land, energy and biodiversity, for example. Other factors like gender inequity, poverty, political exclusion, remoteness from urban centres and physical insecurity can all affect people’s access to healthy food.
Food systems are complicated (Figure FAQ5.5.1). To improve food production, supply and distribution, we need to make changes throughout the food supply chain. For instance: improving the way farmers access the inputs needed to grow food; improving the ways in which food is grown, with climate and market information, training and technical know-how, water-saving and water-harvesting technologies; adopting new low-cost and less carbon-intensive storage and processing methods; and creating local networks of producers and processors For food consumers, we could consider shifts to different diets that are healthier and make more efficient use of natural resources; depending on context, these could involve rebalancing consumption of meat and highly processed foods, reducing food loss and waste, and preparing food in more energy-efficient ways. Policymakers can enable such actions through appropriate price and trade policies, implementing policies for sustainable and low-emission agriculture, providing safety nets where needed, and empowering women, youth and other socially disadvantaged groups.
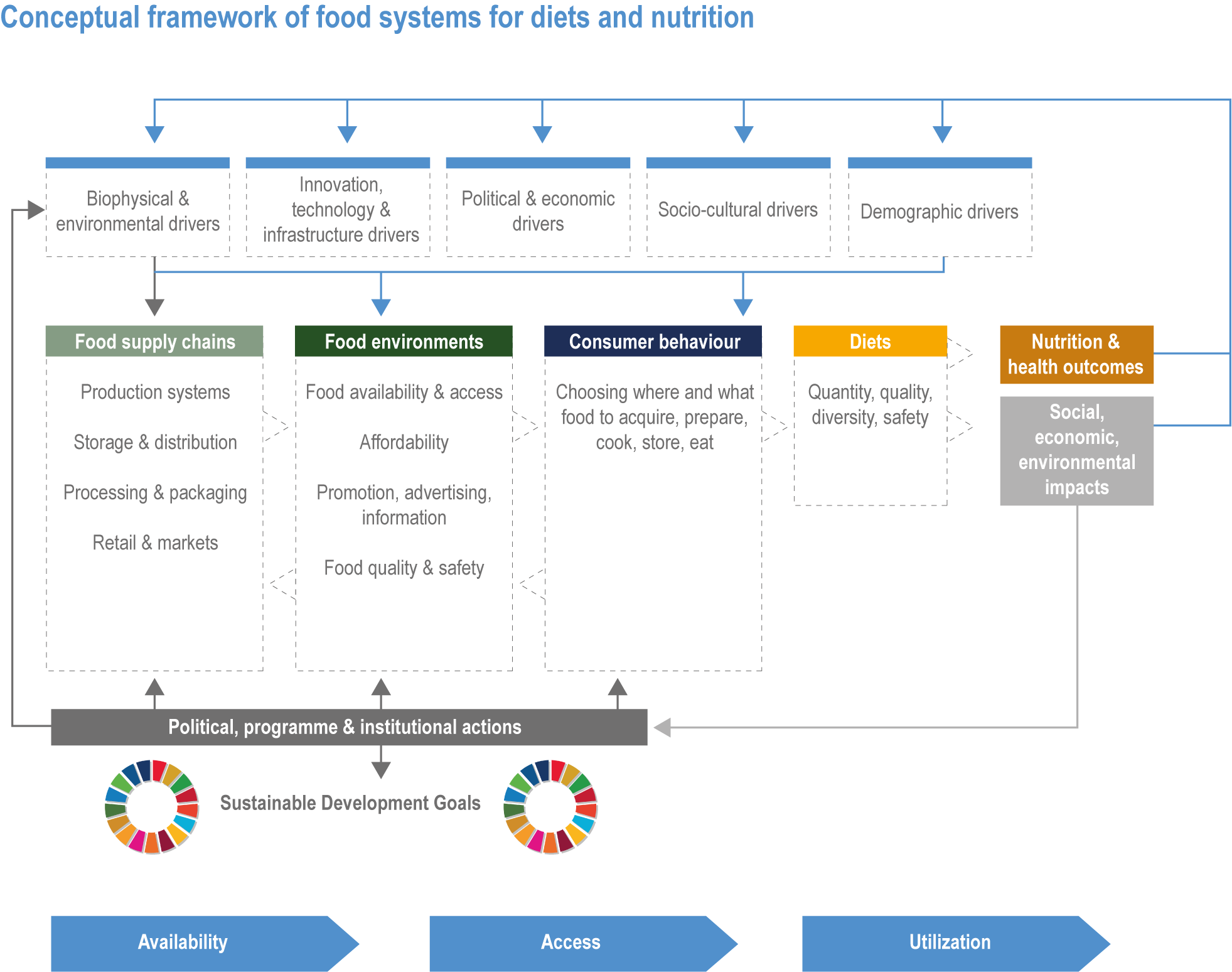
Figure FAQ5.5.1 | Conceptual framework of food systems for diets and nutrition (modified from HLPE, 2017a).
Our food systems need to be robust and sustainable; otherwise we will not be able to manage the additional pressures imposed on them by climate change. We can all contribute to this goal.
References
Aarts, N. and C. Leeuwis, 2010: Participation and power: reflections on the role of government in land use planning and rural development. J. Agric. Educ. Ext. , 16 (2), 131–145, doi:10.1080/13892241003651381.
Lah, A., B.P. Kelaher, D. Bucher and K. Benkendorff, 2018: Ocean warming and acidification affect the nutritional quality of the commercially-harvested turbinid snail Turbo militaris. Mar. Environ. Res. , 141, 100–108, doi:10.1016/j.marenvres.2018.08.009.
Abah, J., P. Mashebe and D.D. Denuga, 2015: Prospect of integrating African indigenous knowledge systems into the teaching of sciences in Africa. Am. J Educ Res, 3 (6), 668–673, doi:10.12691/education-3-6-1.
Abay, K. A., et al., 2021: Landscapes of opportunity: patterns of young people’s engagement with the rural economy in sub-Saharan. Africa. j Dev.stud. , 57 (4), 594–613, doi:10.1080/00220388.2020.1808195.
Abbott, M., M. Bazilian, D. Egel and H.H. Willis, 2017: Examining the food-energy-water and conflict nexus. Curr. Opin. Chem. Eng. , 18, 55–60, doi:10.1016/j.coche.2017.10.002.
Abdel-Tawwab, M., M.N. Monier, S.H. Hoseinifar and C. Faggio, 2019: Fish response to hypoxia stress: growth, physiological, and immunological biomarkers. Fish Physiol Biochem, 45 (3), 997–1013, doi:10.1007/s10695-019-00614-9.
Abdelrahman, H., et al., 2017: Aquaculture genomics, genetics and breeding in the United States: current status, challenges, and priorities for future research. BMC Genomics, 18 (1), 191, doi:10.1186/s12864-017-3557-1.
Abdulai, I., et al., 2018: Cocoa agroforestry is less resilient to sub-optimal and extreme climate than cocoa in full sun. Glob Chang Biol, 24 (1), 273–286, doi:10.1111/gcb.13885.
Aberman, N.-L. and C. Tirado, 2014: Impacts of Climate Change on Food Utilization. Springer, the Netherlands, Dordrecht, 717–724.
Abolmaali, S.M.-R., M. Tarkesh and H. Bashari, 2018: MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inform. , 43, 116–123, doi:10.1016/j.ecoinf.2017.10.002.
Abraham, B., et al., 2014: The system of crop intensification: reports from the field on improving agricultural production, food security, and resilience to climate change for multiple crops. Agric. Food Secur. , 3, 4, doi:10.1186/2048-7010-3-4.
Abtew, A.A., J. Pretzsch, L. Secco and T.E. Mohamod, 2014: Contribution of small-scale gum and resin commercialization to local livelihood and rural economic development in the drylands of eastern Africa. Forests, 5 (5), 952–977, doi:10.3390/f5050952.
Abu Samah, A., H.A.M. Shaffril, A. Hamzah and B. Abu Samah, 2019: Factors affecting small-scale fishermen’s adaptation toward the impacts of climate change: Reflections from Malaysian fishers. SAGE Open, 9 (3), doi:10.1177/2158244019864204.
Acevedo, M., et al., 2020: A scoping review of adoption of climate-resilient crops by small-scale producers in low- and middle-income countries. Nat. Plants, 6 (10), 1231–1241, doi:10.1038/s41477-020-00783-z.
Adams, E.A., E.D. Kuusaana, A. Ahmed and B.B. Campion, 2019: Land dispossessions and water appropriations: Political ecology of land and water grabs in Ghana. Land Use Policy, (June 2018), 87, doi:10.1016/j.landusepol.2019.104068.
Adams, M. A., et al., 2018: Crops, nitrogen, water: Are legumes friend, foe, or misunderstood ally?Trends Plant Sci. , 23 (6), 539–550, doi:10.1016/j.tplants.2018.02.009.
Addison, L. and M. Schnurr, 2016: Growing burdens? Disease-resistant genetically modified bananas and the potential gendered implications for labor in Uganda. Agric Hum Values, 33, 967–978, doi:10.1007/s10460-015-9655-2.
Adhikari, L., A. Hussain and G. Rasul, 2017: Tapping the potential of neglected and underutilized food crops for sustainable nutrition security in the mountains of Pakistan and Nepal. Sustainability, 9 (2), 291, doi:10.3390/su9020291.
Adhikari, P., et al., 2016: Simulating future climate change impacts on seed cotton yield in the Texas High Plains using the CSM-CROPGRO-Cotton model. Agric. Water Manag. , 164, 317–330, doi:10.1016/j.agwat.2015.10.011.
Adhikari, U., A.P. Nejadhashemi and S.A. Woznicki, 2015: Climate change and eastern Africa: a review of impact on major crops. Food Energy Secur. , 4 (2), 110–132, doi:10.1002/fes3.61.
Afidchao, M.M., et al., 2014: Analysing the farm level economic impact of GM corn in the Philippines. NJAS-wageningen J. Life Sci. , 70-71, 113–121, doi:10.1016/j.njas.2014.05.008.
Aggarwal, A., 2014: How sustainable are forestry clean development mechanism projects?—A review of the selected projects from India. Mitig Adapt Strateg Glob Change, 19 (1), 73–91, doi:10.1007/s11027-012-9427-x.
Aggarwal, P., S. Vyas, P. Thornton and B.M. Campbell, 2019: How much does climate change add to the challenge of feeding the planet this century?Environ. Res. Lett. , 14 (4), 43001, doi:10.1088/1748-9326/aafa3e.
Agnolucci, P. and V. De Lipsis, 2020: Long-run trend in agricultural yield and climatic factors in Europe. Clim Change, 159 (3), 385–405, doi:10.1007/s10584-019-02622-3.
Agrawal, A., A. Chhatre and E.R. Gerber, 2015: Motivational crowding in sustainable development interventions. Am. Polit. Sci. Rev. , 109 (3), 470–487, doi:10.1017/S0003055415000209.
Ágreda, T., et al., 2015: Increased evapotranspiration demand in a Mediterranean climate might cause a decline in fungal yields under global warming. Glob. Chang. Biol. , 21 (9), 3499–3510, doi:10.1111/gcb.12960.
Agyekumhene, C., et al., 2018: Digital platforms for smallholder credit access: The mediation of trust for cooperation in maize value chain financing. NJAS Wagening. J. Life. Sci. , 86-87, 77–88, doi:10.1016/j.njas.2018.06.001.
Aha, B. and J.Z. Ayitey, 2017: Biofuels and the hazards of land grabbing: Tenure (in)security and indigenous farmers’ investment decisions in Ghana. Land Use Policy, 60, 48–59, doi:10.1016/j.landusepol.2016.10.012.
Ahmed, B.M.S., et al., 2017: Cows exposed to heat stress during fetal life exhibit improved thermal tolerance. J. Anim. Sci. , 95 (8), 3497–3497, doi:10.2527/jas2016.1298.
Ahmed, I., et al., 2018: Assessing the impact of climate variability on maize using simulation modeling under semi-arid environment of Punjab, Pakistan. Environ. Sci. Pollut. Res. Int. , 25 (28), 28413–28430, doi:10.1007/s11356-018-2884-3.
Ahmed, N., S. Thompson and M. Glaser, 2019: Global aquaculture productivity, environmental sustainability, and climate change adaptability. Environ Manage, 63 (2), 159–172, doi:10.1007/s00267-018-1117-3.
Ahmed, S. and J.R. Stepp, 2016: Beyond yields: climate change effects on specialty crop quality and agroecological management. Elem. Sci. Anthropocene, 4, 1–16, doi:10.12952/journal.elementa.000092.
Ahsan, N. and D. Mitra, 2016: Can the Whole be Greater than the Sum of Its Parts? Lessons from India’s Growing Economy and its Evolving Structure. International Food Policy Research Institute, https://ebrary.ifpri.org/digital/collection/p15738coll2/id/131168. (38--46 pp).
Ainsworth, E.A. and S.P. Long, 2021: 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation?Glob Chang Biol, 27 (1), 27–49, doi:10.1111/gcb.15375.
Aitken, S.N. and J.B. Bemmels, 2016: Time to get moving: assisted gene flow of forest trees. Evol Appl, 9 (1), 271–290, doi:10.1111/eva.12293.
Ajates Gonzalez, R., J. Thomas and M. Chang, 2018: Translating agroecology into policy: The case of France and the United Kingdom. Sustainability, 10 (8), 2930, doi:10.3390/su10082930.
Akil, L., H.A. Ahmad and R.S. Reddy, 2014: Effects of climate change on Salmonella infections. Foodborne Pathog. Dis. , 11 (12), 974–980, doi:10.1089/fpd.2014.1802.
Aksnes, D.L., et al., 2019: Multi-decadal warming of Atlantic water and associated decline of dissolved oxygen in a deep fjord. Estuar. Coast. Shelf Sci. , 228, 106392, doi:10.1016/j.ecss.2019.106392.
Akter, R., et al., 2019: The dominant climate change event for salinity intrusion in the GBM delta. Climate, 7 (5), 69, doi:10.3390/cli7050069.
Al-Gabbiesh, A., M. Kleinwächter and D. Selmar, 2015: Influencing the contents of secondary metabolites in spice and medicinal plants by deliberately applying drought stress during their cultivation. Jordan J. Biol. Sci. , 8 (1), 1–10, doi:10.12816/0026941.
Al-Kodmany, K., 2018: The vertical farm: A review of developments and implications for the vertical city. Buildings, 8 (2), 24, doi:10.3390/buildings8020024.
Alae-Carew, C., et al., 2020: The impact of environmental changes on the yield and nutritional quality of fruits, nuts and seeds: a systematic review. Environ. Res. Lett. , 15 (2), 23002, doi:10.1088/1748-9326/ab5cc0.
Alam, A. and P. Dwivedi, 2019: Modeling site suitability and production potential of carinata-based sustainable jet fuel in the southeastern United States. J Clean Prod, 239, 117817–117817, doi:10.1016/J.JCLEPRO.2019.117817.
Alava, J.J., W.W.L. Cheung, P.S. Ross and U.R. Sumaila, 2017: Climate change-contaminant interactions in marine food webs: toward a conceptual framework. Glob Chang Biol, 23 (10), 3984–4001, doi:10.1111/gcb.13667.
Alba-Sánchez, F., J.A. López-Sáez, D. Nieto-Lugilde and J.-C. Svenning, 2015: Long-term climate forcings to assess vulnerability in North Africa dry argan woodlands. Appl. Veg. Sci. , 18 (2), 283–296, doi:10.1111/avsc.12133.
Albert, C., J.H. Spangenberg and B. Schroter, 2017: Nature-based solutions: criteria. Nature, 543 (7645), 315, doi:10.1038/543315b.
Albert, M., R.V. Nagel, J. Sutmoller and M. Schmidt, 2018: Quantifying the effect of persistent dryer climates on forest productivity and implications for forest planning: a case study in northern Germany. For. Ecosyst. , 5, 33, doi:10.1186/s40663-018-0152-0.
Albizua, A., E. Corbera and U. Pascual, 2019: Farmers’ vulnerability to global change in Navarre, Spain: large-scale irrigation as maladaptation. Reg Environ Change, 19 (4), 1147–1158, doi:10.1007/s10113-019-01462-2.
Albon, S.D., et al., 2017: Contrasting effects of summer and winter warming on body mass explain population dynamics in a food-limited Arctic herbivore. Glob Chang Biol, 23 (4), 1374–1389, doi:10.1111/gcb.13435.
Albrich, K., W. Rammer and R. Seidl, 2020: Climate change causes critical transitions and irreversible alterations of mountain forests. Glob Chang Biol, 26 (7), 4013–4027, doi:10.1111/gcb.15118.
Albrich, K., W. Rammer, D. Thom and R. Seidl, 2018: Trade-offs between temporal stability and level of forest ecosystem services provisioning under climate change. Ecol Appl, 28 (7), 1884–1896, doi:10.1002/eap.1785.
Alcon, F., et al., 2020: Valuing diversification benefits through intercropping in Mediterranean agroecosystems: A choice experiment approach. Ecol. Econ. , 171, 106593–106593, doi:10.1016/j.ecolecon.2020.106593.
Alday, J.G., et al., 2017: Record breaking mushroom yields in Spain. Fungal Ecol, 26, 144–146, doi:10.1016/j.funeco.2017.01.004.
Alexander, P., et al., 2017: Could consumption of insects, cultured meat or imitation meat reduce global agricultural land use?Glob. Food Secur. , 15, 22–32, doi:10.1016/j.gfs.2017.04.001.
Alexander, P., et al., 2015: Drivers for global agricultural land use change: The nexus of diet, population, yield and bioenergy. Glob. Environ. Change. , 35, 138–147, doi:10.1016/j.gloenvcha.2015.08.011.
Algert, S.J., et al., 2016: Vegetable output, cost savings, and nutritional value of low-income families’ home gardens in San Jose, CA. J. Hunger. Environ. Nutr. , 11 (3), 328–336, doi:10.1080/19320248.2015.1128866.
Algur, K.D., S.K. Patel and S. Chauhan, 2021: The impact of drought on the health and livelihoods of women and children in India: A systematic review. Child Youth Serv Rev, 122 (C), S190740920323318.
Ali, A. and O. Erenstein, 2017: Assessing farmer use of climate change adaptation practices and impacts on food security and poverty in Pakistan. Clim. Risk Manag. , 16, 183–194, doi:10.1016/j.crm.2016.12.001.
Ali, A. and D.B. Rahut, 2018: Farmers willingness to grow GM food and cash crops: empirical evidence from Pakistan. GM Crop. Food-Biotechnol Agric. Food Chain, 9 (4), 199–210, doi:10.1080/21645698.2018.1544831.
Alikadic, A., et al., 2019: The impact of climate change on grapevine phenology and the influence of altitude: A regional study. Agric For Meteorol, 271, 73–82, doi:10.1016/j.agrformet.2019.02.030.
Allen, M.R., O.P. Dube, W. Solecki, F. Aragón-Durand, W. Cramer, S. Humphreys, M. Kainuma, J. Kala, N. Mahowald, Y. Mulugetta, R. Perez, M. Wairiu, and K. Zickfeld, 2018: Framing and Context. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty[Masson-Delmotte, V., P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P.R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J.B.R. Matthews, Y. Chen, X. Zhou, M.I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, and T. Waterfield (eds.)]. In press.
Alleway, H.K., et al., 2019: The ecosystem services of marine aquaculture: valuing benefits to people and nature. BioScience, 69 (1), 59–68, doi:10.1093/biosci/biy137.
Allison, E.H. and H.R. Bassett, 2015: Climate change in the oceans: human impacts and responses. Science, 350 (6262), 778–782, doi:10.1126/science.aac8721.
ALRahahleh, L., et al., 2018: Effects of CMIP5 projections on volume growth, carbon stock and timber yield in managed Scots pine, Norway spruce and silver birch stands under southern and northern boreal conditions. Forests, 9 (4), 208, doi:10.3390/f9040208.
Alshannaq, A. and J.H. Yu, 2017: Occurrence, toxicity, and analysis of major mycotoxins in food. Int J Environ Res Public Health, 14 (6), 632, doi:10.3390/ijerph14060632.
Alston, M. and B. Akhter, 2016: Gender and food security in Bangladesh: the impact of climate change. Gend. Place Cult. , 23 (10), 1450–1464, doi:10.1080/0966369x.2016.1204997.
Altieri, M. A., C.I. Nicholls, A. Henao and M. A. Lana, 2015: Agroecology and the design of climate change-resilient farming systems. Agron. Sustain. Dev. , 35 (3), 869–890, doi:10.1007/s13593-015-0285-2.
Altrichter, M., 2006: Wildlife in the life of local people of the semi-arid Argentine Chaco. Biodivers Conserv, 15 (8), 2719–2736, doi:10.1007/s10531-005-0307-5.
Amadou, I., M. Soulé and A. Salé, 2020: An overview on the importance of acacia nilotica (L.) willd. ex del.: A review. Asian J. Res. Agric. For. , 12–18, doi:10.9734/ajraf/2020/v5i330085.
Amadu, F.O., D.C. Miller and P.E. McNamara, 2020: Agroforestry as a pathway to agricultural yield impacts in climate-smart agriculture investments: evidence from southern Malawi. Ecol. Econ. , 167, 106443, doi:10.1016/j.ecolecon.2019.106443.
Amjath-Babu, T.S., et al., 2016: Climate change and indicators of probable shifts in the consumption portfolios of dryland farmers in Sub-Saharan Africa: implications for policy. Ecol Indic, 67, 830–838, doi:10.1016/j.ecolind.2016.03.030.
Amjath-Babu, T.S., et al., 2019: Integrated modelling of the impacts of hydropower projects on the water-food-energy nexus in a transboundary Himalayan river basin. Appl Energy, 239, 494–503, doi:10.1016/j.apenergy.2019.01.147.
Ammer, C., 2019: Diversity and forest productivity in a changing climate. New Phytol. , 221 (1), 50–66, doi:10.1111/nph.15263.
An, R., M. Ji and S. Zhang, 2018: Global warming and obesity: a systematic review. Obes. Rev. , 19 (2), 150–163, doi:10.1111/obr.12624.
Anders, E.J., L.C. Zulu and E.R. Jambo, 2020: Limits to grain-legume technology integration by smallholder farmers: The case of time-sensitive labor demands and food security primacy in Malawi. Agric Syst , 184, 102879, doi:10.1016/j.agsy.2020.102879.
Anderson, C.R., et al., 2019a: From Transition to Domains of Transformation: Getting to Sustainable and Just Food Systems through Agroecology. Sustainability, 11 (19), 5272–5272, doi:10.3390/su11195272.
Anderson, D., J.D. Ford and R.G. Way, 2018: The impacts of climate and social changes on cloudberry (bakeapple) picking: a case study from southeastern Labrador. Hum. Ecol. Interdiscip. J. , 46 (6), 849–863, doi:10.1007/s10745-018-0038-3.
Anderson, W., C. Johansen and K.H.M. Siddique, 2016: Addressing the yield gap in rainfed crops: a review. Agron. Sustain. Dev. , 36 (1), doi:10.1007/s13593-015-0341-y.
Anderson, W.B., et al., 2019b: Synchronous crop failures and climate-forced production variability. Sci. Adv. , 5 (7), eaaw1976, doi:10.1126/sciadv.aaw1976.
Andersson Djurfeldt, A., A. Kalindi, K. Lindsjö and M. Wamulume, 2019: Yearning to farm – Youth, agricultural intensification and land in Mkushi, Zambia. J Rural Stud, 71, 85–93, doi:10.1016/j.jrurstud.2019.08.010.
Andersson, E., E.C.H. Keskitalo and A. Lawrence, 2017: Adaptation to climate change in forestry: a perspective on forest ownership and adaptation responses. Forests, 8 (12), 493, doi:10.3390/f8120493.
Andreotti, F., et al., 2018: Exploring management strategies to enhance the provision of ecosystem services in complex smallholder agroforestry systems. Ecol Indic, 94, 257–265, doi:10.1016/j.ecolind.2018.06.048.
Angelsen, A., et al., 2014: Environmental income and rural livelihoods: A global-comparative analysis. World Dev, 64 (Suppl 1), 12–S28, doi:10.1016/j.worlddev.2014.03.006.
Anisimov, O., et al., 2001: Polar regions (arctic and antarctic). Clim. Chang. , 801–841.
Ankrah, J., 2018: Climate change impacts and coastal livelihoods; an analysis of fishers of coastal Winneba, Ghana. Ocean. Coast. Manag. , 161, 141–146, doi:10.1016/j.ocecoaman.2018.04.029.
Antle, J.M., et al., 2018: Using AgMIP regional integrated assessment methods to evaluate vulnerability, resilience and adaptive capacity for climate smart agricultural systems. In: Climate Smart Agriculture: Building Resilience to Climate Change[Lipper, L., N. McCarthy, D. Zilberman, S. Asfaw and G. Branca(eds.)]. Springer, Cham, Switzerland, pp. 307–333. ISBN 978-3319611938.
Antle, J.M., et al., 2017: Design and use of representative agricultural pathways for integrated assessment of climate change in US Pacific Northwest cereal-based systems. Front. Ecol. Evol. , 5, 99, doi:10.3389/fevo.2017.00099.
Antonelli, C., et al., 2020: Climate impacts on nutrition and labor supply disentangled – an analysis for rural areas of Uganda. Environ Dev Econ, 1–26, doi:10.1017/s1355770x20000017.
Anttila-Hughes, J., A. Jina and G. McCord, 2021: ENSO Impacts Child Undernutrition in the Global Tropics. Nat Commun. 12 (1), 5785, doi:10.1038/s41467-021-26048-7.
Antwi-Agyei, P., A.J. Dougill, L.C. Stringer and S.N.A. Codjoe, 2018: Adaptation opportunities and maladaptive outcomes in climate vulnerability hotspots of northern Ghana. Clim. Risk Manag. , 19, 83–93, doi:10.1016/j.crm.2017.11.003.
Aoki, K., et al., 2019: Exceptional red-tide of fish-killing dinoflagellate Karenia mikimotoi promoted by typhoon-induced upwelling. Estuar. Coast. Shelf Sci. , 219, 14–23, doi:10.1016/j.ecss.2019.01.014.
Apablaza, P., et al., 2017: Primary isolation and characterization of Tenacibaculum maritimum from Chilean Atlantic salmon mortalities associated with a Pseudochattonella spp. algal bloom. J. Aquat. Anim. Health. , 29 (3), 143–149, doi:10.1080/08997659.2017.1339643.
Applequist, W.L., et al., 2020: Scientists’ warning on climate change and medicinal plants. Planta Med, 86 (1), 10–18, doi:10.1055/a-1041-3406.
Apuri, I., K. Peprah and G.T.W. Achana, 2018: Climate change adaptation through agroforestry: the case of Kassena Nankana West District, Ghana. Environ. Dev. , 28, 32–41, doi:10.1016/j.envdev.2018.09.002.
Aquilue, N., et al., 2020: Evaluating forest resilience to global threats using functional response traits and network properties. Ecol Appl, 30 (5), e2095, doi:10.1002/eap.2095.
Araújo, S.S., et al., 2015: Abiotic stress responses in legumes: strategies used to cope with environmental challenges. Crit. Rev. Plant Sci. , 34 (1-3), 237–280, doi:10.1080/07352689.2014.898450.
Araus, J.L. and S.C. Kefauver, 2018: Breeding to adapt agriculture to climate change: affordable phenotyping solutions. Curr. Opin. Plant Biol. , 45 (Pt B), 237–247, doi:10.1016/j.pbi.2018.05.003.
Ardestani, E. G. and Z. Heidari Ghahfarrokhi, 2021: Ensembpecies distribution modeling of Salvia hydrangea under future climate change scenarios in Central Zagros Mountains, Iran. Glob. Ecol. Conserv. , 26, e1488, doi:10.1016/j.gecco.2021.e01488.
Arenas-Sanchez, A., et al., 2019: Effects of increased temperature, drought, and an insecticide on freshwater zooplankton communities. Environ. Toxicol. Chem. , 38 (2), 396–411, doi:10.1002/etc.4304.
Arezki, R., K. Deininger and H. Selod, 2015: What drives the global “land rush”?World Bank Econ. Rev. , 29 (2), 207–233, doi:10.1093/wber/lht034.
Aristizábal-Marulanda, V. and C.A. Cardona Alzate, 2019: Methods for designing and assessing biorefineries: Review. Biofuels Bioprod. Biorefining, 13 (3), 789–808, doi:10.1002/bbb.1961.
Ariti, A.T., J. van Vliet and P.H. Verburg, 2018: Farmers’ participation in the development of land use policies for the Central Rift Valley of Ethiopia. Land Use Policy, 71, 129–137, doi:10.1016/J.LANDUSEPOL.2017.11.051.
Arneth, A., F. Denton, F. Agus, A. Elbehri, K. Erb, B. Osman Elasha, M. Rahimi, M. Rounsevell, A. Spence, R. Valentini, 2019: Framing and Context. In: Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems[P.R. Shukla, J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H.-O. Pörtner, D.C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi, J. Malley, (eds.)], In press, pp. 1–98.
Arora-Jonsson, S., 2011: Virtue and vulnerability: Discourses on women, gender and climate change. Glob. Environ. Change. , 21 (2), 744–751, doi:10.1016/j.gloenvcha.2011.01.005.
Arslan, A., et al., 2015: Climate smart agriculture? Assessing the adaptation implications in Zambia. J. Agric. Econ. , 66 (3), 753–780, doi:10.1111/1477-9552.12107.
Artmann, M. and K. Sartison, 2018: The role of urban agriculture as a nature-based solution: a review for developing a systemic assessment framework. Sustainability, 10 (6), 1937, doi:10.3390/su10061937.
Asase, A. and A.T. Peterson, 2019: Predicted impacts of global climate change on the geographic distribution of an invaluable African medicinal plant resource, Alstonia boonei De Wild. J. Appl. Res. Med. Aromat. Plants, 14, 100206, doi:10.1016/j.jarmap.2019.100206.
Asbjornsen, H., et al., 2014: Targeting perennial vegetation in agricultural landscapes for enhancing ecosystem services. Renew. Agric. Food Syst. , 29 (2), 101–125, doi:10.1017/S1742170512000385.
Asche, F., et al., 2015: Fair enough? Food security and the international trade of seafood. World Dev, 67, 151–160.
Ashley, L., M. Zhumanova, A. Isaeva and C. Dear, 2016: Examining changes in local adaptive capacity resulting from climate change adaptation programming in rural Kyrgyzstan. Clim. Dev. , 8 (3), 281–287, doi:10.1080/17565529.2015.1034230.
Asiedu, B., F.K.E. Nunoo and S. Iddrisu, 2017: Prospects and sustainability of aquaculture development in Ghana, West Africa. Cogent Food Agric, 3 (1), 1349531, doi:10.1080/23311932.2017.1349531.
Asiyanbi, A.P., 2016: A political ecology of REDD plus : property rights, militarised protectionism, and carbonised exclusion in Cross River. Geoforum, 77, 146–156, doi:10.1016/j.geoforum.2016.10.016.
Assan, E., M. Suvedi, L.S. Olabisi and A. Allen, 2018: Coping with and adapting to climate change: a gender perspective from smallholder farming in Ghana. Environments, 5 (8), 86, doi:10.3390/environments5080086.
Asseng, S., et al., 2015: Rising temperatures reduce global wheat production. Nat. Clim. Chang. , 5 (2), 143–147.
Asseng, S., et al., 2013: Uncertainty in simulating wheat yields under climate change. Nat. Clim. Chang. , 3 (9), 827–832, doi:10.1038/Nclimate1916.
Asseng, S., et al., 2019: Climate change impact and adaptation for wheat protein. Glob Chang Biol, 25 (1), 155–173, doi:10.1111/gcb.14481.
Atindana, S.A., et al., 2020: Coping with climate variability and non-climate stressors in the West African Oyster (Crassostrea tulipa) fishery in coastal Ghana. Marit. Stud. , 19 (1), 81–92, doi:10.1007/s40152-019-00132-7.
Atlin, G.N., J.E. Cairns and B. Das, 2017: Rapid breeding and varietal replacement are critical to adaptation of cropping systems in the developing world to climate change. Glob Food Sec, 12, 31–37, doi:10.1016/j.gfs.2017.01.008.
Attridge, S. and L. Engen, 2019: Blended finance in the poorest countries: The need for a better approach. Overseas Development Institute, London, https://cdn.odi.org/media/documents/12666.pdf.
Atukunda, P., et al., 2021: Unlocking the potential for achievement of the UN Sustainable Development Goal 2 – ‘Zero Hunger’ – in Africa: targets, strategies, synergies and challenges. Food Nutr. Res. , 65, doi:10.29219/fnr.v65.7686.
Atuoye, K.N., I. Luginaah, H. Hambati and G. Campbell, 2021: Who are the losers? Gendered-migration, climate change, and the impact of large scale land acquisitions on food security in coastal Tanzania. Land Use Policy, 101 (August 2020), 105154–105154, doi:10.1016/j.landusepol.2020.105154.
Aubry-Kientz, M., et al., 2019: Temperature rising would slow down tropical forest dynamic in the Guiana Shield. Sci Rep, 9 (1), 10235, doi:10.1038/s41598-019-46597-8.
Augustine, D.J., et al., 2018: Elevated CO2 induces substantial and persistent declines in forage quality irrespective of warming in mixedgrass prairie. Ecol Appl, 28 (3), 721–735, doi:10.1002/eap.1680.
Avelino, J., et al., 2015: The coffee rust crises in Colombia and Central America (2008-2013): impacts, plausible causes and proposed solutions. Food Secur, 7 (2), 303–321, doi:10.1007/s12571-015-0446-9.
Avery, S.V., I. Singleton, N. Magan and G.H. Goldman, 2019: The fungal threat to global food security. Fungal Biol, 123 (8), 555–557, doi:10.1016/j.funbio.2019.03.006.
Aveytua-Alcazar, L., D.M. Canu, V.F. Camacho-Ibar and C. Solidoro, 2020: Changes in upwelling regimes in a Mediterranean-type lagoon: a model application. Ecol. Model. , 418, 108908, doi:10.1016/j.ecolmodel.2019.108908.
Avila-Forcada, S., A.L. Martinez-Cruz, R. Rodriguez-Ramirez and E. Sanjurjo-Rivera, 2020: Transitioning to alternative livelihoods: the case of PACE-Vaquita. Ocean. Coast. Manag. , 183, 104984, doi:10.1016/j.ocecoaman.2019.104984.
Avnery, S., D.L. Mauzerall and A.M. Fiore, 2013: Increasing global agricultural production by reducing ozone damages via methane emission controls and ozone-resistant cultivar selection. Glob Chang Biol, 19 (4), 1285–1299, doi:10.1111/gcb.12118.
Avtar, R., K. Tsusaka and S. Herath, 2019: REDD+ implementation in community-based muyong forest management in Ifugao, Philippines. Land, 8 (11), 164, doi:10.3390/land8110164.
Awazi, N.P., M.N. Tchamba and T.M. Avana, 2019: Climate change resiliency choices of small-scale farmers in Cameroon: determinants and policy implications. J Environ Manage, 250, 109560, doi:10.1016/j.jenvman.2019.109560.
Awoye, O.H.R., F. Pollinger, E.K. Agbossou and H. Paeth, 2017: Dynamical-statistical projections of the climate change impact on agricultural production in Benin by means of a cross-validated linear model combined with Bayesian statistics. Agric For Meteorol, 234, 80–94, doi:10.1016/j.agrformet.2016.12.010.
Ayala, L.M., et al., 2016: Impact of agricultural expansion on water footprint in the Amazon under climate change scenarios. Sci. Total Environ. , 569, 1159–1173, doi:10.1016/j.scitotenv.2016.06.191.
Ayers, A.L., J.N. Kittinger and M.B. Vaughan, 2018: Whose right to manage? Distribution of property rights affects equity and power dynamics in comanagement. Ecol. Soc. , 23 (2), 37, doi:10.5751/es-10124-230237.
Azadi, H., F. Taube and F. Taheri, 2018: Co-existence of GM, conventional and organic crops in developing countries: main debates and concerns. Crit. Rev. Food Sci. Nutr. , 58 (16), 2677–2688, doi:10.1080/10408398.2017.1322553.
Büntgen, U., et al., 2012: Drought-induced decline in Mediterranean truffle harvest. Nat. Clim. Chang. , 2 (12), 827–829, doi:10.1038/nclimate1733.
Büntgen, U., et al., 2019: Black truffle winter production depends on Mediterranean summer precipitation. Environ. Res. Lett. , 14 (7), 74004, doi:10.1088/1748-9326/ab1880.
Baarsch, F., et al., 2015: Impacts of Low Aggregate INDCs Ambition: Research commissioned by Oxfam. Climate Analytics.
Baca, M., et al., 2014: An integrated framework for assessing vulnerability to climate change and developing adaptation strategies for coffee growing families in Mesoamerica. PLoS One, 9 (2), e88463, doi:10.1371/journal.pone.0088463.
Bacon, C.M., et al., 2014: Explaining the ‘hungry farmer paradox’: Smallholders and fair trade cooperatives navigate seasonality and change in Nicaragua’s corn and coffee markets. Glob. Environ. Change. , 25, 133–149, doi:10.1016/j.gloenvcha.2014.02.005.
Bacon, C.M., W.A. Sundstrom, I.T. Stewart and D. Beezer, 2017: Vulnerability to cumulative hazards: coping with the coffee leaf rust outbreak, drought, and food insecurity in Nicaragua. World Dev, 93, 136–152, doi:10.1016/j.worlddev.2016.12.025.
Bacon, C.M., et al., 2021: Towards smallholder food and water security: Climate variability in the context of multiple livelihood hazards in Nicaragua. World Dev, 143, 105468–105468, doi:10.1016/j.worlddev.2021.105468.
Bacon, L. and D. Krpan, 2018: (Not) eating for the environment: the impact of restaurant menu design on vegetarian food choice. Appetite, 125, 190–200, doi:10.1016/j.appet.2018.02.006.
Badami, M.G. and N. Ramankutty, 2015: Urban agriculture and food security: a critique based on an assessment of urban land constraints. Glob. Food Secur. Policy Econ. Environ. , 4, 8–15, doi:10.1016/j.gfs.2014.10.003.
Bahadur, A., et al., 2015: The 3 As: Tracking Resilience Across BRACED. London, UK, https://cdn.odi.org/media/documents/9812.pdf. (57-57 pp).
Bahta, Y.T., A. Jordaan and F. Muyambo, 2016: Communal farmers’ perception of drought in South Africa: policy implication for drought risk reduction. Int. J. Disaster Risk Reduct. , 20, 39–50, doi:10.1016/j.ijdrr.2016.10.007.
Bai, Y., et al., 2019: Does climate adaptation of vulnerable households to extreme events benefit livestock production?J Clean Prod, 210, 358–365, doi:10.1016/j.jclepro.2018.10.250.
Baines, J., 2017: Accumulating through food crisis? Farmers, commodity traders and the distributional politics of financialization. Rev. Int. Polit. Econ. , 24 (3), 497–537, doi:10.1080/09692290.2017.1304434.
Baker-Austin, C., J. Trinanes, N. Gonzalez-Escalona and J. Martinez-Urtaza, 2017: Non-cholera vibrios: the microbial barometer of climate change. Trends Microbiol. , 25 (1), 76–84, doi:10.1016/j.tim.2016.09.008.
Baker, R.E. and J. Anttila-Hughes, 2020: Characterizing the contribution of high temperatures to child undernourishment in Sub-Saharan Africa. Sci Rep, 10 (1), 18796–18796, doi:10.1038/s41598-020-74942-9.
Baldos, U.L.C. and T.W. Hertel, 2015: The role of international trade in managing food security risks from climate change. Food Secur, 7 (2), 275–290, doi:10.1007/s12571-015-0435-z.
Balehegn, M., 2015: Unintended Consequences: The Ecological Repercussions of Land Grabbing in Sub-Saharan Africa. Environ. Sci. Policy Sustain. Dev. , 57 (2), 4–21, doi:10.1080/00139157.2015.1001687.
Balehegn, M., S. Balehey, C. Fu and W. Liang, 2019: Indigenous weather and climate forecasting knowledge among Afar pastoralists of north eastern Ethiopia: role in adaptation to weather and climate variability. Pastoralism, 9, 8, doi:10.1186/s13570-019-0143-y.
Balehey, S., G. Tesfay and M. Balehegn, 2018: Traditional gender inequalities limit pastoral women’s opportunities for adaptation to climate change: evidence from the Afar pastoralists of Ethiopia. Pastoralism, 8, 23, doi:10.1186/s13570-018-0129-1.
Ballew, C., A.R. Tzilkowski, K. Hamrick and E.D. Nobmann, 2006: The contribution of subsistence foods to the total diet of Alaska natives in 13 rural communities. Ecol. Food. Nutr. , 45 (1), 1–26, doi:10.1080/03670240500408302.
Balmford, A., et al., 2018: The environmental costs and benefits of high-yield farming. Nat Sustain, 1 (9), 477–485, doi:10.1038/s41893-018-0138-5.
Bandin, I. and S. Souto, 2020: Betanodavirus and VER disease: a 30-year research review. Pathogens, 9 (2), doi:10.3390/pathogens9020106.
Bandyopadhyay, R., et al., 2016: Biological control of aflatoxins in Africa: current status and potential challenges in the face of climate change. World Mycotoxin J. , 9 (5), 771–789, doi:10.3920/wmj2016.2130.
Banerjee, S., et al., 2019: Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J, 13 (7), 1722–1736, doi:10.1038/s41396-019-0383-2.
Bannister, W., et al., 2019: Potential anthropogenic regime shifts in three freshwater lakes in Tropical East Asia. Freshw Biol, 64 (4), 708–722, doi:10.1111/fwb.13256.
Baptiste, A.K. and H. Devonish, 2019: The Manifestation of Climate Injustices: The Post-Hurricane Irma Conflicts Surrounding Barbuda’s Communal Land Tenure. J. Extrem. Events, 06 (01), 1940002, doi:10.1142/S2345737619400025.
Barange, M., 2019: Avoiding misinterpretation of climate change projections of fish catches. ICES J. Mar. Sci. , 76 (6), 1390–1392, doi:10.1093/icesjms/fsz061.
Barange, M. and K. Cochrane, 2018: Impacts of climate change on fisheries and aquaculture: conclusions. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 611–628. ISBN 978-9251306079.
Barange, M., et al., 2014: Impacts of climate change on marine ecosystem production in societies dependent on fisheries. Nat. Clim. Chang. , 4 (3), 211–216, doi:10.1038/Nclimate2119.
Barbarossa, V., et al., 2021: Threats of global warming to the world’s freshwater fishes. Nat Commun, 12 (1), 1–10.
Barbato, M., et al., 2020: Adaptive introgression from indicine cattle into white cattle breeds from Central Italy. Sci Rep, 10 (1), 1–11, doi:10.1038/s41598-020-57880-4.
Barber, I., B.W. Berkhout and Z. Ismail, 2016: Thermal change and the dynamics of multi-host parasite life cycles in aquatic ecosystems. Integr. Comp. Biol. , 56 (4), 561–572, doi:10.1093/icb/icw025.
Barber, Q.E., et al., 2018: Potential impacts of climate change on the habitat of boreal woodland caribou. Ecosphere, 9 (10), e2472, doi:10.1002/ecs2.2472.
Barbier, B., et al., 2009: Human vulnerability to climate variability in the Sahel: farmers’ adaptation strategies in northern Burkina Faso. Environ Manage, 43 (5), 790–803, doi:10.1007/s00267-008-9237-9.
Barbieri, P., S. Pellerin, V. Seufert and T. Nesme, 2019: Changes in crop rotations would impact food production in an organically farmed world. Nat. Sustain. , 2 (5), 378–385, doi:10.1038/s41893-019-0259-5.
Barbosa, V., et al., 2019: Paralytic shellfish toxins and ocean warming: bioaccumulation and ecotoxicological responses in juvenile gilthead seabream (Sparus aurata). Toxins, 11 (7), 408, doi:10.3390/toxins11070408.
Barik, B., et al., 2016: Water-food-energy nexus: changing scenarios in India during recent decades. Hydrol. Earth. Syst. Sci. Discuss. , 21, 3041–3060, doi:10.5194/hess-2016-647.
Barlow, K., et al., 2015: Simulating the impact of extreme heat and frost events on wheat crop production: A review. Field Crop. Res. , 171, 109–119.
Barnuud, N.N., A. Zerihun, M. Gibberd and B. Bates, 2014: Berry composition and climate: responses and empirical models. Int. J. Biometeorol. , 58 (6), 1207–1223, doi:10.1007/s00484-013-0715-2.
Barral, M.P., R. Benayas, P. Meli and N.O. Maceira, 2015: Quantifying the impacts of ecological restoration on biodiversity and ecosystem services in agroecosystems: a global meta-analysis. Agric. Ecosyst. Environ. , 202, 223–231, doi:10.1016/j.agee.2015.01.009.
Barrios, E., et al., 2020: The 10 Elements of Agroecology: enabling transitions towards sustainable agriculture and food systems through visual narratives. Ecosyst. People, 16 (1), 230–247, doi:10.1080/26395916.2020.1808705.
Bartomeus, I., et al., 2011: Climate-associated phenological advances in bee pollinators and bee-pollinated plants. Proc. Natl. Acad. Sci. USA. , 108 (51), 20645–20649, doi:10.1073/pnas.1115559108.
Barton, A., et al., 2015: Impacts of coastal acidification on the Pacific Northwest shellfish industry and adaptation strategies implemented in response. Oceanography, 28 (2), 146–159, doi:10.5670/oceanog.2015.38.
Basche, A.D. and M.S. DeLonge, 2019: Comparing infiltration rates in soils managed with conventional and alternative farming methods: a meta-analysis. PLoS ONE, 14 (9), e215702–e215702, doi:10.1371/journal.pone.0215702.
Bassett, T.J. and M. Koné, 2017: Peanuts for cashews? Agricultural diversification and the limits of adaptability in Côte d’Ivoire. In: Other Geographies:The Influences of Michael Watts[Chari, S., S. Friedberg, V. Gidwani, J. Ribot and W. Wolford(eds.)]. Wiley Blackwell, Hoboken, NJ, USA and Chichester, UK, pp. 79–96. ISBN 978-1119184775.
Basso, B., et al., 2018: Soil organic carbon and nitrogen feedbacks on crop yields under climate change. Agric. Environ. Lett. , 3 (1), 180026, doi:10.2134/ael2018.05.0026.
Battilani, P., 2016: Recent advances in modeling the risk of mycotoxin contamination in crops. Curr. Opin. Food Sci. , 11, 10–15, doi:10.1016/j.cofs.2016.08.009.
Battilani, P., et al., 2016: Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci Rep, 6 (1), 24328, doi:10.1038/srep24328.
Baudron, A.R., et al., 2020: Changing fish distributions challenge the effective management of European fisheries. Ecography, 43 (4), 494–505.
Bava, L., et al., 2019: Rearing of Hermetia illucens on different organic by-products: influence on growth, waste reduction, and environmental impact. Animals, 9 (6), 289, doi:10.3390/ani9060289.
Bayrak, M.M. and L.M. Marafa, 2016: Ten years of REDD plus : a critical review of the impact of REDD plus on forest-dependent communities. Sustainability, 8 (7), 620, doi:10.3390/su8070620.
Beach, R.H., et al., 2015: Climate change impacts on US agriculture and forestry: benefits of global climate stabilization. Environ. Res. Lett. , 10 (9), 95004, doi:10.1088/1748-9326/10/9/095004.
Beach, R.H., et al., 2019: Combining the effects of increased atmospheric carbon dioxide on protein, iron, and zinc availability and projected climate change on global diets: a modelling study. Lancet Planet. Health, 3 (7), e307–e317, doi:10.1016/s2542-5196(19)30094-4.
Beal, T., et al., 2017: Global trends in dietary micronutrient supplies and estimated prevalence of inadequate intakes. PLoS ONE, 12 (4), e175554, doi:10.1371/journal.pone.0175554.
Beaumier, M.C. and J.D. Ford, 2010: Food insecurity among Inuit women exacerbated by socio-economic stresses and climate change. Can. J. Public Health, 101 (3), 196–201, doi:10.1007/Bf03404373.
Bebber, D.P., 2015: Range-expanding pests and pathogens in a warming world. Annu. Rev. Phytopathol. , 53 (1), 335–356, doi:10.1146/annurev-phyto-080614-120207.
Bebber, D.P., A.D. Castillo and S.J. Gurr, 2016: Modelling coffee leaf rust risk in Colombia with climate reanalysis data. Philos. Trans. R. Soc. Lond. B Biol. Sci. , 371 (1709), 20150458, doi:10.1098/rstb.2015.0458.
Bebber, D.P., M.A.T. Ramotowski and S.J. Gurr, 2013: Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. , 3 (11), 985–988, doi:10.1038/Nclimate1990.
Beckford, C.L. and K. Rhiney, 2016: Future of food and agriculture in the Caribbean in the context of climate change and globalization: where do we go from here? In: Globalization, Agriculture and Food in the Caribbean: Climate Change, Gender, and Geography[Beckford, C.L. and K. Rhiney(eds.)]. Palgrave, Macmillian, London, UK, pp. 267–295. ISBN 978-1137538376.
Beckmann, M., et al., 2019: Conventional land-use intensification reduces species richness and increases production: A global meta-analysis. Glob Chang Biol, 25 (6), 1941–1956, doi:10.1111/gcb.14606.
Behringer, D.C., C.L. Wood, M. Krkosek and D. Bushek, 2020: Disease in fisheries and aquaculture. Mar. Dis. Ecol. , 183–209, doi:10.1093/oso/9780198821632.003.0010.
Beillouin, D., T. Ben-Ari and D. Makowski, 2019a: A dataset of meta-analyses on crop diversification at the global scale. Data Br, 24, 103898, doi:10.1016/j.dib.2019.103898.
Beillouin, D., T. Ben-Ari and D. Makowski, 2019b: Evidence map of crop diversification strategies at the global scale. Environ. Res. Lett. , 14 (12), 123001–123001, doi:10.1088/1748-9326/ab4449.
Bélanger, J. and D. Pilling (eds.), 2019: The State of the World’s Biodiversity for Food and Agriculture. FAO Commission on Genetic Resources for Food and Agriculture Assessments, Rome.
Belay, A., J.W. Recha, T. Woldeamanuel and J.F. Morton, 2017: Smallholder farmers’ adaptation to climate change and determinants of their adaptation decisions in the Central Rift Valley of Ethiopia. Agric. Food Secur. , 6 (1), doi:10.1186/s40066-017-0100-1.
Belesova, K., et al., 2019: Drought exposure as a risk factor for child undernutrition in low- and middle-income countries: A systematic review and assessment of empirical evidence. Environ Int , 131, 104973–104973, doi:10.1016/j.envint.2019.104973.
Belhabib, D., et al., 2020: Catching industrial fishing incursions into inshore waters of Africa from space. Fish. Fish. , 21 (2), 379–392, doi:10.1111/faf.12436.
Belhabib, D., V.W.Y. Lam and W.W.L. Cheung, 2016: Overview of West African fisheries under climate change: impacts, vulnerabilities and adaptive responses of the artisanal and industrial sectors. Mar. Policy. , 71, 15–28, doi:10.1016/j.marpol.2016.05.009.
Belhabib, D., U.R. Sumaila and P. Le Billon, 2019: The fisheries of Africa: exploitation, policy, and maritime security trends. Mar. Policy. , 101, 80–92, doi:10.1016/j.marpol.2018.12.021.
Belhabib, D., U.R. Sumaila and D. Pauly, 2015: Feeding the poor: contribution of West African fisheries to employment and food security. Ocean. Coast. Manag. , 111, 72–81, doi:10.1016/j.ocecoaman.2015.04.010.
Bell, D., L. Molloy, J. Prpic and M. Tomko, 2019: A new World Heritage site for Aboriginal engineering. Nature, 572, 32, doi:10.1038/d41586-019-02315-y.
Bell, J.D., et al., 2018: Climate change impacts, vulnerabilities and adaptations: Western and Central Pacific Ocean marine fisheries. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 305–324. ISBN 978-9251306079.
Bell, L.W., A.D. Moore and J.A. Kirkegaard, 2014: Evolution in crop-livestock integration systems that improve farm productivity and environmental performance in Australia. Eur J Agron, 57, 10–20, doi:10.1016/j.eja.2013.04.007.
Bell, R.J., J. Odell, G. Kirchner and S. Lomonico, 2020: Actions to promote and achieve climate-ready fisheries: summary of current practice. Mar. Coast. Fish. , 12 (3), 166–190, doi:10.1002/mcf2.10112.
Bellon, M.R., G.D. Ntandou-Bouzitou and F. Caracciolo, 2016: On-farm diversity and market participation are positively associated with dietary diversity of rural mothers in Southern Benin, West Africa. PLoS ONE, 11 (9), e162535, doi:10.1371/journal.pone.0162535.
Bellon, S. and G. Ollivier, 2018: Institutionalizing agroecology in France: social circulation changes the meaning of an idea. Sustainability, 10 (5), 1380, doi:10.3390/su10051380.
Belote, R.T., J.F. Weltzin and R.J. Norby, 2004: Response of an understory plant community to elevated [CO2 ] depends on differential responses of dominant invasive species and is mediated by soil water availability. New Phytol. , 161 (3), 827–835, doi:10.1111/j.1469-8137.2004.00977.x.
Belton, B., et al., 2020: Farming fish in the sea will not nourish the world. Nat Commun, 11 (1), 5804, doi:10.1038/s41467-020-19679-9.
Béné, C., et al., 2016: Contribution of fisheries and aquaculture to food security and poverty reduction: assessing the current evidence. World Dev, 79, 177–196, doi:10.1016/j.worlddev.2015.11.007.
Bene, C., et al., 2015: Feeding 9 billion by 2050 - putting fish back on the menu. Food Secur, 7 (2), 261–274, doi:10.1007/s12571-015-0427-z.
Bene, C., et al., 2019: Global map and indicators of food system sustainability. Sci Data, 6 (1), 279, doi:10.1038/s41597-019-0301-5.
Bengtsson, J., et al., 2019: Grasslands-more important for ecosystem services than you might think. Ecosphere, 10 (2), e2582–e2582, doi:10.1002/ecs2.2582.
Benito-Garzón, M. and J.F. Fernández-Manjarrés, 2015: Testing scenarios for assisted migration of forest trees in Europe. New For. , 46 (5), 979–994, doi:10.1007/s11056-015-9481-9.
Benjamin, E.O., O. Ola and G. Buchenrieder, 2018: Does an agroforestry scheme with payment for ecosystem services (PES) economically empower women in sub-Saharan Africa?Ecosyst. Serv. , 31, 1–11, doi:10.1016/j.ecoser.2018.03.004.
Bennett, E.L., et al., 2007: Hunting for consensus: reconciling bushmeat harvest, conservation, and development policy in West and Central Africa. Conserv. Biol. , 21 (3), 884–887, doi:10.1111/j.1523-1739.2006.00595.x.
Bennett, N.J., P. Dearden, G. Murray and A. Kadfak, 2014: The capacity to adapt?: communities in a changing climate, environment, and economy on the northern Andaman coast of Thailand. Ecol. Soc. , 19 (2), 5, doi:10.5751/es-06315-190205.
Bennett, N.J., A. Kadfak and P. Dearden, 2016: Community-based scenario planning: a process for vulnerability analysis and adaptation planning to social-ecological change in coastal communities. Environ. Dev. Sustain. , 18 (6), 1771–1799, doi:10.1007/s10668-015-9707-1.
Benton, T.G., et al., 2018: Designing sustainable landuse in a 1.5 degrees C world: the complexities of projecting multiple ecosystem services from land. Curr. Opin. Environ. Sustain. , 31, 88–95, doi:10.1016/j.cosust.2018.01.011.
Berhe, M., et al., 2017: The effects of adaptation to climate change on income of households in rural Ethiopia. Pastoralism, 7 (1), doi:10.1186/s13570-017-0084-2.
Berkes, F., J. Colding and C. Folke (eds.), 2003: Navigating Social-Ecological Systems: Building Resilience for Complexity and Change. Cambridge University Press, Cambridge, UK, ISBN 978-0521815924.
Berner, J., et al., 2016: Adaptation in Arctic circumpolar communities: food and water security in a changing climate. Int. J. Circumpolar Health, 75 (1), 33820, doi:10.3402/ijch.v75.33820.
Berner, S., et al., 2019: Roadmapping to enhance local food supply: case study of a city-region in Austria. Sustainability, 11 (14), 3876, doi:10.3390/su11143876.
Berners-Lee, M., C. Kennelly, R. Watson and C.N. Hewitt, 2018: Current global food production is sufficient to meet human nutritional needs in 2050 provided there is radical societal adaptation. Elem. Sci. Anthropocene, 6 (1), 52, doi:10.1525/elementa.310.
Bernues, A., T. Rodriguez-Ortega, R. Ripoll-Bosch and F. Alfnes, 2014: Socio-cultural and economic valuation of ecosystem services provided by Mediterranean mountain agroecosystems. Plos One, 9 (7), e102479, doi:10.1371/journal.pone.0102479.
Berrang-Ford, L., et al., 2021a: The Global Adaptation Mapping Initiative (GAMI): Part 1 – Introduction and overview of methods. protocolexchange, doi:10.21203/rs.3.pex-1240/v1.
Berrang-Ford, L., et al., 2021b: A systematic global stocktake of evidence on human adaptation to climate change. Nat. Clim. Chang. 11 (11), 989-1000, doi:10.1038/s41558-021-01170-y..
Bett, B., et al., 2017: Effects of climate change on the occurrence and distribution of livestock diseases. Prev. Vet. Med. , 137 (Pt B), 119–129, doi:10.1016/j.prevetmed.2016.11.019.
Bettini, G., S.L. Nash and G. Gioli, 2017: One step forward, two steps back? The fading contours of (in)justice in competing discourses on climate migration. Geogr J, 183 (4), 348–358, doi:10.1111/geoj.12192.
Beveridge, L., S. Whitfield and A. Challinor, 2018a: Crop modelling: towards locally relevant and climate-informed adaptation. Clim Change, 147 (3-4), 475–489, doi:10.1007/s10584-018-2160-z.
Beveridge, M.C.M., et al., 2018b: Climate change and aquaculture: interactions with fisheries and agriculture. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 491–516. ISBN 978-9251306079.
Beyene, T., D. Endalamaw, Y. Tolossa and A. Feyisa, 2015: Evaluation of rational use of veterinary drugs especially antimicrobials and anthelmintics in Bishoftu, Central Ethiopia. BMC Res Notes, 8, 482, doi:10.1186/s13104-015-1466-4.
Bezner Kerr, R., 2014: Lost and Found Crops: Agrobiodiversity, Indigenous Knowledge, and a Feminist Political Ecology of Sorghum and Finger Millet in Northern Malawi. Ann. Assoc. Am. Geogr. , 104 (3), 577–593.
Bezner Kerr, R., et al., 2019: Participatory agroecological research on climate change adaptation improves smallholder farmer household food security and dietary diversity in Malawi. Agric. Ecosyst. Environ. , 279, 109–121, doi:10.1016/j.agee.2019.04.004.
Bezner Kerr, R., et al., 2021: Can agroecology improve food security and nutrition? A review. Glob. Food Secur. , 29, 100540–100540, doi:10.1016/j.gfs.2021.100540.
Bharucha, Z. and J. Pretty, 2010: The roles and values of wild foods in agricultural systems. Philos. Trans. Royal Soc. B: Biol. Sci. , 365 (1554), 2913–2926.
Bharucha, Z.P., S.B. Mitjans and J. Pretty, 2020: Towards redesign at scale through zero budget natural farming in Andhra Pradesh, India. Int. J. Agric. Sustain. , 18 (1), 1–20, doi:10.1080/14735903.2019.1694465.
Bhatt, I.D., S. Rawat, A. Badhani and R.S. Rawal, 2017: Nutraceutical potential of selected wild edible fruits of the Indian Himalayan region. Food Chem, 215, 84–91, doi:10.1016/j.foodchem.2016.07.143.
Bianchi, F., et al., 2018: Restructuring physical micro-environments to reduce the demand for meat: a systematic review and qualitative comparative analysis. Lancet Planet Health, 2 (9), e384–e397, doi:10.1016/S2542-5196(18)30188-8.
Biasi, R., E. Brunori, C. Ferrara and L. Salvati, 2019: Assessing impacts of climate change on phenology and quality traits of Vitis vinifera L.: the contribution of local knowledge. Plants, 8 (5), 121, doi:10.3390/plants8050121.
Biasutti, M., 2019: Rainfall trends in the African Sahel: Characteristics, processes, and causes. Wiley. Interdiscip. Rev. Clim. Change. , 10 (4), e591, doi:10.1002/wcc.591.
Bichet, A. and A. Diedhiou, 2018: West African Sahel has become wetter during the last 30 years, but dry spells are shorter and more frequent. Clim. Res. , 75 (2), 155–162, doi:10.3354/cr01515.
Biewener, C., 2016: Paid work, unpaid work, and economic viability in alternative food initiatives: reflections from three Boston urban agriculture endeavors. J Agric Food Syst Community Dev, 1–19, doi:10.5304/jafscd.2016.062.019.
Bigler, C., M. Amacker, C. Ingabire and E. Birachi, 2017: Rwanda’s gendered agricultural transformation: A mixed-method study on the rural labour market, wage gap and care penalty. Womens Stud Int Forum, 64, 17–27, doi:10.1016/j.wsif.2017.08.004.
Bindoff, N.L., W.W.L. Cheung, J.G. Kairo, J. Arístegui, V.A. Guinder, R. Hallberg, N. Hilmi, N. Jiao, M.S. Karim, L. Levin, S. O’Donoghue, S.R. Purca Cuicapusa, B. Rinkevich, T. Suga, A. Tagliabue, and P. Williamson, 2019: Changing Ocean, Marine Ecosystems, and Dependent Communities. In: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate[H.-O. Pörtner, D.C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegría, M. Nicolai, A. Okem, J. Petzold, B. Rama, N.M. Weyer (eds.)]. In press, pp. 447–587.
Birthal, P.S. and J. Hazrana, 2019: Crop diversification and resilience of agriculture to climatic shocks: evidence from India. Agric Syst , 173, 345–354, doi:10.1016/j.agsy.2019.03.005.
Bisaga, I., P. Parikh and C. Loggia, 2019: Challenges and opportunities for sustainable urban farming in South African low-income settlements: a case study in Durban. Sustainability, 11 (20), 5660, doi:10.3390/su11205660.
Bisbis, M.B., N. Gruda and M. Blanke, 2018: Potential impacts of climate change on vegetable production and product quality - A review. J Clean Prod, 170, 1602–1620, doi:10.1016/j.jclepro.2017.09.224.
Bishop, K. A., A.D.B. Leakey and E.A. Ainsworth, 2014: How seasonal temperature or water inputs affect the relative response of C-3 crops to elevated [CO2]: a global analysis of open top chamber and free air CO2 enrichment studies. Food Energy Secur, 3 (1), 33–45, doi:10.1002/fes3.44.
Bisht, I.S., et al., 2018: Farmers’ rights, local food systems, and sustainable household dietary diversification: A case of Uttarakhand Himalaya in north-western India. Agroecol. Sustain. Food Syst. , 42 (1), 77–113, doi:10.1080/21683565.2017.1363118.
Bitter, M.C., L. Kapsenberg, J.P. Gattuso and C.A. Pfister, 2019: Standing genetic variation fuels rapid adaptation to ocean acidification. Nat Commun, 10 (1), 5821, doi:10.1038/s41467-019-13767-1.
Blanchard, J.L., et al., 2017: Linked sustainability challenges and trade-offs among fisheries, aquaculture and agriculture. Nat Ecol Evol, 1 (9), 1240–1249, doi:10.1038/s41559-017-0258-8.
Blanco, J. and S.M. Carriere, 2016: Sharing local ecological knowledge as a human adaptation strategy to arid environments: Evidence from an ethnobotany survey in Morocco. J. Arid Environ. , 127, 30–43, doi:10.1016/j.jaridenv.2015.10.021.
Blaser, W.J., et al., 2018: Climate-smart sustainable agriculture in low-to-intermediate shade agroforests. Nat. Sustain. , 1 (5), 234–239, doi:10.1038/s41893-018-0062-8.
Blasiak, R., et al., 2017: Climate change and marine fisheries: Least developed countries top global index of vulnerability. PLoS ONE, 12 (6), e179632, doi:10.1371/journal.pone.0179632.
Blay-Palmer, A., R. Carey, E. Valette and M.R. Sanderson, 2020: Post COVID 19 and food pathways to sustainable transformation. Agric Human Values, 37 (3), 1–3, doi:10.1007/s10460-020-10051-7.
Blay-Palmer, A., et al., 2018: Validating the city region food system approach: enacting inclusive, transformational city region food systems. Sustainability, 10 (5), 1680, doi:10.3390/su10051680.
Bleischwitz, R., et al., 2018: Resource nexus perspectives towards the United Nations Sustainable Development Goals. Nat. Sustain. , 1 (12), 737–743, doi:10.1038/s41893-018-0173-2.
Blesh, J., et al., 2019: Development pathways toward “zero hunger”. World Dev, 118, 1–14, doi:10.1016/j.worlddev.2019.02.004.
Bloemen, P., et al., 2018: Lessons learned from applying adaptation pathways in flood risk management and challenges for the further development of this approach. Mitig Adapt Strateg Glob Change, 23 (7), 1083–1108, doi:10.1007/s11027-017-9773-9.
Blumenthal, D.M., et al., 2016: Cheatgrass is favored by warming but not CO2 enrichment in a semi-arid grassland. Glob. Chang. Biol. , 22 (9), 3026–3038, doi:10.1111/gcb.13278.
Blumenthal, D.M., et al., 2013: Invasive forb benefits from water savings by native plants and carbon fertilization under elevated CO2 and warming. New Phytol. , 200 (4), 1156–1165, doi:10.1111/nph.12459.
Blundell, R., et al., 2020: Organic management promotes natural pest control through altered plant resistance to insects. Nat. Plants, 6 (5), 483–491, doi:10.1038/s41477-020-0656-9.
Boafo, Y.A., et al., 2016: Provisioning ecosystem services-sharing as a coping and adaptation strategy among rural communities in Ghana’s semi-arid ecosystem. Ecosyst. Serv. , 19, 92–102, doi:10.1016/j.ecoser.2016.05.002.
Boddicker, R.L., et al., 2014: Gestational Heat Stress Alters Postnatal Offspring Body Composition Indices and Metabolic Parameters in Pigs. Plos One, 9 (11), e110859–e110859, doi:10.1371/journal.pone.0110859.
Bodin, P., S. Olin, T.A.M. Pugh and A. Arneth, 2016: Accounting for interannual variability in agricultural intensification: The potential of crop selection in Sub-Saharan Africa. Agric Syst , 148, 159–168, doi:10.1016/j.agsy.2016.07.012.
Bodmer, R., et al., 2018: Major shifts in Amazon wildlife populations from recent intensification of floods and drought. Conserv. Biol. , 32 (2), 333–344, doi:10.1111/cobi.12993.
Boedecker, J., et al., 2019: Participatory farm diversification and nutrition education increase dietary diversity in Western Kenya. Matern Child Nutr, 15 (3), e12803, doi:10.1111/mcn.12803.
Boedecker, J., et al., 2014: Dietary contribution of wild edible plants to women’s diets in the buffer zone around the Lama forest, Benin - an underutilized potential. Food Secur, 6 (6), 833–849, doi:10.1007/s12571-014-0396-7.
Bogdanova, E., et al., 2021: The impact of climate change on the food (in)security of the Siberian indigenous peoples in the Arctic: environmental and health risks. Sustainability, 13 (5), doi:10.3390/su13052561.
Bois, B., S. Zito and A. Calonnec, 2017: Climate vs grapevine pests and diseases worldwide : the first results of a global survey. Oeno One, 51 (2), 133–139, doi:10.20870/oeno-one.2016.0.0.1780.
Boissière, M., et al., 2013: Local perceptions of climate variability and change in tropical forests of Papua, Indonesia. Ecol. Soc. , 18 (4), 13, doi:10.5751/ES-05822-18041.
Bonada, M., et al., 2015: Impact of elevated temperature and water deficit on the chemical and sensory profiles of B arossa S hiraz grapes and wines. Aust J Grape Wine Res, 21 (2), 240–253, doi:10.1111/ajgw.12142.
Bonaudo, T., et al., 2014: Agroecological principles for the redesign of integrated crop-livestock systems. Eur J Agron, 57, 43–51, doi:10.1016/j.eja.2013.09.010.
Bond, W.J., N. Stevens, G.F. Midgley and C.E.R. Lehmann, 2019: The trouble with trees: afforestation plans for Africa. Trends Ecol. Evol. , 34 (11), 963–965, doi:10.1016/j.tree.2019.08.003.
Bondad-Reantaso, M.G., E. Garrido-Gamarro and S.E. McGladdery, 2018: Climate change-driven hazards on food safety and aquatic animal health. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 517–533. ISBN 978-9251306079.
Bondé, L., et al., 2019: Variability and estimating in fruiting of shea tree (Vitellaria paradoxa CF Gaertn) associated to climatic conditions in West Africa: implications for sustainable management and development. Plant. Prod. Sci. , 22 (2), 143–158, doi:10.1080/1343943X.2018.1541712.
Bonebrake, T.C., et al., 2018: Managing consequences of climate-driven species redistribution requires integration of ecology, conservation and social science. Biol. Rev. Camb. Philos. Soc. , 93 (1), 284–305, doi:10.1111/brv.12344.
Bonfante, A., et al., 2017: Evaluation of the effects of future climate change on grape quality through a physically based model application: a case study for the Aglianico grapevine in Campania region, Italy. Agric Syst , 152, 100–109, doi:10.1016/j.agsy.2016.12.009.
Bonny, S., 2017: Corporate concentration and technological change in the global seed industry. Sustainability, 9 (9), 1632, doi:10.3390/su9091632.
Boone, R.B., et al., 2018: Climate change impacts on selected global rangeland ecosystem services. Glob Chang Biol, 24 (3), 1382–1393, doi:10.1111/gcb.13995.
Booth, S. and D. Zeller, 2005: Mercury, food webs, and marine mammals: implications of diet and climate change for human health. Environ. Health Perspect. , 113 (5), 521–526, doi:10.1289/ehp.7603.
Borelli, S., M. Conigliaro, S. Quaglia and F. Salbitano, 2017: Urban and peri-urban agroforestry as multifunctional land use. In: Agroforestry: Anecdotal to Modern Science[Dagar, J.C. and V.P. Tewari(eds.)]. Springer Nature Singapore, Singapore, pp. 705–724. ISBN 978-9811076497.
Bornemann, F.J., et al., 2019: Future changes and uncertainty in decision-relevant measures of East African climate. Clim Change, 156 (3), 365–384, doi:10.1007/s10584-019-02499-2.
Borquez, R., P. Aldunce and C. Adler, 2017: Resilience to climate change: from theory to practice through co-production of knowledge in Chile. Sustain Sci, 12 (1), 163–176, doi:10.1007/s11625-016-0400-6.
Borras, S.M. and J.C. Franco, 2018a: The challenge of locating land-based climate change mitigation and adaptation politics within a social justice perspective : towards an idea of agrarian climate justice. Third World Q, 39 (7), 1308–1325, doi:10.1080/01436597.2018.1460592.
Borras, J. and J.C. Franco, 2018b: Agrarian climate justice: Imperative and opportunity. Transnational Institute, Amsterdam.
Borras, S.M., J.C. Franco and Z. Nam, 2020: Climate change and land: Insights from Myanmar. World Dev, 129, 104864–104864, doi:10.1016/j.worlddev.2019.104864.
Borras, S.M., et al., 2011: Towards a better understanding of global land grabbing: an editorial introduction. J Peasant Stud, 38 (2), 209–216, doi:10.1080/03066150.2011.559005.
Borsatto, R.S., M. A. Altieri, H.C. Duval and J. Perez-Cassarino, 2020: Public procurement as strategy to foster organic transition: insights from the Brazilian experience. Renew. Agric. Food Syst. , 35 (6), 688–696, doi:10.1017/S174217051900036X.
Bosch, G., et al., 2019: Conversion of organic resources by black soldier fly larvae: legislation, efficiency and environmental impact. J Clean Prod, 222, 355–363, doi:10.1016/j.jclepro.2019.02.270.
Bosma, R.H., et al., 2017: The financial feasibility of producing fish and vegetables through aquaponics. Aquac Eng, 78, 146–154, doi:10.1016/j.aquaeng.2017.07.002.
Bosomworth, K. and E. Gaillard, 2019: Engaging with uncertainty and ambiguity through participatory ‘Adaptive Pathways’ approaches: scoping the literature. Environ. Res. Lett. , 14 (9), 93007, doi:10.1088/1748-9326/ab3095.
Bosso, L., et al., 2016: Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa in the Mediterranean basin. Biol Invasions, 18 (6), 1759–1768, doi:10.1007/s10530-016-1118-1.
Botana, L.M., 2016: Toxicological perspective on climate change: aquatic toxins. Chem. Res. Toxicol. , 29 (4), 619–625, doi:10.1021/acs.chemrestox.6b00020.
Botana, L.M. and M.J. Sainz (eds.), 2015: Climate Change and Mycotoxins. De Gruyter, Berlin, Germany, München, Germany, and Boston, MA, USA, 185. ISBN 978-3110333053.
Bottazzi, P., D. Crespo, L.O. Bangura and S. Rist, 2018: Evaluating the livelihood impacts of a large-scale agricultural investment: Lessons from the case of a biofuel production company in northern Sierra Leone. Land Use Policy, 73, 128–137, doi:10.1016/j.landusepol.2017.12.016.
Bottero, A., et al., 2017: Density-dependent vulnerability of forest ecosystems to drought. J Appl Ecol, 54 (6), 1605–1614, doi:10.1111/1365-2664.12847.
Bouahmed, A., et al., 2019: Modeling Cedrus atlantica potential distribution in North Africa across time: new putative glacial refugia and future range shifts under climate change. Reg Environ Change, 19 (6), 1667–1682, doi:10.1007/s10113-019-01503-w.
Boulanger-Lapointe, N., et al., 2019: Berry plants and berry picking in Inuit Nunangat: traditions in a changing socio-ecological landscape. Hum Ecol, 47 (1), 81–93, doi:10.1007/s10745-018-0044-5.
Boullis, A., C. Detrain, F. Francis and F.J. Verheggen, 2016: Will climate change affect insect pheromonal communication?Curr. Opin. Insect. Sci. , 17, 87–91, doi:10.1016/j.cois.2016.08.006.
Bourgault, M., et al., 2017: Yield, growth and grain nitrogen response to elevated CO2 in six lentil (Lens culinaris) cultivars grown under Free Air CO2 Enrichment (FACE) in a semi-arid environment. Eur J Agron, 87, 50–58, doi:10.1016/j.eja.2017.05.003.
Bourgault, M., et al., 2018: Effect of a heat wave on lentil grown under free-air CO2 enrichment (FACE) in a semi-arid environment. Crop Sci. , 58, 803–812, doi:0.2135/cropsci2017.09.0565.
Bourke, M., A. Atkinson and T. Neale, 2020: Putting Country back together: a conversation about collaboration and Aboriginal fire management. Postcolonial Stud. , 23 (4), 546–551, doi:10.1080/13688790.2020.1751909.
Bouroncle, C., et al., 2017: Mapping climate change adaptive capacity and vulnerability of smallholder agricultural livelihoods in Central America: ranking and descriptive approaches to support adaptation strategies. Clim Change, 141 (1), 123–137, doi:10.1007/s10584-016-1792-0.
Bouroncle, C., et al., 2019: A systematic approach to assess climate information products applied to agriculture and food security in Guatemala and Colombia. Clim. Serv. , 16, 100137, doi:A10.1016/j.cliser.2019.100137.
Bowles, T.M., et al., 2020: Long-term evidence shows that crop-rotation diversification increases agricultural resilience to adverse growing conditions in North America. One Earth, 2 (3), 284–293, doi:10.1016/j.oneear.2020.02.007.
Boyd, E., M. Gutierrez and M.Y. Chang, 2007: Small-scale forest carbon projects: adapting CDM to low-income communities. Glob. Environ. Change. , 17 (2), 250–259, doi:10.1016/j.gloenvcha.2006.10.001.
Bozzola, M. and M. Smale, 2020: The welfare effects of crop biodiversity as an adaptation to climate shocks in Kenya. World Dev, 135, 105065, doi:10.1016/j.worlddev.2020.105065.
Bradai, L., S. Bissati, H. Chenchouni and K. Amrani, 2015: Effects of climate on the productivity of desert truffles beneath hyper-arid conditions. Int. J. Biometeorol. , 59 (7), 907–915, doi:10.1007/s00484-014-0891-8.
Bradley, B.A., D.M. Blumenthal, D.S. Wilcove and L.H. Ziska, 2010: Predicting plant invasions in an era of global change. Trends Ecol. Evol. , 25 (5), 310–318, doi:10.1016/j.tree.2009.12.003.
Bradley, B.A., C.A. Curtis and J.C. Chambers, 2016: Bromus response to climate and projected changes with climate change. In: Exotic Brome-Grasses in Arid and Semiarid Ecosystems of the Western US: Causes, Consequences, and Management Implications[Germino, M.J., J.C. Chambers and C.S. Brown(eds.)]. Springer, Cham, Switzerland, pp. 257–274. ISBN 978-3319249285.
Bradshaw, C.J., et al., 2016: Massive yet grossly underestimated global costs of invasive insects. Nat Commun, 7, 12986, doi:10.1038/ncomms12986.
Braga, H.O., et al., 2020: The role of local ecological knowledge for the conservation and sustainable fisheries of the sea lamprey (Petromyzon marinus Linnaeus, 1758) in the Iberian Peninsula. Ocean. Coast. Manag. , 198, 105345, doi:10.1016/j.ocecoaman.2020.105345.
Branca, F., et al., 2019: Transforming the food system to fight non-communicable diseases. BMJ, 364, l296, doi:10.1136/bmj.l296.
Brancalion, P.H.S. and R.L. Chazdon, 2017: Beyond hectares: four principles to guide reforestation in the context of tropical forest and landscape restoration. Restor. Ecol. , 25 (4), 491–496, doi:10.1111/rec.12519.
Brander, K., K. Cochrane, M. Barange and D. Soto, 2018: Climate change implications for fisheries and aquaculture. In: Climate Change Impacts on Fisheries and Aquaculture[Phillips, B.F. and M. Pérez-Ramírez(eds.)]. Wiley Blackwell, Hoboken, NJ, USA and Chichester, UK, pp. 45–62. ISBN 978-1119154044.
Brandt, J.S., et al., 2013: Regime shift on the roof of the world: alpine meadows converting to shrublands in the southern Himalayas. Biol. Conserv. , 158, 116–127, doi:10.1016/j.biocon.2012.07.026.
Brás, T.A., J. Seixas, N. Carvalhais and J. Jägermeyr, 2021: Severity of drought and heatwave crop losses tripled over the last five decades in Europe. Environ. Res. Lett. , 16, 65012, doi:10.1088/1748-9326/abf004.
Brassard, J.P. and B. Singh, 2008: Impacts of climate change and CO2 increase on agricultural production and adaptation options for Southern Quebec, Canada. Mitig Adapt Strateg Glob Change, 13 (3), 241–265, doi:10.1007/s11027-007-9109-2.
Brecka, A.F.J., et al., 2020: Sustainability of Canada’s forestry sector may be compromised by impending climate change. For. Ecol. Manag. , 474, 118352, doi:10.1016/j.foreco.2020.118352.
Brendler, T., J.A. Brinckmann and U. Schippmann, 2018: Sustainable supply, a foundation for natural product development: The case of Indian frankincense (Boswellia serrata Roxb. ex Colebr.). J. Ethnopharmacol. , 225, 279–286, doi:10.1016/j.jep.2018.07.017.
Bresnahan, P.J., et al., 2020: Equipping smart coasts with marine water quality IoT sensors. Results Eng, 5, doi:10.1016/j.rineng.2019.100087.
Breu, T., et al., 2016: Large-scale land acquisition and its effects on the water balance in investor and host countries. PLoS ONE, 11 (3), 1–18, doi:10.1371/journal.pone.0150901.
Bricknell, I.R., et al., 2021: Resilience of cold water aquaculture: a review of likely scenarios as climate changes in the Gulf of Maine. Rev. Aquac. , 13 (1), 460–503, doi:10.1111/raq.12483.
Brinkman, T.J., et al., 2016: Arctic communities perceive climate impacts on access as a critical challenge to availability of subsistence resources. Clim Change, 139, 413–427, doi:10.1007/s10584-016-1819-6.
Briske, D.D., et al., 2021: Future climate variability will challenge rangeland beef cattle production in the Great Plains. Rangelands, 43 (1), 29–36, doi:10.1016/j.rala.2020.11.001.
Brito, B.P., et al., 2017: Review of the global distribution of foot-and-mouth disease virus from 2007 to 2014. Transbound Emerg Dis, 64 (2), 316–332, doi:10.1111/tbed.12373.
Britten, G.L., M. Dowd, L. Kanary and B. Worm, 2017: Extended fisheries recovery timelines in a changing environment. Nat Commun, 8, 15325, doi:10.1038/ncomms15325.
Brockington, J.D., I.M. Harris and R.M. Brook, 2016: Beyond the project cycle: a medium-term evaluation of agroforestry adoption and diffusion in a south Indian village. Agrofor. Syst. , 90 (3), 489–508, doi:10.1007/s10457-015-9872-0.
Brodie, J.F. and H.K. Gibbs, 2009: Bushmeat hunting as climate threat. Science, 326 (5951), 364–365, doi:10.1126/science.326_364b.
Brondizio, E.S., J. Settele, S. Diaz and H.T. Ngo (eds.), 2019: Global Assessment Report on Biodiversity and Ecosystem Services. Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, Bonn, Germany, ISBN 978-3947851201. 1148 pp.
Brooks, K., et al., 2019: Climate and Jobs for Rural Young People. IFAD. Rome, Italy, 40 pp.
Brottem, L. and B. Brooks, 2018: Crops and livestock under the sun: obstacles to rural livelihood adaptations to hotter 21st century temperatures in eastern Senegal. Land Degrad. Dev. , 29 (1), 118–126, doi:10.1002/ldr.2844.
Brown, C.J., et al., 2016: Ecological and methodological drivers of species’ distribution and phenology responses to climate change. Glob. Chang. Biol. , 22 (4), 1548–1560, doi:10.1111/gcb.13184.
Brozynska, M., A. Furtado and R.J. Henry, 2016: Genomics of crop wild relatives: expanding the gene pool for crop improvement. Plant Biotechnol J, 14, 1070–1085, doi:10.1111/pbi.12454.
Brugère, C., J. Aguilar-Manjarrez, M.C.M. Beveridge and D. Soto, 2019: The ecosystem approach to aquaculture 10 years on - a critical review and consideration of its future role in blue growth. Rev. Aquac. , 11 (3), 493–514, doi:10.1111/raq.12242.
Brugère, C. and C. De Young, 2015: Assessing Climate Change Vulnerability in Fisheries and Aquaculture: Available Methodologies and Their Relevance for the Sector. FAO Fisheries and Aquaculture Technical Paper. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251089460. 86 pp.
Brugere, C., et al., 2020: Can innovation empower? Reflections on introducing tubular nets to women seaweed farmers in Zanzibar. Gend. Technol. Dev. , 24 (1), 89–109, doi:10.1080/09718524.2019.1695307.
Brugnach, M., M. Craps and A. Dewulf, 2017: Including indigenous peoples in climate change mitigation: addressing issues of scale, knowledge and power. Clim Change, 140 (1), 19–32, doi:10.1007/s10584-014-1280-3.
Brun, P., et al., 2020: Large-scale early-wilting response of Central European forests to the 2018 extreme drought. Glob Chang Biol, 26 (12), 7021–7035, doi:10.1111/gcb.15360.
Bruna, N., 2019: Land of plenty, land of misery: Synergetic resource grabbing in. Mozambique. land, 8 (8), doi:10.3390/land8080113.
Brunette, M., R. Bourke, M. Hanewinkel and R. Yousefpour, 2018: Adaptation to climate change in forestry: a multiple correspondence analysis (MCA). Forests, 9 (1), 20, doi:10.3390/f9010020.
Bruyere, B.L., et al., 2018: Paired comparison of visions for the future among young pastoralists and students in Samburu, Kenya. Pastoralism, 8, 21, doi:10.1186/s13570-018-0127-3.
Bryndum-Buchholz, A., et al., 2019: Twenty-first-century climate change impacts on marine animal biomass and ecosystem structure across ocean basins. Glob Chang Biol, 25 (2), 459–472, doi:10.1111/gcb.14512.
Buchner, B., et al., 2019: Global Landscape of Climate Finance 2019. Climate Policy Initiative, London, UK, https://www.climatepolicyinitiative.org/publication/global-landscape-of-climate-finance-2019/. Accessed 2021 . (36 pp).
Buckley, Y.M. and A.M. Csergo, 2017: Predicting invasion winners and losers under climate change. Proc. Natl. Acad. Sci. U. S. A. , 114 (16), 4040–4041, doi:10.1073/pnas.1703510114.
Budiman, I., 2019: Climate-smart agriculture policy and (in)justice for smallholders in developing countries. Future Food: J. Food Agric. Soc. , 7 (1), 31–41, doi:10.17170/kobra-2018122074.
Bueno, P.B. and D. Soto, 2017: Adaptation Strategies of the Aquaculture Sector to the Impacts of Climate Change. FAO Fisheries and Aquaculture Circular, Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251096864. 28 pp.
Bugge, M.M., S. Bolwig, T. Hansen and A.N. Tanner, 2019: Theoretical perspectives on innovation for waste valorisation in the bioeconomy. In: From Waste to Value[Klitkou, A., A.M. Fevolden and M. Capasso(eds.)]. Routledge, London, pp. 51–70. ISBN 978-0429460289.
Bukari, K.N. and E.D. Kuusaana, 2018: Impacts of large-scale land holdings on Fulani pastoralists’ in the Agogo Traditional Area of Ghana. Land Use Policy, 79 (November 2016), 748–758, doi:10.1016/j.landusepol.2018.09.018.
Bulengela, G., et al., 2020: “Bring fishermen at the center”: the value of local knowledge for understanding fisheries resources and climate-related changes in Lake Tanganyika. Environ. Dev. Sustain. , 22 (6), 5621–5649, doi:10.1007/s10668-019-00443-z.
Bunting, S.W., N. Kundu and N. Ahmed, 2017: Evaluating the contribution of diversified shrimp-rice agroecosystems in Bangladesh and West Bengal, India to social-ecological resilience. Ocean Coast. Manag. , 148, 63–74, doi:10.1016/j.ocecoaman.2017.07.010.
Buongiorno, J., 2015: Modeling some long-term implications of CO2 fertilization for global forests and forest industries. For. Ecosyst. , 2, 29, doi:10.1186/s40663-015-0054-3.
Buontempo, C., et al., 2019: Fostering the development of climate services through Copernicus Climate Change Service (C3S) for agriculture applications. Weather. Clim. Extrem. , 27, 100226, doi:10.1016/j.wace.2019.100226.
Buras, A., A. Rammig and C.S. Zang, 2020: Quantifying impacts of the 2018 drought on European ecosystems in comparison to 2003. Biogeosciences, 17 (6), 1655–1672, doi:10.5194/bg-17-1655-2020.
Burchfield, E.K. and A.T. de la Poterie, 2018: Determinants of crop diversification in rice-dominated Sri Lankan agricultural systems. J Rural Stud, 61, 206–215, doi:10.1016/j.jrurstud.2018.05.010.
Burden, M. and R. Fujita, 2019: Better fisheries management can help reduce conflict, improve food security, and increase economic productivity in the face of climate change. Mar. Policy. , 108, 103610, doi:10.1016/j.marpol.2019.103610.
Burney, J. and V. Ramanathan, 2014: Recent climate and air pollution impacts on Indian agriculture. Proc. Natl. Acad. Sci. U. S. A. , 111 (46), 16319–16324, doi:10.1073/pnas.1317275111.
Burton, R.J.F., 2019: The potential impact of synthetic animal protein on livestock production: the new “war against agriculture”?J Rural Stud, 68, 33–45, doi:10.1016/j.jrurstud.2019.03.002.
Bush, R. and G. Martiniello, 2017: Food riots and protest: agrarian modernizations and structural crises. World Dev, 91, 193–207, doi:10.1016/j.worlddev.2016.10.017.
Butler, E.E., N.D. Mueller and P. Huybers, 2018: Peculiarly pleasant weather for US maize. Proc. Natl. Acad. Sci. U. S. A. , 115 (47), 11935–11940, doi:10.1073/pnas.1808035115.
Butler, J.R.A., et al., 2020: Adapting scenarios for climate adaptation: practitioners’ perspectives on a popular planning method. Environ Sci Policy, 104, 13–19, doi:10.1016/j.envsci.2019.10.014.
Butler, J.R.A., S. Busilacchi and T. Skewes, 2019: How resilient is the Torres Strait Treaty (Australia and Papua New Guinea) to global change? A fisheries governance perspective. Environ Sci Policy, 91, 17–26, doi:10.1016/j.envsci.2018.10.005.
Butler, J.R.A., et al., 2016: Priming adaptation pathways through adaptive co-management: design and evaluation for developing countries. Clim. Risk Manag. , 12, 1–16, doi:10.1016/j.crm.2016.01.001.
Buytaert, W., J. Friesen, J. Liebe and R. Ludwig, 2012: Assessment and management of water resources in developing, semi-arid and arid regions. Water Resour. Manag. , 26 (4), 841–844, doi:10.1007/s11269-012-9994-3.
Byerly, H., et al., 2018: Nudging pro-environmental behavior: evidence and opportunities. Front Ecol Environ, 16 (3), 159–168, doi:10.1002/fee.1777.
Cable, J., et al., 2017: Global change, parasite transmission and disease control: lessons from ecology. Philos. Trans. Royal Soc. B: Biol. Sci. , 372 (1719), 20160088, doi:10.1098/rstb.2016.0088.
Cacho, J.F., M.C. Negri, C.R. Zumpf and P. Campbell, 2018: Introducing perennial biomass crops into agricultural landscapes to address water quality challenges and provide other environmental services. Wiley Interdiscip. Rev. Energy Environ. , 7 (2), e275–e275, doi:10.1002/wene.275.
Cadiz, L., et al., 2018: Moderate hypoxia but not warming conditions at larval stage induces adverse carry-over effects on hypoxia tolerance of European sea bass (Dicentrarchus labrax) juveniles. Mar. Environ. Res. , 138, 28–35.
Cagalanan, D., 2016: Public-private partnerships for improved reforestation outcomes in the Philippines. World Dev. Perspect. , 3, 32–34, doi:10.1016/j.wdp.2016.11.004.
Cai, J.N., H. Huang and P.S. Leung, 2019: Understanding and measuring the contribution of aquaculture and fisheries to gross domestic product (GDP). FAO Fisheries and Aquaculture Technical Paper, Vol. 606. FAO, Rome, Italy, ISBN 978-9251312803.
Cai, W., et al., 2018: Stabilised frequency of extreme positive Indian Ocean Dipole under 1.5 degrees C warming. Nat Commun, 9 (1), 1419, doi:10.1038/s41467-018-03789-6.
Cai, W.J., et al., 2014: Increasing frequency of extreme El Nino events due to greenhouse warming. Nat. Clim. Chang. , 4 (2), 111–116, doi:10.1038/Nclimate2100.
Cai, Y.Y., J.S. Bandara and D. Newth, 2016: A framework for integrated assessment of food production economics in South Asia under climate change. Environ. Model. Softw. , 75, 459–497, doi:10.1016/j.envsoft.2015.10.024.
Calderón, C.I., et al., 2018: Agroecology-based farming provides grounds for more resilient livelihoods among smallholders in Western Guatemala. Agroecol. Sustain. Food Syst. , 42, 1128–1169, doi:10.1080/21683565.2018.1489933.
Call, M. and S. Sellers, 2019: How does gendered vulnerability shape the adoption and impact of sustainable livelihood interventions in an era of global climate change?Environ. Res. Lett. , 14 (8), doi:10.1088/1748-9326/ab2f57.
Callen, S.T., L.L. Klein and A.J. Miller, 2016: Climatic niche characterization of 13 North American Vitis species. Am J Enol Vitic. , 67 (3), 339–349, doi:10.5344/ajev.2016.15110.
Calvet-Mir, L., E. Gomez-Baggethun and V. Reyes-Garcia, 2012: Beyond food production: ecosystem services provided by home gardens. A case study in Vall Fosca, Catalan Pyrenees, Northeastern Spain. Ecol. Econ. , 74, 153–160, doi:10.1016/j.ecolecon.2011.12.011.
Camacho-Henriquez, A., et al., 2015: Decentralized collaborative plant breeding for utilization and conservation of neglected and underutilized crop genetic resources. In: Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools[Al-Khayri, J.M., S.M. Jain and D.V. Johnson(eds.)]. Springer, Cham, Switzerland, pp. 25–61. ISBN 978-3319225203.
Camacho, A. and E. Conover, 2019: The impact of receiving SMS price and weather information on small scale farmers in Colombia. World Dev, 123, 104596, doi:10.1016/j.worlddev.2019.06.020.
Camacho, L.D., D.T. Gevaña, A.P. Carandang and S.C. Camacho, 2016: Indigenous knowledge and practices for the sustainable management of Ifugao forests in Cordillera, Philippines. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. , 12, 5–13, doi:10.1080/21513732.2015.1124453.
Campana, S.E., et al., 2020: Arctic freshwater fish productivity and colonization increase with climate warming. Nat. Clim. Chang. , 10 (5), 428–433, doi:10.1038/s41558-020-0744-x.
Campanaro, A., et al., 2019: DNA barcoding to promote social awareness and identity of neglected, underutilized plant species having valuable nutritional properties. Food. Res. Int. , 115, 1–9, doi:10.1016/j.foodres.2018.07.031.
Campbell-Arvai, V., J. Arvai and L. Kalof, 2012: Motivating Sustainable Food Choices: The Role of Nudges, Value Orientation, and Information Provision. Environ Behav, 46 (4), 453–475, doi:10.1177/0013916512469099.
Campbell, B.M., et al., 2017: Agriculture production as a major driver of the Earth system exceeding planetary boundaries. Ecol. Soc. , 22 (4), 8, doi:10.5751/Es-09595-220408.
Campbell, B.M., et al., 2018: Urgent action to combat climate change and its impacts (SDG 13): transforming agriculture and food systems. Curr. Opin. Environ. Sustain. , 34, 13–20, doi:10.1016/j.cosust.2018.06.005.
Campbell, B.M., et al., 2016: Reducing risks to food security from climate change. Glob. Food Secur. Policy Econ. Environ. , 11, 34–43, doi:10.1016/j.gfs.2016.06.002.
Canadell, J.G., P.M.S. Monteiro, M.H. Costa, L. Cotrim da Cunha, P.M. Cox, A.V. Eliseev, S. Henson, M. Ishii, S. Jaccard, C. Koven, A. Lohila, P.K. Patra, S. Piao, J. Rogelj, S. Syampungani, S. Zaehle and K. Zickfeld, 2021: Global Carbon and other Biogeochemical Cycles and Feedbacks. In: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. [Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press., Cambridge, UK.
Canevari-Luzardo, L.M., F. Berkhout and M. Pelling, 2019: A relational view of climate adaptation in the private sector: how do value chain interactions shape business perceptions of climate risk and adaptive behaviours?Bus. Strategy Environ. , 29 (2), 432–444, doi:10.1002/bse.2375.
Cao, J.J., et al., 2018a: Can individual land ownership reduce grassland degradation and favor socioeconomic sustainability on the Qinghai-Tibetan Plateau?Environ Sci Policy, 89, 192–197, doi:10.1016/j.envsci.2018.08.003.
Cao, R., et al., 2018b: CO2-induced ocean acidification impairs the immune function of the Pacific oyster against Vibrio splendidus challenge: an integrated study from a cellular and proteomic perspective. Sci. Total Environ. , 625, 1574–1583, doi:10.1016/j.scitotenv.2018.01.056.
Cardell, M.F., A. Amengual and R. Romero, 2019: Future effects of climate change on the suitability of wine grape production across Europe. Reg. Environ. Change, 19 (8), 2299–2310, doi:10.1007/s10113-019-01502-x.
Carey, J.C., et al., 2016: Temperature response of soil respiration largely unaltered with experimental warming. Proc. Natl. Acad. Sci. U. S. A. , 113 (48), 13797–13802, doi:10.1073/pnas.1605365113.
Caron, P., et al., 2018: Food systems for sustainable development: proposals for a profound four-part transformation. Agron. Sustain. Dev. , 38 (4), 41, doi:10.1007/s13593-018-0519-1.
Carr, E.R. and M.C. Thompson, 2014: Gender and climate change adaptation in agrarian settings: current thinking, new directions, and research frontiers. Geogr. Compass, 8 (3), 182–197, doi:10.1111/gec3.12121.
Carrasco, D., et al., 2018: With or without you: Effects of the concurrent range expansion of an herbivore and its natural enemy on native species interactions. Glob Chang Biol, 24 (2), 631–643, doi:10.1111/gcb.13836.
Carro-Ripalda, S. and M. Astier, 2014: Silenced voices, vital arguments: smallholder farmers in the Mexican GM maize controversy. Agric Hum Values, 31 (4), 655–663, doi:10.1007/s10460-014-9533-3.
Carsan, S., et al., 2014: Can agroforestry option values improve the functioning of drivers of agricultural intensification in Africa?Curr. Opin. Environ. Sustain. , 6, 35–40, doi:10.1016/j.cosust.2013.10.007.
Carter, B., et al., 2014: Coral reef, water quality status and community understanding of threats in the eastern gulf of Thailand. APN Sci. Bull. , 4, 76–78.
Carter, S., et al., 2017: Large scale land acquisitions and REDD plus : a synthesis of conflicts and opportunities. Environ. Res. Lett. , 12 (3), doi:10.1088/1748-9326/aa6056.
Carvalhais, N., et al., 2014: Global covariation of carbon turnover times with climate in terrestrial ecosystems. Nature, 514 (7521), 213–217, doi:10.1038/nature13731.
Castagneri, D., et al., 2021: Meta-analysis reveals different competition effects on tree growth resistance and resilience to drought. Ecosystems, doi:10.1007/s10021-021-00638-4. In press.
Castañeda-Álvarez, N.P., et al., 2016: Global conservation priorities for crop wild relatives. Nat. Plants, 2 (4), 16022, doi:10.1038/nplants.2016.22.
Castellanos-Frías, E., D.G. De León, F. Bastida and J.L. González-Andújar, 2016: Predicting global geographical distribution of Lolium rigidum (rigid ryegrass) under climate change. J. Agric. Sci. , 154 (5), 755–764, doi:10.1017/S0021859615000799.
Catacora-Vargas, G., et al., 2017: Brief historical review of agroecology in Bolivia. Agroecol. Sustain. Food Syst. , 41 (3-4), 429–447, doi:10.1080/21683565.2017.1290732.
Catalán, I. A., et al., 2019: Critically examining the knowledge base required to mechanistically project climate impacts: a case study of Europe’s fish and shellfish. Fish. Fish. , 20 (3), 501–517, doi:10.1111/faf.12359.
Cavaliere, C., 2009: The effects of climate change on medicinal and aromatic plants. Herb. Gram, 81, 44–57.
Ceccarelli, S. and S. Grando, 2020: Participatory plant breeding: who did it, who does it and where?Exp. Agric. , 56 (1), 1–11, doi:10.1017/s0014479719000127.
Celentano, D., et al., 2017: Land Use Policy Towards zero deforestation and forest restoration in the Amazon region of Maranhão state, Brazil. Land Use Policy, 68 (July), 692–698, doi:10.1016/j.landusepol.2017.07.041.
Cerato, S., A.R. Davis, D. Coleman and M.Y.L. Wong, 2019: Reversal of competitive dominance between invasive and native freshwater crayfish species under near-future elevated water temperature. Aquat. Invasions, 14 (3), 518–530, doi:10.3391/ai.2019.14.3.10.
Cerda, R., et al., 2017: Effects of shade, altitude and management on multiple ecosystem services in coffee agroecosystems. Eur J Agron, 82, 308–319, doi:10.1016/j.eja.2016.09.019.
Chakravorty, U., M.H. Hubert, M. Moreaux and L. Nøstbakken, 2017: Long-Run Impact of Biofuels on Food Prices. Scand J Econ, 119 (3), 733–767, doi:10.1111/sjoe.12177.
Challinor, A., et al., 2014: Making the most of climate impacts ensembles. Nat. Clim. Chang. , 4 (2), 77–80, doi:10.1038/nclimate2117.
Challinor, A.J., et al., 2018: Transmission of climate risks across sectors and borders. Philos. Trans. A. Math. Phys. Eng. Sci. , 376, 20170301, doi:10.1098/rsta.2017.0301.
Challinor, A.J., et al., 2016: Current warming will reduce yields unless maize breeding and seed systems adapt immediately. Nat. Clim. Chang. , 6 (10), 954–958, doi:10.1038/Nclimate3061.
Chambault, P., et al., 2018: Sea surface temperature predicts the movements of an Arctic cetacean: the bowhead whale. Sci Rep, 8 (1), 9658, doi:10.1038/s41598-018-27966-1.
Chamberlin, J. and J. Sumberg, 2021: Are young people transforming the rural economy? In Youth and the Rural Economy in Africa: Hard Work and Hazard. Ed. J. Sumberg. 92–124, CABI, Wallingford, UK, doi:10.1079/9781789245011.0006.
Chan, K.M. A., T. Satterfield and J. Goldstein, 2012: Rethinking ecosystem services to better address and navigate cultural values. Ecol. Econ. , 74, 8–18, doi:10.1016/j.ecolecon.2011.11.011.
Chanamuto, N.J.C. and S.J.G. Hall, 2015: Gender equality, resilience to climate change, and the design of livestock projects for rural livelihoods. Gend. Dev. , 23 (3), 515–530, doi:10.1080/13552074.2015.1096041.
Chanana-Nag, N. and P.K. Aggarwal, 2020: Woman in agriculture, and climate risks: hotspots for development. Clim Change, 158 (1), 13–27, doi:10.1007/s10584-018-2233-z.
Chandra, A., K.E. McNamara and P. Dargusch, 2017: The relevance of political ecology perspectives for smallholder climate-smart agriculture: a review. J. Polit. Ecol. , 24, 821–842, doi:10.2458/v24i1.20969.
Chandra, A., K.E. McNamara and P. Dargusch, 2018: Climate-smart agriculture: perspectives and framings. Clim. Policy, 18 (4), 526–541, doi:10.1080/14693062.2017.1316968.
Chang-Fung-Martel, J., et al., 2017: The impact of extreme climatic events on pasture-based dairy systems: a review. Crop. Pasture Sci. , 68 (12), 1158–1158, doi:10.1071/cp16394.
Chang, J., et al., 2017: Future productivity and phenology changes in European grasslands for different warming levels: Implications for grassland management and carbon balance. Carbon Balance Manage, 12 (1), doi:10.1186/s13021-017-0079-8.
Chang, J.D., et al., 2016: Effects of elevated CO2 and temperature on Gynostemma pentaphyllum physiology and bioactive compounds. J. Plant Physiol. , 196-197, 41–52, doi:10.1016/j.jplph.2016.02.020.
Chapagain, D., F. Baarsch, M. Schaeffer and S. D’Haen, 2020: Climate change adaptation costs in developing countries: insights from existing estimates. Clim. Dev. , 12 (10), 934–942, doi:10.1080/17565529.2020.1711698.
Chapin, F.S., et al., 2016: Community-empowered adaptation for self-reliance. Curr. Opin. Environ. Sustain. , 19, 67–75, doi:10.1016/j.cosust.2015.12.008.
Chaplin-Kramer, R., et al., 2014: Global malnutrition overlaps with pollinator-dependent micronutrient production. Proc. Biol. Sci. , 281 (1794), 20141799, doi:10.1098/rspb.2014.1799.
Chapman, E.J., et al., 2020: Effects of climate change on coastal ecosystem food webs: Implications for aquaculture. Mar. Environ. Res. , 162, 105103, doi:10.1016/j.marenvres.2020.105103.
Chapman, S., et al., 2017: The impact of urbanization and climate change on urban temperatures: a systematic review. Landsc Ecol, 32 (10), 1921–1935, doi:10.1007/s10980-017-0561-4.
Chatuphale, P. and L. Armstrong, 2018: Indian mobile agricultural services using big data and internet of things (IoT). In: Intelligent Systems Design and Applications[Abraham, A., P.K. Muhuri, A.K. Muda and N. Gandhi(eds.)]. 17th International Conference on Intelligent Systems Design and Applications, 14-16 December 2017, New Delhi, India. Vol. 736, Springer, Cham, Switzerland, pp. 1028–1037. doi:10.1007/978-3-319-76348-4.
Chaubey, O.P., A. Sharma and G. Krishnamurthy, 2015: Ex-situ conservation of indigenous, threatened and ethno-medicinal diversity of forest species. Int. J. Biosci. Bio-technology, 7 (3), 9–22.
Chávez, C., J. Dresdner, Y. Figueroa and M. Quiroga, 2019: Main issues and challenges for sustainable development of salmon farming in Chile: a socio-economic perspective. Rev. Aquac. , 11 (2), 403–421, doi:10.1111/raq.12338.
Chemura, A., D. Kutywayo, P. Chidoko and C. Mahoya, 2016: Bioclimatic modelling of current and projected climatic suitability of coffee (Coffea arabica) production in Zimbabwe. Reg Environ Change, 16 (2), 473–485, doi:10.1007/s10113-015-0762-9.
Chen, B.W. and N.B. Villoria, 2019: Climate shocks, food price stability and international trade: evidence from 76 maize markets in 27 net-importing countries. Environ. Res. Lett. , 14 (1), 14007, doi:10.1088/1748-9326/aaf07 f.
Chen, C., et al., 2018a: Exposure to aflatoxin and fumonisin in children at risk for growth impairment in rural Tanzania. Environ Int , 115, 29–37, doi:10.1016/j.envint.2018.03.001.
Chen, C., Y.M. Pang, X.B. Pan and L.Z. Zhang, 2015a: Impacts of climate change on cotton yield in China from 1961 to 2010 based on provincial data. J. Meteorol. Res. , 29 (3), 515–524, doi:10.1007/s13351-014-4082-7.
Chen, J.-L., 2020: Fishers’ perceptions and adaptation on climate change in northeastern Taiwan. Environ. Dev. Sustain. , 23 (1), 611–634, doi:10.1007/s10668-020-00598-0.
Chen, J., et al., 2020: Soil carbon loss with warming: New evidence from carbon-degrading enzymes. Glob Chang Biol, doi:10.1111/gcb.14986.
Chen, J., K. Kosec and V. Mueller, 2019: Moving to despair? Migration and well-being in Pakistan. World Dev, 113, 186–203, doi:10.1016/j.worlddev.2018.09.007.
Chen, J. and V. Mueller, 2018: Coastal climate change, soil salinity and human migration in Bangladesh. Nat. Clim. Chang. , 8 (11), 981–985, doi:10.1038/s41558-018-0313-8.
Chen, T.-L. and H.-W. Cheng, 2020: Applying traditional knowledge to resilience in coastal rural villages. Int. J. Disaster Risk Reduct. , 47, 101564–101564, doi:10.1016/j.ijdrr.2020.101564.
Chen, Y.S., A.S. Todd, M.H. Murphy and G. Lomnicky, 2016: Anticipated water quality changes in response to climate change and potential consequences for inland fishes. Fisheries, 41 (7), 413–416, doi:10.1080/03632415.2016.1182509.
Chen, Y.Z., et al., 2018b: Great uncertainties in modeling grazing impact on carbon sequestration: a multi-model inter-comparison in temperate Eurasian Steppe. Environ. Res. Lett. , 13 (7), 75005, doi:10.1088/1748-9326/aacc75.
Chen, Z., H. Shang, J.X. Cao and H. Yu, 2015b: Effects of ambient ozone concentrations on contents of nonstructural carbohydrates in Phoebe bournei and Pinus massoniana seedlings in subtropical China. Water Air Soil Pollut. , 226 (9), 310, doi:10.1007/s11270-015-2555-7.
Cheung, W.W.L., B.J. and B.M., 2019: Projected changes in global and national potential marine fisheries catch under climate change scenarios in the twenty-first century. In: Impacts of climate change on fisheries and aquaculture, [Barange, M., T. Bahri, M. Beveridge, K. Cochrane, S. Funge-Smith and F. Poulain (eds.)]. FAO, Rome, Italy, pp. 63-85.
Chiarelli, D.D., et al., 2021: Large-scale land acquisition as a potential driver of slope instability. Land Degrad Dev, 32 (4), 1773–1785, doi:10.1002/ldr.3826.
Chiarelli, D.D., K.F. Davis, M.C. Rulli and P. D’Odorico, 2016: Climate change and large-scale land acquisitions in Africa: Quantifying the future impact on acquired water resources. Adv. Water. Resour. , 94, 231–237, doi:10.1016/j.advwatres.2016.05.016.
Chiayarak, B., G. Tattiyakul and N. Kamsunthad, 2019: Climate Change Vulnerability Assessment Bang Pakong River Wetland, Thailand. X, International Union of Conservation of Nature and Natural Resources Asia Regional Office, Bangkok, Thailand, 73 pp. Available at: https://www.iucn.org/sites/dev/files/climate_change_vulnerability_assessment_bang_pakong_river_wetland_thailand.pdf (accessed 19/07/2021).
Chingala, G., et al., 2017: Determinants of smallholder farmers’ perceptions of impact of climate change on beef production in Malawi. Clim Change, 142 (1-2), 129–141, doi:10.1007/s10584-017-1924-1.
Chirwa, P.W., L. Mahamane and G. Kowero, 2017: Forests, people and environment: some African perspectives. South. For. , 79 (2), 79–85, doi:10.2989/20702620.2017.1295347.
Chitale, V., R. Silwal and M. Matin, 2018: Assessing the impacts of climate change on distribution of major non-timber forest plants in Chitwan Annapurna Landscape, Nepal. Resources, 7 (4), 66, doi:10.3390/resources7040066.
Cho, S.J. and B.A. McCarl, 2017: Climate change influences on crop mix shifts in the United States. Sci Rep, 7, 40845, doi:10.1038/srep40845.
Christen, B. and T. Dalgaard, 2013: Buffers for biomass production in temperate European agriculture: A review and synthesis on function, ecosystem services and implementation. Biomass Bioenergy, 55, 53–67, doi:10.1016/j.biombioe.2012.09.053.
Churkina, G., et al., 2020: Buildings as a global carbon sink. Nat. Sustain. , 3 (4), 269–276, doi:10.1038/s41893-019-0462-4.
Chuwah, C., et al., 2015: Global impacts of surface ozone changes on crop yields and land use. Atmos. Environ. , 106, 11–23, doi:10.1016/j.atmosenv.2015.01.062.
Cilas, C. and P. Bastide, 2020: Challenges to Cocoa Production in the Face of Climate Change and the Spread of Pests and Diseases. Agronomy, 10 (9), doi:10.3390/agronomy10091232.
Cisneros-Montemayor, A.M., G. Ishimura, G.R. Munro and U.R. Sumaila, 2020: Ecosystem-based management can contribute to cooperation in transboundary fisheries: the case of pacific sardine. Fish. Res. , 221, 105401, doi:10.1016/j.fishres.2019.105401.
Cisneros-Montemayor, A.M., D. Pauly, L.V. Weatherdon and Y. Ota, 2016: A global estimate of seafood consumption by coastal indigenous peoples. PLoS ONE, 11 (12), e166681, doi:10.1371/journal.pone.0166681.
Claeys, F., et al., 2019: Climate change would lead to a sharp acceleration of Central African forests dynamics by the end of the century. Environ. Res. Lett. , 14 (4), doi:10.1088/1748-9326/aafb81.
Claeys, P. and D. Delgado Pugley, 2017: Peasant and indigenous transnational social movements engaging with climate justice. Can. J. Dev. Stud. / Revue Can. D’études Du Développement , 38 (3), 325–340, doi:10.1080/02255189.2016.1235018.
Clapp, J., 2014: Financialization, distance and global food politics. J Peasant Stud, 41 (5), 797–814, doi:10.1080/03066150.2013.875536.
Clapp, J., 2017: Responsibility to the rescue? Governing private financial investment in global agriculture. Agric Hum Values, 34 (1), 223–235, doi:10.1007/s10460-015-9678-8.
Clapp, J. and S.R. Isakson, 2018: Risky returns: the implications of financialization in the food system. Dev Change, 49 (2), 437–460, doi:10.1111/dech.12376.
Clark, M. A., M. Springmann, J. Hill and D. Tilman, 2019: Multiple health and environmental impacts of foods. Proc. Natl. Acad. Sci. U. S. A. , 116 (46), 23357–23362, doi:10.1073/pnas.1906908116.
Clay, L.A., M. A. Papas, K.B. Gill and D.M. Abramson, 2018: Factors associated with continued food insecurity among households recovering from Hurricane Katrina. Int J Environ Res Public Health, 15 (8), 1647, doi:10.3390/ijerph15081647.
Clay, N. and B. King, 2019: Smallholders’ uneven capacities to adapt to climate change amid Africa’s ‘green revolution’: case study of Rwanda’s crop intensification program. World Dev, 116, 1–14, doi:10.1016/j.worlddev.2018.11.022.
Clement, A., et al., 2016: Exceptional summer conditions and HABs of Pseudochattonella in Southern Chile create record impacts on salmon farms. Harmful Algae News, 53, 1–3.
Clinton, N., et al., 2018: A global geospatial ecosystem services estimate of urban agriculture. Earths Future, 6 (1), 40–60, doi:10.1002/2017ef000536.
Cobb, J.N., et al., 2019: Enhancing the rate of genetic gain in public-sector plant breeding programs: lessons from the breeder’s equation. Theor. Appl. Genet. , 132 (3), 627–645, doi:10.1007/s00122-019-03317-0.
Cochrane, K.L., et al., 2019: Tools to enrich vulnerability assessment and adaptation planning for coastal communities in data-poor regions: application to a case study in Madagascar. Front. Mar. Sci. , 5, 505, doi:10.3389/fmars.2018.00505.
Coe, R., F. Sinclair and E. Barrios, 2014: Scaling up agroforestry requires research ‘in’ rather than ‘for’ development. Curr. Opin. Environ. Sustain. , 6, 73–77, doi:10.1016/j.cosust.2013.10.013.
Cohen, A., et al., 2018: Combined fish and lettuce cultivation: an aquaponics life cycle assessment. [Laurent, A., et al.(ed.)]. 25th CIRP Life Cycle Engineering (LCE) Conference, 30 April - 2 May 2018, Copenhagen, Denmark. 69, Elsevier, In, pp. 551–556. doi:10.1016/j.procir.2017.11.029.
Cohen, A.S., et al., 2016: Climate warming reduces fish production and benthic habitat in Lake Tanganyika, one of the most biodiverse freshwater ecosystems. Proc. Natl. Acad. Sci. U. S. A. , 113 (34), 9563–9568, doi:10.1073/pnas.1603237113.
Cohen, M.J., 2020: Does the COVID-19 outbreak mark the onset of a sustainable consumption transition?Sustain. Sci. Pract. Policy, 16 (1), 1–3, doi:10.1080/15487733.2020.1740472.
Cold, H.S., et al., 2020: Assessing vulnerability of subsistence travel to effects of environmental change in Interior Alaska. Ecol. Soc. , 25 (1), 20, doi:10.5751/Es-11426-250120.
Coleman, M. A., et al., 2019: Climate change does not affect the seafood quality of a commonly targeted fish. Glob Chang Biol, 25 (2), 699–707, doi:10.1111/gcb.14513.
Colfer, C.J.P. (ed.), 2008: The Longhouse of the Tarsier: Changing Landscape, Gender, and Well Being in Borneo. Monograph Series, Vol. 10. Borneo Research Council, Phillips, ME, USA, ISBN 978-1929900104. 418 pp.
Collier, R.J. and K.G. Gebremedhin, 2015: Thermal biology of domestic animals. Annu. Rev. Anim. Biosci. , 3 (1), 513–532, doi:10.1146/annurev-animal-022114-110659.
Collins, A., 2018: Saying all the right things? Gendered discourse in climate-smart agriculture. J Peasant Stud, 45, 175–191, doi:10.1080/03066150.2017.1377187.
Collste, D., et al., 2019: A cross-scale participatory approach to discuss pathways to the 2030 Agenda SDGs: the example of The World In 2050 African Dialogues. A Methodology paper on the The Three Horizons Framework for the SDGs (3H4SDG). doi:10.31235/osf.io/uhskb.
Colombo, S.M., et al., 2020: Projected declines in global DHA availability for human consumption as a result of global warming. Ambio, 49 (4), 865–880, doi:10.1007/s13280-019-01234-6.
Cominassi, L., et al., 2020: Food availability modulates the combined effects of ocean acidification and warming on fish growth. Sci Rep, 10 (1), 2338, doi:10.1038/s41598-020-58846-2.
Comte, L. and G. Grenouillet, 2015: Distribution shifts of freshwater fish under a variable climate: comparing climatic, bioclimatic and biotic velocities. Divers. Distrib. , 21 (9), 1014–1026.
Comte, L. and J.D. Olden, 2017: Climatic vulnerability of the world’s freshwater and marine fishes. Nat. Clim. Chang. , 7 (10), 718–722, doi:10.1038/Nclimate3382.
Conigliani, C., N. Cuffaro and G. D’Agostino, 2018: Large-scale land investments and forests in Africa. Land Use Policy, 75 (February 2017), 651–660, doi:10.1016/j.landusepol.2018.02.005.
Conijn, J.G., P.S. Bindraban, J.J. Schröder and R.E.E. Jongschaap, 2018: Can our global food system meet food demand within planetary boundaries?Agric. Ecosyst. Environ. , 251, 244–256, doi:10.1016/j.agee.2017.06.001.
Conway, D., et al., 2015: Climate and southern Africa’s water-energy-food nexus. Nat. Clim. Chang. , 5 (9), 837–846, doi:10.1038/Nclimate2735.
Cook, B.I. and E.M. Wolkovich, 2016: Climate change decouples drought from early wine grape harvests in France. Nat. Clim. Change, 6 (7), 715–719, doi:10.1038/Nclimate2960.
Cook, N.J., T. Grillos and K.P. Andersson, 2019: Gender quotas increase the equality and effectiveness of climate policy interventions. Nat. Clim. Chang. , 9 (4), 330–334, doi:10.1038/s41558-019-0438-4.
Cooley, S.R., C.R. Ono, S. Melcer and J. Roberson, 2016: Community-level actions that can address ocean acidification. Front. Mar. Sci. , 2, 128, doi:10.3389/fmars.2015.00128.
Cooley, S.R., et al., 2015: An integrated assessment model for helping the United States sea scallop (Placopecten magellanicus) fishery plan ahead for ocean acidification and warming. PLoS ONE, 10 (5), e124145, doi:10.1371/journal.pone.0124145.
Cooper, M.W., et al., 2019: Mapping the effects of drought on child stunting. Proc. Natl. Acad. Sci. U. S. A. , 116 (35), 17219–17224, doi:10.1073/pnas.1905228116.
Cooper, O.R., et al., 2014: Global distribution and trends of tropospheric ozone: an observation-based review. Elem. Sci. Anthropocene, 2, 29, doi:10.12952/journal.elementa.000029.
Coq-Huelva, D., et al., 2017: Co-evolution and bio-social construction: the Kichwa agroforestry systems (Chakras) in the Ecuadorian Amazonia. Sustainability, 9 (10), 1920, doi:10.3390/su9101920.
Corbeels, M., et al., 2019: The 4 per 1000 goal and soil carbon storage under agroforestry and conservation agriculture systems in sub-Saharan Africa. Soil Tillage Res. , 188, 16–26, doi:10.1016/j.still.2018.02.015.
Corbeels, M., et al., 2020: Carbon sequestration potential through conservation agriculture in Africa has been largely overestimated comment on: “Meta-analysis on carbon sequestration through conservation agriculture in Africa”. Soil Tillage Res. , 196, 104300, doi:10.1016/j.still.2019.104300.
Corbera, E., C. Hunsberger and C. Vaddhanaphuti, 2017: Climate change policies, land grabbing and conflict: perspectives from Southeast Asia. Can. J. Dev. Stud. , 38 (3), 297–304, doi:10.1080/02255189.2017.1343413.
Córdova, R., N.J. Hogarth and M. Kanninen, 2019: Mountain farming systems’ exposure and sensitivity to climate change and variability: agroforestry and conventional agriculture systems compared in Ecuador’s indigenous territory of Kayambi people. Sustainability, 11 (9), doi:10.3390/su11092623.
Cormier-Salem, M.C. and J. Panfili, 2016: Mangrove reforestation: greening or grabbing coastal zones and deltas? Case studies in Senegal. Afr. J. Aquat. Sci. , 41 (1), 89–98, doi:10.2989/16085914.2016.1146122.
Cornwall, W., 2019: In hot water: Five years ago, a marine heat wave began to wash across Pacific ecosystems. Researchers fear it is a preview of the oceans’ future. Science, 363 (6426), 442–445, doi:10.1126/science.363.6426.442.
Costa, R., et al., 2019: Grapevine phenology of cv. Touriga Franca and Touriga Nacional in the Douro wine region: modelling and climate change projections. Agronomy, 9 (4), 210, doi:10.3390/agronomy9040210.
Costella, C., et al., 2017: Scalable and Sustainable: How to Build Anticipatory Capacity into Social Protection Systems. IDS Bull. , 48 (4), doi:10.19088/1968-2017.151.
Costello, C., et al., 2020: The future of food from the sea. Nature, 588 (7836), 95–100, doi:10.1038/s41586-020-2616-y.
Costinot, A., D. Donaldson and C. Smith, 2016: Evolving comparative advantage and the impact of climate change in agricultural markets: evidence from 1.7 million fields around the world. J. Polit. Econ. , 124 (1), 205–248, doi:10.1086/684719.
Côte, F.-X., et al., 2019: The agroecological transition of agricultural systems in the Global South. éditions Quae, ISBN 978-2759230570.
Cottrell, R.S., et al., 2020: Global adoption of novel aquaculture feeds could substantially reduce forage fish demand by 2030. Nat. Food, 1 (5), 301–308, doi:10.1038/s43016-020-0078-x.
Cottrell, R.S., et al., 2018: Considering land-sea interactions and trade-offs for food and biodiversity. Glob Chang Biol, 24 (2), 580–596, doi:10.1111/gcb.13873.
Cottrell, R.S., et al., 2019: Food production shocks across land and sea. Nat. Sustain. , 2 (2), 130–137, doi:10.1038/s41893-018-0210-1.
Cotula, L., 2012: The international political economy of the global land rush: a critical appraisal of trends, scale, geography and drivers. J Peasant Stud, 39 (3-4), 649–680, doi:10.1080/03066150.2012.674940.
Coughlan de Perez, E., et al., 2016: Action-based flood forecasting for triggering humanitarian action. Hydrol. Earth Syst. Sci. , 20 (9), 3549–3560, doi:10.5194/hess-20-3549-2016.
Coulibaly, J.Y., B. Chiputwa, T. Nakelse and G. Kundhlande, 2017: Adoption of agroforestry and the impact on household food security among farmers in Malawi. Agric Syst , 155, 52–69, doi:10.1016/j.agsy.2017.03.017.
Cozzetto, K., et al., 2013: Climate change impacts on the water resources of American Indians and Alaska Natives in the US. In: Climate Change and Indigenous Peoples in the United States: Impacts, Experiences, and Action[Maldonado, J.K., B. Colombi and R. Pandya(eds.)]. Springer, Cham, Switzerland, pp. 61–76. ISBN 978-3319052656.
Cradock-Henry, N.A., et al., 2020: Climate adaptation pathways for agriculture: Insights from a participatory process. Environ Sci Policy, 107, 66–79, doi:10.1016/j.envsci.2020.02.020.
Craparo, A.C.W., et al., 2015: Coffea arabica yields decline in Tanzania due to climate change: Global implications. Agric For Meteorol, 207, 1–10, doi:10.1016/j.agrformet.2015.03.005.
Crespo-Perez, V., et al., 2015: Changes in the distribution of multispecies pest assemblages affect levels of crop damage in warming tropical Andes. Glob Chang Biol, 21 (1), 82–96, doi:10.1111/gcb.12656.
Cristina Rulli, M. and P. D’Odorico, 2014: Food appropriation through large scale land acquisitions. Environ. Res. Lett. , 9 (6), doi:10.1088/1748-9326/9/6/064030.
Cross, J.N., et al., 2019: Building the knowledge-to-action pipeline in North America: connecting ocean acidification research and actionable decision support. Front. Mar. Sci. , 6, 356, doi:10.3389/fmars.2019.00356.
Crous-Duran, J., et al., 2019: Assessing food sustainable intensification potential of agroforestry using a carbon balance method. iForest - Biogeosciences For. , 12 (1), 85–91, doi:10.3832/ifor2578-011.
Cubins, J.A., et al., 2019: Management of pennycress as a winter annual cash cover crop. A review. Agron. Sustain. Dev. , 39 (5), 1–11, doi:10.1007/S13593-019-0592-0.
Cuccia, C., et al., 2014: Phenological model performance to warmer conditions: application to Pinot noir in Burgundy. Oeno One, 48 (3), doi:10.20870/oeno-one.2014.48.3.1572.
Cummings, V.J., et al., 2019: Effect of reduced pH on physiology and shell integrity of juvenile Haliotis iris (paua) from New Zealand. Peer J, 7 (9), e7670, doi:10.7717/peerj.7670.
Cunsolo Willox, A., et al., 2012: “From this place and of this place:” climate change, sense of place, and health in Nunatsiavut, Canada. Soc. Sci. Med. , 75 (3), 538–547, doi:10.1016/j.socscimed.2012.03.043.
Curtis, P.G., et al., 2018: Classifying drivers of global forest loss. Science, 361 (6407), 1108–1111, doi:10.1126/science.aau3445.
Custódio, M., S. Villasante, R. Calado and A.I. Lillebø, 2020: Valuation of ecosystem services to promote sustainable aquaculture practices. Rev. Aquac. , 12 (1), 392–405, doi:10.1111/raq.12324.
D’agata, S., et al., 2020: Multiscale determinants of social adaptive capacity in small-scale fishing communities. Environ Sci Policy, 108, 56–66, doi:10.1016/j.envsci.2020.03.006.
D’Annolfo, R., B. Gemmill-Herren, B. Graeub and L.A. Garibaldi, 2017: A review of social and economic performance of agroecology. Int. J. Agric. Sustain. , 15 (6), 632–644, doi:10.1080/14735903.2017.1398123.
D’Odorico, P., et al., 2018: The global food-energy-water nexus. Rev. Geophys. , 56 (3), 456–531, doi:10.1029/2017rg000591.
D’Odorico, P., M.C. Rulli, J. Dell’Angelo and K.F. Davis, 2017: New frontiers of land and water commodification: socio-environmental controversies of large-scale land acquisitions. Land Degrad Dev, 28 (7), 2234–2244, doi:10.1002/ldr.2750.
da Fonseca de Oliveira, A.C., et al., 2019: Impacts on performance of growing-finishing pigs under heat stress conditions: a meta-analysis. Vet. Res. Commun. , 43 (1), 37–43, doi:10.1007/s11259-018-9741-1.
Dabbadie, L., et al., 2018: Effects of climate change on aquaculture: drivers, impacts and policies. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 449–463. ISBN 978-9251306079.
Daftary-Steel, S., H. Herrera and C.M. Porter, 2015: The unattainable trifecta of urban agriculture. J. Agric. Food Syst. Community Dev. , 6 (1), 19–32, doi:10.5304/jafscd.2015.061.014.
Dainese, M., et al., 2019: A global synthesis reveals biodiversity-mediated benefits for crop production. Sci. Adv. , 5 (10), eaax121, doi:10.1126/sciadv.aax0121.
Daliakopoulos, I.N., et al., 2016: The threat of soil salinity: a European scale review. Sci. Total Environ. , 573, 727–739, doi:10.1016/j.scitotenv.2016.08.177.
Dalin, C., Y. Wada, T. Kastner and M.J. Puma, 2017: Groundwater depletion embedded in international food trade. Nature, 543 (7647), 700–704, doi:10.1038/nature21403.
Dalu, T., R.J. Wasserman and M.T.B. Dalu, 2017: Agricultural intensification and drought frequency increases may have landscape-level consequences for ephemeral ecosystems. Glob. Chang. Biol. , 23 (3), 983–985, doi:10.1111/gcb.13549.
Daron, J., et al., 2021: Integrating seasonal climate forecasts into adaptive social protection in the Sahel. Clim. Dev. , 13 (6), 543–550, doi:10.1080/17565529.2020.1825920.
Darrouzet-Nardi, et al., 2016: Child dietary quality in rural Nepal: effectiveness of a community-led development initiative. Food Policy, 61, 185–197.
Daryanto, S., L.X. Wang and P.A. Jacinthe, 2017: Global synthesis of drought effects on cereal, legume, tuber and root crops production: a review. Agric. Water Manag. , 179, 18–33, doi:10.1016/j.agwat.2016.04.022.
Das, I., et al., 2020: Effects of climate change and management policies on marine fisheries productivity in the north-east coast of India. Sci. Total Environ. , 724, 138082, doi:10.1016/j.scitotenv.2020.138082.
Das, M., V. Jain and S.K. Malhotra, 2016a: Impact of climate change on medicinal and aromatic plants: Review. Indian J. Agric. Sci. , 86 (11), 1375–1382.
Das, R., et al., 2016b: Impact of heat stress on health and performance of dairy animals: a review. Vet World, 9 (3), 260–268, doi:10.14202/vetworld.2016.260-268.
Dasgupta, P., J.F. Morton, D. Dodman, B. Karapinar, F. Meza, M.G. Rivera-Ferre, A. Toure Sarr and K.E. Vincent, 2014: Rural Areas. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. [Field, C. B., V. R. Barros, D. J. Dokken, K. J. Mach, M. D. Mastrandrea, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea and L. L. White (eds.)]. Cambridge University Press, In, pp. 613–657. ISBN 978-1107058071 Cambridge, UK, and New York, NY, USA).
Datta, S., et al., 2018: Aquaponics: a green and sustainable eco-tech for environmental cum economic benefits through integration of fish and edible crop cultivation. In: Wastewater Management Through Aquaculture[Jana, B.B., R.N. Mandal and P. Jayasankar(eds.)]. Springer Nature, Singapore, Singapore, pp. 207–224. ISBN 978-9811072475.
Dauber, J. and S. Miyake, 2016: To integrate or to segregate food crop and energy crop cultivation at the landscape scale? Perspectives on biodiversity conservation in agriculture in Europe. Energ Sustain Soc, 6 (1), 25–25, doi:10.1186/s13705-016-0089-5.
Daugstad, K., K. Rønningen and B. Skar, 2006: Agriculture as an upholder of cultural heritage? Conceptualizations and value judgements—a Norwegian perspective in international context. J Rural Stud, 22 (1), 67–81, doi:10.1016/j.jrurstud.2005.06.002.
Dave, D. and W. Routray, 2018: Current scenario of Canadian fishery and corresponding underutilized species and fishery byproducts: a potential source of omega-3 fatty acids. J Clean Prod, 180, 617–641, doi:10.1016/j.jclepro.2018.01.091.
David, A.T., J.E. Asarian and F.K. Lake, 2018: Wildfire Smoke Cools Summer River and Stream Water Temperatures. Water Resour. Res. , 54 (10), 7273–7290, doi:10.1029/2018wr022964.
Davis, H., et al., 2017: The economic potential of fruit trees as shade in blue mountain coffee agroecosystems of the Yallahs River watershed, Jamaica W.I. Agrofor. Syst. , 93 (2), 581–589, doi:10.1007/s10457-017-0152-z.
Davis, K.F., P. D’Odorico and M.C. Rulli, 2014: Land grabbing: a preliminary quantification of economic impacts on rural livelihoods. Popul Environ, 36 (2), 180–192, doi:10.1007/s11111-014-0215-2.
Davis, K.F., S. Downs and J.A. Gephart, 2021: Towards food supply chain resilience to environmental shocks. Nat. Food, 2 (1), 54–65, doi:10.1038/s43016-020-00196-3.
Davis, K.F., et al., 2020: Tropical forest loss enhanced by large-scale land acquisitions. Nat. Geosci. , 13 (7), 482–488, doi:10.1038/s41561-020-0592-3.
Davis, K.F., M.C. Rulli and P. D’Odorico, 2015a: The global land rush and climate change. Earth’s Future, 3 (8), 298–311, doi:10.1002/2014EF000281.
Davis, K.F., et al., 2015b: Accelerated deforestation driven by large-scale land acquisitions in Cambodia. Nat Geosci, 8 (10), 772, doi:10.1038/Ngeo2540.
Davis, K.T., et al., 2019: Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc. Natl. Acad. Sci. USA, 116 (13), 6193–6198, doi:10.1073/pnas.1815107116.
Davis, M., 2001: Late Victorian Holocausts: El Niño Famines and the Making of the Third World. Verso, London, UK, and Brooklyn, NY, USA, ISBN 978-1781680612.
Day, E., et al., 2019: Upholding labour productivity under climate change: an assessment of adaptation options. Clim Policy, 19 (3), 367–385, doi:10.1080/14693062.2018.1517640.
Dayamba, D.S., et al., 2018: Assessment of the use of participatory integrated climate services for agriculture (PICSA) approach by farmers to manage climate risk in Mali and Senegal. Clim. Serv. , 12, 27–35, doi:10.1016/j.cliser.2018.07.003.
de Albuquerque Nunes, P.A., et al., 2021: Livestock integration into soybean systems improves long-term system stability and profits without compromising crop yields. Sci Rep, 11 (1), 1649, doi:10.1038/s41598-021-81270-z.
de Amorim, W.S., et al., 2018: The nexus between water, energy, and food in the context of the global risks: an analysis of the interactions between food, water, and energy security. Environ Impact Assess Rev, 72, 1–11, doi:10.1016/j.eiar.2018.05.002.
de Boer, H.J. and C. Cotingting, 2014: Medicinal plants for women’s healthcare in southeast Asia: a meta-analysis of their traditional use, chemical constituents, and pharmacology. J. Ethnopharmacol. , 151 (2), 747–767, doi:10.1016/j.jep.2013.11.030.
de Boer, I.J.M. and M.K. van Ittersum, 2018: Circularity in Agricultural Production. Wageningen University and Research, https://library.wur.nl/WebQuery/wurpubs/fulltext/470625. Accessed 2021 . (74 pp).
de Bruyn, J., et al., 2018: Does village chicken-keeping contribute to young children’s diets and growth? A longitudinal observational study in rural Tanzania. Nutrients, 10 (11), 1799, doi:10.3390/nu10111799.
de Coninck, H., A. Revi, M. Babiker, P. Bertoldi, M. Buckeridge, A. Cartwright, W. Dong, J. Ford, S. Fuss, J.-C. Hourcade, D. Ley, R. Mechler, P. Newman, A. Revokatova, S. Schultz, L. Steg, and T. Sugiyama, 2018: Strengthening and Implementing the Global Response. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty[Masson-Delmotte, V., P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P.R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J.B.R. Matthews, Y. Chen, X. Zhou, M.I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, and T. Waterfield (eds.)]. ISBN 978-9291691517 In press.
De Frenne, P., et al., 2013: Microclimate moderates plant responses to macroclimate warming. Proc. Natl. Acad. Sci. U. S. A. , 110 (46), 18561–18565, doi:10.1073/pnas.1311190110.
de Leeuw, J., et al., 2020: The pastoral farming system: balancing between tradition and transition. In: Farming Systems and Food Security in Africa: Priorities for Science and Policy under Global Change[Dixon, J., et al.(ed.)]. Routledge, Oxon, UK, and New York, NY, USA, pp. 318–353. ISBN 978-1138963351.
de Lima, C.Z., et al., 2021: Heat stress on agricultural workers exacerbates crop impacts of climate change. Environ. Res. Lett. , 16 (4), 44020, doi:10.1088/1748-9326/abeb9 f.
de Melo, C.M.R., E. Durland and C. Langdon, 2016: Improvements in desirable traits of the Pacific oyster, Crassostrea gigas, as a result of five generations of selection on the West Coast, USA. Aquaculture, 460, 105–115, doi:10.1016/j.aquaculture.2016.04.017.
de Miguel, Á., A.Y. Hoekstra and E. García-Calvo, 2015: Sustainability of the water footprint of the Spanish pork industry. Ecol Indic, 57, 465–474, doi:10.1016/j.ecolind.2015.05.023.
de Sousa, K., et al., 2019: The future of coffee and cocoa agroforestry in a warmer Mesoamerica. Sci Rep, 9 (1), 8828, doi:10.1038/s41598-019-45491-7.
de Vries, F.T., et al., 2020: Harnessing rhizosphere microbiomes for drought-resilient crop production. Science, 368 (6488), 270–274, doi:10.1126/science.aaz5192.
De Zeeuw, H., R. Van Veenhuizen and M. Dubbeling, 2011: The role of urban agriculture in building resilient cities in developing countries. J. Agric. Sci. , 149 (S1), 153–163, doi:10.1017/s0021859610001279.
Deb, J.C., S. Phinn, N. Butt and C.A. McAlpine, 2018: Climate change impacts on tropical forests: identifying risks for tropical Asia. J. Trop. For. Sci. , 30 (2), 182–194, doi:10.26525/jtfs2018.30.2.182194.
DeBano, S.J., et al., 2016: Interactive influences of climate change and agriculture on aquatic habitat in a Pacific Northwestern watershed. Ecosphere, 7 (6), e1357, doi:10.1002/ecs2.1357.
Debonne, N., J. van Vliet and P. Verburg, 2019: Future governance options for large-scale land acquisition in Cambodia: Impacts on tree cover and tiger landscapes. Environ Sci Policy, 94 (December 2018), 9–19, doi:10.1016/j.envsci.2018.12.031.
Deeb, R., et al., 2018: Impact of climate change on Vibrio vulnificus abundance and exposure risk. Estuaries Coasts, 41 (8), 2289–2303, doi:10.1007/s12237-018-0424-5.
Degani, E., et al., 2019: Crop rotations in a climate change scenario: short-term effects of crop diversity on resilience and ecosystem service provision under drought. Agric. Ecosyst. Environ. , 285, 106625, doi:10.1016/j.agee.2019.106625.
Dégremont, L., et al., 2015a: New insight for the genetic evaluation of resistance to ostreid herpesvirus infection, a worldwide disease, in Crassostrea gigas. PLoS ONE, 10 (6), e127917, doi:10.1371/journal.pone.0127917.
Dégremont, L., M. Nourry and E. Maurouard, 2015b: Mass selection for survival and resistance to OsHV-1 infection in Crassostrea gigas spat in field conditions: response to selection after four generations. Aquaculture, 446, 111–121, doi:10.1016/j.aquaculture.2015.04.029.
Dehm, J., 2016: Indigenous peoples and REDD+ safeguards: rights as resistance or as disciplinary inclusion in the green economy?J. Hum. Rights Environ. , 7 (2), 170–217, doi:10.4337/jhre.2016.02.01.
del Pozo, A., et al., 2019: Climate change impacts and adaptation strategies of agriculture in Mediterranean-climate regions (MCRs). Sustainability, 11 (10), 2769–2769, doi:10.3390/su11102769.
Delang, C.O., 2006: Not just minor forest products: the economic rationale for the consumption of wild food plants by subsistence farmers. Ecol. Econ. , 59 (1), 64–73, doi:10.1016/j.ecolecon.2005.10.006.
Delcour, I., P. Spanoghe and M. Uyttendaele, 2015: Literature review: impact of climate change on pesticide use. Food. Res. Int. , 68, 7–15, doi:10.1016/j.foodres.2014.09.030.
Delesalle, V.A. and S. Blum, 1994: Variation in germination and survival among families of Sagittaria-Latifolia in response to salinity and temperature. Int. J. Plant Sci. , 155 (2), 187–195, doi:10.1086/297158.
Dell’Angelo, J., P. D’Odorico and M.C. Rulli, 2017: Threats to sustainable development posed by land and water grabbing. Curr. Opin. Environ. Sustain. , 26-27 (June), 120–128, doi:10.1016/j.cosust.2017.07.007.
Dell’Angelo, J., M.C. Rulli and P. D’Odorico, 2018: The global water grabbing syndrome. Ecol. Econ. , 143, 276–285, doi:10.1016/j.ecolecon.2017.06.033.
Dell’Angelo, J., P. D’Odorico, M.C. Rulli and P. Marchand, 2016: The Tragedy of the Grabbed Commons: Coercion and Dispossession in the Global Land Rush. World Dev, xx, doi:10.1016/j.worlddev.2016.11.005.
Dell’Angelo, J., et al., 2021: Commons grabbing and agribusiness : Violence , resistance and social mobilization. Ecol. Econ. , 184, 107004–107004, doi:10.1016/j.ecolecon.2021.107004.
Dellar, M., C.F.E. Topp, G. Banos and E. Wall, 2018: A meta-analysis on the effects of climate change on the yield and quality of European pastures. Agric Ecosyst Environ, 265, 413–420, doi:10.1016/j.agee.2018.06.029.
DeLonge, M.S., A. Miles and L. Carlisle, 2016: Investing in the transition to sustainable agriculture. Environ Sci Policy, 55, 266–273, doi:10.1016/j.envsci.2015.09.013.
Delzeit, R., F. Zabel, C. Meyer and T. Václavík, 2017: Addressing future trade-offs between biodiversity and cropland expansion to improve food security. Reg Environ Change, 17 (5), 1429–1441, doi:10.1007/s10113-016-0927-1.
Demeke, M., et al., 2017: Linking Farm Diversification to Household Diet Diversification: Evidence from a Sample of Kenyan Ultrapoor Farmers. ESA Working Paper. Food and Agriculture Organisation of the United Nations, Rome, Italy, http://www.fao.org/3/i6852e/i6852e.pdf. Accessed 2021 . (27 pp).
Dempewolf, H., et al., 2014: Adapting Agriculture to Climate Change: A Global Initiative to Collect, Conserve, and Use Crop Wild Relatives. Agroecol. Sustain. Food Syst. , 38 (4), 369–377, doi:10.1080/21683565.2013.870629.
Denton, F., T.J. Wilbanks, A.C. Abeysinghe, I. Burton, Q. Gao, M.C. Lemos, T. Masui, K.L. O’Brien, and K. Warner, 2014: Climate-resilient pathways: adaptation, mitigation, and sustainable development. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[Field, C. B., V.R. Barros, D.J. Dokken, K.J. Mach, M.D. Mastrandrea, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L. White (ed.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 1101-1131.
Department of Environmental Affairs, 2016: South Africa National Adaptation Strategy (draft). Department of Environmental Affairs, Republic of South Africa, 122 pp. Available at: https://www.dffe.gov.za/sites/default/files/docs/nas2016.pdf (accessed June 15, 2021).
Derner, J., et al., 2018: Vulnerability of grazing and confined livestock in the Northern Great Plains to projected mid- and late-twenty-first century climate. Clim Change, 146 (1-2), 19–32, doi:10.1007/s10584-017-2029-6.
Deryng, D., et al., 2016: Regional disparities in the beneficial effects of rising CO 2 concentrations on crop water productivity. Nat. Clim. Chang. , 6 (8), 786, doi:10.1038/nclimate2995.
Des, M., et al., 2020: How can ocean warming at the NW Iberian Peninsula affect mussel aquaculture?Sci. Total Environ. , 709, 136117, doi:10.1016/j.scitotenv.2019.136117.
Descheemaeker, K., et al., 2018: Effects of climate change and adaptation on the livestock component of mixed farming systems: a modelling study from semi-arid Zimbabwe. Agric Syst , 159, 282–295, doi:10.1016/j.agsy.2017.05.004.
Deur, D., A. Dick, K. Recalma-Clutesi and N.J. Turner, 2015: Kwakwaka’wakw “clam gardens”. Hum Ecol, 43 (2), 201–212.
Deutsch, C.A., et al., 2018: Increase in crop losses to insect pests in a warming climate. Science, 361 (6405), 916–919, doi:10.1126/science.aat3466.
Dhakal, B. and R.R. Kattel, 2019: Effects of global changes on ecosystems services of multiple natural resources in mountain agricultural landscapes. Sci. Total Environ. , 676, 665–682, doi:10.1016/j.scitotenv.2019.04.276.
Dhyani, A., R. Kadaverugu, B.P. Nautiyal and M.C. Nautiyal, 2021: Predicting the potential distribution of a critically endangered medicinal plant Lilium polyphyllum in Indian Western Himalayan Region. Reg Environ Change, 21 (2), 30, doi:10.1007/s10113-021-01763-5.
Di Leo, N., F.J. Escobedo and M. Dubbeling, 2016: The role of urban green infrastructure in mitigating land surface temperature in Bobo-Dioulasso, Burkina Faso. Environ. Dev. Sustain. , 18 (2), 373–392, doi:10.1007/s10668-015-9653-y.
Diacono, M., et al., 2017: Agro-ecology for potential adaptation of horticultural systems to climate change: agronomic and energetic performance evaluation. Agronomy, 7 (2), 35, doi:10.3390/agronomy7020035.
Diaz, D. and F. Moore, 2017: Quantifying the economic risks of climate change. Nat. Clim. Chang. , 7 (11), 774–782, doi:10.1038/nclimate3411.
Diekert, F.K. and E. Nieminen, 2017: International fisheries agreements with a shifting stock. Dyn. Games Appl. , 7 (2), 185–211, doi:10.1007/s13235-016-0184-4.
Ding, Q., X.J. Chen, R. Hilborn and Y. Chen, 2017: Vulnerability to impacts of climate change on marine fisheries and food security. Mar. Policy. , 83, 55–61, doi:10.1016/j.marpol.2017.05.011.
DiTomaso, J.M., T.A. Monaco, J.J. James and J. Firn, 2017: Invasive Plant Species and Novel Rangeland Systems. In: Rangeland Systems: Processes, Management and Challenges[Briske, D.D.(ed.)]. Springer International Publishing, Cham, pp. 429–465. ISBN 978-3319467092.
Djoudi, H., M. Brockhaus and B. Locatelli, 2013: Once there was a lake: vulnerability to environmental changes in northern Mali. Reg Environ Change, 13 (3), 493–508, doi:10.1007/s10113-011-0262-5.
Dobardzic, S., et al., 2019: 2019 State of Climate Services: Agriculture and Food Security. World Meteorological Organization (WMO), Geneva, Switzerland, ISBN 978-9263112422. 44 pp.
Doblas-Reyes, F.J., A.A. Sörensson, M. Almazroui, A. Dosio, W.J. Gutowski, R. Haarsma, R. Hamdi, B. Hewitson, W.-T. Kwon, B.L. Lamptey, D. Maraun, T.S. Stephenson, I. Takayabu, L. Terray, A. Turner, and Z. Zuo, 2021: Linking Global to Regional Climate Change. In: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Chang[Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press.
Doddabasawa, B.M. Chittapur and M.M. Murthy, 2018: Traditional agroforestry systems and biodiversity conservation. Bangladesh J. Bot. , 47 (4), 927–935, doi:10.3329/bjb.v47i4.47388.
Doelman, J.C., et al., 2018: Exploring SSP land-use dynamics using the IMAGE model: regional and gridded scenarios of land-use change and land-based climate change mitigation. Glob. Environ. Change. , 48, 119–135, doi:10.1016/j.gloenvcha.2017.11.014.
Doetterl, S., et al., 2015: Soil carbon storage controlled by interactions between geochemistry and climate. Nat Geosci, 8 (10), 780–783, doi:10.1038/Ngeo2516.
Doherty, E., et al., 2021: A discrete choice experiment exploring farmer preferences for insurance against extreme weather events. J. Environ. Manag. , 290, 112607, doi:10.1016/j.jenvman.2021.112607.
Doherty, R.M., et al., 2010: Implications of future climate and atmospheric CO2content for regional biogeochemistry, biogeography and ecosystem services across East Africa. Glob. Chang. Biol. , 16 (2), 617–640, doi:10.1111/j.1365-2486.2009.01997.x.
Donatti, C.I., et al., 2019: Vulnerability of smallholder farmers to climate change in Central America and Mexico: current knowledge and research gaps. Clim. Dev. , 11 (3), 264–286, doi:10.1080/17565529.2018.1442796.
Doney, S.C., D.S. Busch, S.R. Cooley and K.J. Kroeker, 2020: The Impacts of Ocean Acidification on Marine Ecosystems and Reliant Human Communities. Annu. Rev. Environ. Resour. , 45 (1), 83–112, doi:10.1146/annurev-environ-012320-083019.
Dong, Z.Q., et al., 2018: A quantitative method for risk assessment of agriculture due to climate change. Theor Appl Climatol, 131 (1-2), 653–659, doi:10.1007/s00704-016-1988-2.
Dono, G., et al., 2016: Winners and losers from climate change in agriculture: insights from a case study in the Mediterranean basin. Agric Syst , 147, 65–75, doi:10.1016/j.agsy.2016.05.013.
Doody, D.G., C.A. Augustenborg, P.J.A. Withers and S. Crosse, 2015: A systematic map protocol: what evidence exists to link agricultural practices with ecological impacts for Irish waterbodies?Environ. Evid. , 4 (1), 14, doi:10.1186/s13750-015-0040-9.
Dop, M.C., et al., 2020: Identification and frequency of consumption of wild edible plants over a year in central Tunisia: a mixed-methods approach. Public Health Nutr. , 23 (5), 782–794, doi:10.1017/S1368980019003409.
Dotchamou, F.T., G. Atindogbe, A.I. Sode and H.N. Fonton, 2016: Density and spatial distribution of Parkia biglobosa pattern in Benin under climate change. J. Agric. Environ. Int. Dev. , 110, doi:10.12895/jaeid.20161.447.
Douhard, M., et al., 2016: The influence of weather conditions during gestation on life histories in a wild Arctic ungulate. Proc. Biol. Sci. , 283 (1841), 20161760, doi:10.1098/rspb.2016.1760.
Dounias, E. and M. Ichikawa, 2017: Seasonal Bushmeat Hunger in the Congo Basin. EcoHealth, 14 (3), 575–590, doi:10.1007/s10393-017-1252-y.
Dounias, E., A. Selzner, M. Koizumi and P. Levang, 2007: From sago to rice, from forest to town: the consequences of sedentarization for the nutritional ecology of Punan former hunter-gatherers of Borneo. Food Nutr Bull, 28 (2 Suppl), S294–S302, doi:10.1177/15648265070282S208.
Dovie, D.B.K., 2017: A communication framework for climatic risk and enhanced green growth in the eastern coast of Ghana. Land Use Policy, 62, 326–336, doi:10.1016/j.landusepol.2017.01.008.
Dowd-Uribe, B., 2014: Engineering yields and inequality? How institutions and agro-ecology shape Bt cotton outcomes in Burkina Faso. Geoforum, 53, 161–171.
Dowd-Uribe, B. and M. A. Schnurr, 2016: Briefing: Burkina Faso’s reversal on genetically modified cotton and the implications for Africa. Afr Aff (Lond) , 115 (458), 161–172, doi:10.1093/afraf/adv063.
Downing, A. and A. Cuerrier, 2011: A synthesis of the impacts of climate change on the First Nations and Inuit of Canada. Indian. J. Tradit. Knowl. , 10 (1), 57–70.
Doyle, J.T., M.H. Redsteer and M.J. Eggers, 2013: Exploring effects of climate change on Northern Plains American Indian health. In: Climate Change and Indigenous Peoples in the United States: Impacts, Experiences and Actions[Maldonado, J.K., B. Colombi and R. Pandya(eds.)]. Springer, Cham, Switzerland, Heidelberg , Germany, New York, NY, USA, Dordrecht, the Netherlands, London, UK, pp. 135–147. ISBN 978-3319052656.
Doyle, L., L. Oliver and C. Whitworth, 2018: Design of a climate smart farming system in East Africa. In: GHTC 2018. IEEE Global Humanitarian Technology Conference, 18 - 21 October 2018. USA, Institute of Electrical and Electronics Engineers Xplore, Piscataway, NJ, USA, San Jose, CA, pp. 1–6. doi:10.1109/GHTC.2018.8601553.
Drag, D.W., et al., 2020: Soybean photosynthetic and biomass responses to carbon dioxide concentrations ranging from pre-industrial to the distant future. J. Exp. Bot. , 71 (12), 3690–3700.
Dressler, G., et al., 2019: Polarization in (post)nomadic resource use in Eastern Morocco: insights using a multi-agent simulation model. Reg Environ Change, 19 (2), 489–500, doi:10.1007/s10113-018-1412-9.
Drewry, J.L., et al., 2019: Assessment of digital technology adoption and access barriers among crop, dairy and livestock producers in Wisconsin. Comput. Electron. Agric. , 165, 104960, doi:10.1016/j.compag.2019.104960.
Du, B.Z., L. Zhen, H.M. Yan and R. de Groot, 2016: Effects of government grassland conservation policy on household livelihoods and dependence on local grasslands: evidence from Inner Mongolia, China. Sustainability, 8 (12), 1314, doi:10.3390/su8121314.
Du, Z., et al., 2021: Exploring microbial community structure and metabolic gene clusters during silage fermentation of paper mulberry, a high-protein woody plant. Anim. Feed. Sci. Technol. , 275, 114766–114766, doi:10.1016/j.anifeedsci.2020.114766.
Dua, V.K., et al., 2013: Impact of climate change on potato productivity in Punjab - a simulation study. Curr. Sci. , 105 (6), 787–794, doi:10.2307/24097516.
Dubbeling, M., 2015: Integrating Urban Agriculture and Forestry into Climate Change Action Plans: Lessons from Western Province, Sri Lanka and Rosario, Argentina. Resource Centers on Urban Agriculture & Food Security Foundation, https://cdkn.org/wp-content/uploads/2015/02/SriLanka_Argentina_BackgroundPaper_FINAL_WEB.pdf. Accessed 2021 . (18 pp).
Dubbeling, M., et al., 2017: Assessing and planning sustainable city region food systems: insights from two Latin American Cities. Sustainability, 9 (8), 1455, doi:10.3390/su9081455.
Dubey, S.K., et al., 2017: Farmers’ perceptions of climate change, impacts on freshwater aquaculture and adaptation strategies in climatic change hotspots: a case of the Indian Sundarban delta. Environ. Dev. , 21, 38–51, doi:10.1016/j.envdev.2016.12.002.
Dubois, M.J., et al., 2019: Integrating fish into irrigation infrastructure projects in Myanmar: rice-fish what if…?Mar. Freshw. Res. , 70 (9), 1229–1240, doi:10.1071/MF19182.
Dumas, S.E., L. Kassa, S.L. Young and A.J. Travis, 2018: Examining the association between livestock ownership typologies and child nutrition in the Luangwa Valley, Zambia. Plos One, 13 (2), e191339, doi:10.1371/journal.pone.0191339.
Dunlap, A. and J. Fairhead, 2014: The Militarisation and Marketisation of Nature: An Alternative Lens to ‘Climate-Conflict’. Geopolitics, 19 (4), 937–961, doi:10.1080/14650045.2014.964864.
Durland, E., G. Waldbusser and C. Langdon, 2019: Comparison of larval development in domesticated and naturalized stocks of the Pacific oyster Crassostrea gigas exposed to high pCO2 conditions. Mar. Ecol. Prog. Ser. , 621, 107–125, doi:10.3354/meps12983.
Dury, S., et al. (ed.), 2019: Food Systems at Risk: New Trends and Challenges. FAO-CIRAD-European Commission, Rome, Italy, Montpellier, France, and Brussels, Belgium, ISBN 978-2876147515. 130 pp.
Duveneck, M.J. and R.M. Scheller, 2015: Climate-suitable planting as a strategy for maintaining forest productivity and functional diversity. Ecol Appl, 25 (6), 1653–1668, doi:10.1890/14-0738.1.
Dyngeland, C., J.A. Oldekop and K.L. Evans, 2020: Assessing multidimensional sustainability: Lessons from Brazil’s social protection programs. Proc. Natl. Acad. Sci. , 117 (34), 20511–20519, doi:10.1073/pnas.1920998117.
Eastin, J., 2018: Climate change and gender equality in developing states. World Dev, 107, 289–305, doi:10.1016/j.worlddev.2018.02.021.
EAT-LANCET, 2019: Healthy Diets from Sustainable Food Systems. Summary Report of the EAT-Lancet Commission. Lancet, https://eatforum.org/content/uploads/2019/01/EAT-Lancet_Commission_Summary_Report.pdf. Accessed 2021 . (32 pp).
Ebert, A.W., 2017: Vegetable production, diseases, and climate change. In: World Agricultural Resources and Food Security: International Food Security[Schmitz, A., P.L. Kennedy and T.G. Schmitz(eds.)]. Emerald, Bingley, UK, pp. 103–124. ISBN 978-1787145160.
Ebhuoma, E. and D. Simatele, 2017: Defying the odds: climate variability, asset adaptation and food security nexus in the Delta State of Nigeria. Int. J. Disaster Risk Reduct. , 21, 231–242, doi:10.1016/j.ijdrr.2016.12.017.
Ebhuoma, E.E., et al., 2020: Subsistence farmers’ differential vulnerability to drought in Mpumalanga province, South Africa: Under the political ecology spotlight. Cogent Soc. Sci. , 6 (1), 1792155–1792155, doi:10.1080/23311886.2020.1792155.
Ebi, K.L. and I. Loladze, 2019: Elevated atmospheric CO2 concentrations and climate change will affect our food’s quality and quantity. Lancet Planet. Health, 3 (7), E283–E284, doi:10.1016/S2542-5196(19)30108-1.
Economic Commission for Latin America and the Caribbean and Central American Agricultural Council of the Central American Integration System, 2013: Impactos Potenciales del Cambio Climático sobre los Granos Básicos en Centroamérica. Economic Commission for Latin America and the Caribbean, Mexico City, Mexico, https://www.cepal.org/es/publicaciones/27171-impactos-potenciales-cambio-climatico-granos-basicos-centroamerica. Accessed 2021 . (137 pp).
Economic Commission for Latin America and the Caribbean and C. A. A. C. o. t. C. A. I. System, 2014: Impactos Potenciales del Cambio Climático sobre el Café en Centroamérica. Economic Commission for Latin America and the Caribbean, México City, Mexico, https://www.cepal.org/es/publicaciones/37456-impactos-potenciales-cambio-climatico-cafe-centroamerica. Accessed 2021 . (129 pp).
Edwards, E.J., D. Unwin, R. Kilmister and M. Treeby, 2017: Multi-seasonal effects of warming and elevated CO2 on the physiology, growth and production of mature, field grown, Shiraz grapevines. Oeno One, 51 (2), 127–132, doi:10.20870/oeno-one.2016.0.0.1586.
Egli, L., et al., 2018: Winners and losers of national and global efforts to reconcile agricultural intensification and biodiversity conservation. Glob Chang Biol, 24 (5), 2212–2228, doi:10.1111/gcb.14076.
Eigenbrod, C. and N. Gruda, 2015: Urban vegetable for food security in cities. A review. Agron. Sustain. Dev. , 35 (2), 483–498, doi:10.1007/s13593-014-0273-y.
Ekstrom, J.A., et al., 2015: Vulnerability and adaptation of US shellfisheries to ocean acidification. Nat. Clim. Chang. , 5 (3), 207–214, doi:10.1038/Nclimate2508.
El-Essawy, H., P. Nasr and H. Sewilam, 2019: Aquaponics: a sustainable alternative to conventional agriculture in Egypt - a pilot scale investigation. Environ. Sci. Pollut. Res. Int. , 26 (16), 15872–15883, doi:10.1007/s11356-019-04970-0.
El Yaacoubi, A., et al., 2014: Global warming impact on floral phenology of fruit trees species in Mediterranean region. Sci. Hortic. , 180, 243–253.
Elaswad, A., et al., 2018: Microinjection of CRISPR/Cas9 protein into channel catfish, Ictalurus punctatus, embryos for gene editing. J. Vis. Exp. , (131), e56275, doi:10.3791/56275.
Elkin, C., A. Giuggiola, A. Rigling and H. Bugmann, 2015: Short- and long-term efficacy of forest thinning to mitigate drought impacts in mountain forests in the European Alps. Ecol Appl, 25 (4), 1083–1098, doi:10.1890/14-0690.1.
Elkin, C., et al., 2013: A 2 degrees C warmer world is not safe for ecosystem services in the European Alps. Glob Chang Biol, 19 (6), 1827–1840, doi:10.1111/gcb.12156.
Ellis, N.R. and P. Tschakert, 2019: Triple-wins as pathways to transformation? A critical review. Geoforum, 103, 167–170, doi:10.1016/j.geoforum.2018.12.006.
Elmendorf, S.C., et al., 2012: Global assessment of experimental climate warming on tundra vegetation: heterogeneity over space and time. Ecol Lett , 15 (2), 164–175, doi:10.1111/j.1461-0248.2011.01716.x.
Elmhirst, R., M. Siscawati, B.S. Basnett and D. Ekowati, 2017: Gender and generation in engagements with oil palm in East Kalimantan, Indonesia: insights from feminist political ecology. J Peasant Stud, 44 (6), 1137–1159, doi:10.1080/03066150.2017.1337002.
Emmerson, M., et al., 2016: How agricultural intensification affects biodiversity and ecosystem services. In: Large-Scale Ecology: Model Systems to Global Perspectives[Dumbrell, A.J., R.L. Kordas and G. Woodward(eds.)]. Academic, London, UK, Oxford, UK, San Diego, CA, USA, and Cambridge, MA, USA, pp. 43–97. ISBN 978-0081009352.
Engler, C., 2020: Transboundary fisheries, climate change, and the ecosystem approach: taking stock of the international law and policy seascape. Ecol. Soc. , 25 (4), doi:10.5751/ES-11988-250443.
Englund, O., et al., 2020: Beneficial land use change: Strategic expansion of new biomass plantations can reduce environmental impacts from EU agriculture. Glob. Environ. Chang. , 60, 101990–101990, doi:10.1016/j.gloenvcha.2019.101990.
Ensor, J.E., et al., 2016: Can community-based adaptation increase resilience?Clim. Dev. , 10 (2), 134–151, doi:10.1080/17565529.2016.1223595.
Erauskin-Extramiana, M., et al., 2019: Large-scale distribution of tuna species in a warming ocean. Glob Chang Biol, 25 (6), 2043–2060, doi:10.1111/gcb.14630.
Eriksen, C. and D.L. Hankins, 2014: The Retention, Revival, and Subjugation of Indigenous Fire Knowledge through Agency Fire Fighting in Eastern Australia and California. Soc. Nat. Resour. , 27 (12), 1288–1303, doi:10.1080/08941920.2014.918226.
Eriksen, S., et al., 2021: Adaptation interventions and their effect on vulnerability in developing countries: Help, hindrance or irrelevance?World Dev, 141, 105383–105383, doi:10.1016/j.worlddev.2020.105383.
Eriksen, S.H., L.K. Cramer, I. Vetrhus and P. Thornton, 2019: Can climate interventions open up space for transformation? Examining the case of climate-smart agriculture (CSA) in Uganda. Front. Sustain. Food Syst. , 3, doi:10.3389/fsufs.2019.00111.
Espeland, E.K., L. Schreeg and L.M. Porensky, 2020: Managing risks related to climate variability in rangeland-based livestock production: what producer driven strategies are shared and prevalent across diverse dryland geographies?J Environ Manage, 255, 109889, doi:10.1016/j.jenvman.2019.109889.
Estevez, P., et al., 2019: Emerging marine biotoxins in seafood from European coasts: incidence and analytical challenges. Foods, 8 (5), 149, doi:10.3390/foods8050149.
European Commission Directorate-General for Research Innovation, 2018: Sustainable & circular bioeconomy, the European way Outcome report [Burrows, D. (ed.)]. outcome report, Publications Office, Brussels
European Food Safety Authority et al., 2020: Climate change as a driver of emerging risks for food and feed safety, plant, animal health and nutritional quality. EFSA Support. Publ. , 17 (6), doi:10.2903/sp.efsa.2020.EN-1881. EN-1881.
European Food Safety Authority Panel on Plant Protection Products and their Residues et al., 2017: Scientific Opinion of the PPR Panel on the follow-up of the findings of the External Scientific Report ‘Literature review of epidemiological studies linking exposure to pesticides and health effects’. EFS2, 15 (10), e5007, doi:10.2903/j.efsa.2017.5007.
Evaristus, I., 2014: Global change and sustainable mountain tourism: the case of Mount Kenya. In: Impact of Global Changes on Mountains: Responses and Adaptation[Grover, V.I., A. Bordorf, J. Breuste, P. Chandra Tiwari and F. Witkowski Frangetto(eds.)]. CRC Press, Boca Raton, FL, USA, pp. 197–217. ISBN 978-0429172915.
Eyring, V., N.P. Gillett, K.M. Achuta Rao, R. Barimalala, M. Barreiro Parrillo, N. Bellouin, C. Cassou, P.J. Durack, Y. Kosaka, S. McGregor, S. Min, O. Morgenstern, and Y. Sun, 2021: Human Influence on the Climate System. In: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change[Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press.
Eyssartier, C., A.H. Ladio and M. Lozada, 2011: Horticultural and gathering practices complement each other: a case study in a rural population of northwestern Patagonia. Ecol. Food. Nutr. , 50 (5), 429–451, doi:10.1080/03670244.2011.604587.
Fa, J.E., et al., 2015: Disentangling the relative effects of bushmeat availability on human nutrition in central Africa. Sci Rep, 5, 8168, doi:10.1038/srep08168.
Fabbri, S., S.I. Olsen and M. Owsianiak, 2018: Improving environmental performance of post-harvest supply chains of fruits and vegetables in Europe: potential contribution from ultrasonic humidification. J Clean Prod, 182, 16–26, doi:10.1016/j.jclepro.2018.01.157.
Fader, M., et al., 2013: Spatial decoupling of agricultural production and consumption: quantifying dependences of countries on food imports due to domestic land and water constraints. Environ. Res. Lett. , 8 (1), 14046, doi:10.1088/1748-9326/8/1/014046.
Fagundes, C., et al., 2020: Ecological costs of discrimination: racism, red cedar and resilience in farm bill conservation policy in Oklahoma. Renew. Agric. Food Syst. , 35 (4), 420–434, doi:10.1017/S1742170519000322.
Fairbairn, M., 2014: ‘Like gold with yield’: evolving intersections between farmland and finance. J Peasant Stud, 41 (5), 777–795, doi:10.1080/03066150.2013.873977.
Fairbairn, M., 2020: Fields of Gold: Financing the Global Land Rush. Cornell University Press.
Fajinmi, O.O., O.O. Olarewaju and J. Van Staden, 2017: Traditional use of medicinal and aromatic plants in Africa. In: Medicinal and Aromatic Plants of the World - Africa[Neffati, M., H. Najjaa and Á. Máthé(eds.)]. Springer Science + Business, Dordrecht , the Netherlands, pp. 61–76.
Falconer, L., et al., 2020a: The importance of calibrating climate change projections to local conditions at aquaculture sites. Aquaculture, 514, 734487, doi:10.1016/j.aquaculture.2019.734487.
Falconer, L., et al., 2020b: Use of geographic information systems for aquaculture and recommendations for development of spatial tools. Rev. Aquac. , 12 (2), 664–677, doi:10.1111/raq.12345.
Falkenberg, L.J., C.A. Styan and J.N. Havenhand, 2019: Sperm motility of oysters from distinct populations differs in response to ocean acidification and freshening. Sci Rep, 9 (1), 7970, doi:10.1038/s41598-019-44321-0.
FAO, 2013: The Human Right to Adequate Food in the Global Strategic Framework for Food Security and Nutrition: A Global Consensus. Rome, http://www.fao.org/3/i3546e/i3546e.pdf. (26-26 pp).
FAO, 2014: State of the World’s Forests: Enhancing the Socioeconomic Benefits from Forests. The State of the World, Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978925108269. 119 pp.
FAO, 2015: Voluntary Guidelines for Securing Sustainable Small-Scale Fisheries in the Context of Food Security and Poverty Eradication. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251087046. 18 pp.
FAO, 2016: Fisheries and Aquaculture Country Profiles: Ghana (2016). Country Profile Fact Sheets. FAO, Rome.
FAO, 2017: Adaptation Strategies of the Aquaculture Sector to the Impacts of Climate Change. FAO Fisheries and Aquaculture Circular, Vol. 1142. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251096864. 28 pp.
FAO, 2018a: The Future of Food and Agriculture – Alternative Pathways to 2050. Future of Food and Agriculture, Vol. 2. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251301586. 202 pp.
FAO, 2018b: Impacts of climate change on fisheries and aquaculture: synthesis of current knowledge, adaptation and mitigation options. Food and Agriculture Organization of the United Nations, Rome.
FAO, 2018c: The State of World Fisheries and Aquaculture 2018 - Meeting the Sustainable Development Goals. The State of the World, Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9551305621. 210 pp.
FAO, 2019a: Climate-smart agriculture and the Sustainable Development Goals:Mapping interlinkages, synergies and trade-offs and guidelines for integrated implementation. Rome, http://www.fao.org/climate-smart-agriculture/resources/publications/en/. (1-144 pp).
FAO, 2019b: FAO Framework for the Urban Food Agenda. Leveraging Sub-National and Local Government Action to Ensure Sustainable Food Systems and Improved Nutrition. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251312742. 41 pp.
FAO, 2019c: Report of the Special Session on Advancing Integrated Agriculture Aquaculture through Agroecology. Montpellier, France, 25.08.2018. FAO Fisheries and Aquaculture Report, Vol. 1286. FAO, Rome. https://agritrop.cirad.fr/594534/1/ID594534.pdf.
FAO, 2019d: The State of the World’s Aquatic Genetic Resources for Food and Agriculture. FAO Commission on Genetic Resources for Food and Agriculture assessments, FAO, Rome. http://www.fao.org/3/ca5256en/CA5256EN.pdf.
FAO, 2019e: The State of the World’s Biodiversity for Food and Agriculture. In: FAO Commission on Genetic Resources for Food and Agriculture Assessment [Bélanger, J. and D. Pilling(eds.)]. FAO, Rome, Italy. ISBN 978-9251312704 572 pp).
FAO, 2019 f: World Food and Agriculture - Statistical Pocketbook 2019. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251318492. 248 pp.
FAO, 2020a: Climate Change: Unpacking the Burden on Food Safety. Food Safety and Quality Series, Food and Agriculture Organization of the United States, Rome, Italy, ISBN 978-9251322932. 154 pp.
FAO, 2020b: Farmers and agribusinesses at risk under COVID-19;What role for blended finance funds?FAO, Rome, Italy. 11 pp.
FAO, 2020c: Fisheries and Aquaculture Statistics: Global aquaculture and fisheries production 1950-2018 (FishstatJ). [(online)., F. F. a. A. D. (ed.). www.fao.org/fishery/statistics/software/fishstatj/en.
FAO, 2020d: The State of World Fisheries and Aquaculture 2020. Sustainability in Action. The State of the World, Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251326923. 206 pp.
FAO, 2021: The Impact of Disasters and Crises on Agriculture and Food Security. FAO, Rome, Italy, ISBN 978-9251340714. 143 pp.
FAO, et al., 2018: The State of Food Security and Nutrition in the World 2018. Building Climate Resilience for Food Security and Nutrition. The State of the World, Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251305713. 181 pp.
FAO, et al., 2020: The State of Food Security and Nutrition in the World 2020. FAO, IFAD, UNICEF, WFP and WHO, Rome, 320–320. ISBN 978-9251329016.
FAO and ITPS, 2015: Status of the World’s Soil Resources - Main Report . Food and Agriculture Organization of the United Nations and Intergovernmental Technical Panel on Soils, Rome, Italy, ISBN 978-9251090046.
FAO and WHO, 2016: Evaluation of certain contaminants in food. Eighty-third report of the Joint FAO/WHO Expert Committee on Food Additives. WHO Technical Report, World Health Organization and the Food and Agriculture Organization of the United Nations, Malta, ISBN 978-9241210027. 166 pp.
Farady, S.E. and T.E. Bigford, 2019: Fisheries and climate change: legal and management implications. Fisheries, 44 (6), 270–275, doi:10.1002/fsh.10263.
Farcy, P., et al., 2019: Toward a European coastal observing network to provide better answers to science and to societal challenges; the JERICO research infrastructure. Front. Mar. Sci. , 6, 529, doi:10.3389/fmars.2019.00529.
Farhat, Y.A., et al., 2021: Altered arsenic availability, uptake, and allocation in rice under elevated temperature. Sci. Total Environ. , 763, 143049–143049, doi:10.1016/j.scitotenv.2020.143049.
Farmery, A.K., et al., 2021: Food for all: designing sustainable and secure future seafood systems. Rev. Fish Biol. Fish. , doi:10.1007/s11160-021-09663-x.
Farquhar, S.D., et al., 2018: Socio-economic impacts of the women in aquaculture (WiA) project in Nepal. Kasetsart J. Soc. Sci. , 40 (2), 289–295, doi:10.1016/j.kjss.2017.12.014.
Farrell, A.D., K. Rhiney, A. Eitzinger and P. Umaharan, 2018: Climate adaptation in a minor crop species: is the cocoa breeding network prepared for climate change?Agroecol. Sustain. Food Syst. , 42 (7), 812–833, doi:10.1080/21683565.2018.1448924.
Faruque, G., et al., 2017: The evolution of aquatic agricultural systems in Southwest Bangladesh in response to salinity and other drivers of change. Int. J. Agric. Sustain. , 15 (2), 185–207, doi:10.1080/14735903.2016.1193424.
Favero, A., R. Mendelsohn and B. Sohngen, 2018: Can the global forest sector survive 11° C warming?Agric. Econ. Res. Rev. , 47, 388–413, doi:10.1017/age.2018.15.
Favero, A., R. Mendelsohn, B. Sohngen and B.D. Stocker, 2021: Assessing the long-term interactions of climate change and timber markets on forest land and carbon storage. Environ. Res. Lett. , 16 (1), 14051, doi:10.1088/1748-9326/abd589.
Favretto, N., et al., 2020: Delivering Climate-Development Co-Benefits through Multi-Stakeholder Forestry Projects in Madagascar: Opportunities and Challenges. Land, 9 (5), 157–157, doi:10.3390/LAND9050157.
Fawcett, D., et al., 2018: Inuit adaptability to changing environmental conditions over an 11-year period in Ulukhaktok, Northwest Territories. Polar. Rec. , 54 (2), 119–132, doi:10.1017/S003224741800027x.
Faye, P. and J. Ribot, 2017: Causes for adaptation: Access to forests, markets and representation in Eastern Senegal. Sustain. (switzerland) , 9 (2), 1–20, doi:10.3390/su9020311.
Fedele, G., et al., 2019: Transformative adaptation to climate change for sustainable social-ecological systems. Environ Sci Policy, 101, 116–125, doi:10.1016/j.envsci.2019.07.001.
Fedele, G., B. Locatelli, H. Djoudi and M.J. Colloff, 2018: Reducing risks by transforming landscapes: cross-scale effects of land-use changes on ecosystem services. PLoS ONE, 13 (4), e195895, doi:10.1371/journal.pone.0195895.
Fei, S., et al., 2017: Divergence of species responses to climate change. Sci. Adv. , 3 (5), e1603055, doi:10.1126/sciadv.1603055.
Feliciano, D., A. Ledo, J. Hillier and D.R. Nayak, 2018: Which agroforestry options give the greatest soil and above ground carbon benefits in different world regions?Agric. Ecosyst. Environ. , 254, 117–129, doi:10.1016/j.agee.2017.11.032.
Felton, A., et al., 2016: How climate change adaptation and mitigation strategies can threaten or enhance the biodiversity of production forests: Insights from Sweden. Biol. Conserv. , 194, 11–20, doi:10.1016/j.biocon.2015.11.030.
Feng, W., et al., 2017: Enhanced decomposition of stable soil organic carbon and microbial catabolic potentials by long-term field warming. Glob Chang Biol, 23 (11), 4765–4776, doi:10.1111/gcb.13755.
Fenton, A., J. Paavola and A. Tallontire, 2017: Autonomous adaptation to riverine flooding in Satkhira District, Bangladesh: implications for adaptation planning. Reg Environ Change, 17 (8), 2387–2396, doi:10.1007/s10113-017-1159-8.
Fernandes, J.A., et al., 2017: Estimating the ecological, economic and social impacts of ocean acidification and warming on UK fisheries. Fish. Fish. , 18 (3), 389–411, doi:10.1111/faf.12183.
Fernández-Giménez, M. E., B. Batkhishig, B. Batbuyan and T. Ulambayar, 2015: Lessons from the Dzud: community-based rangeland management increases the adaptive capacity of Mongolian herders to winter disasters. World Dev, 68 (1), 48–65, doi:10.1016/j.worlddev.2014.11.015.
Fernando, N., et al., 2019: Annual ryegrass (Lolium rigidum Gaud) competition altered wheat grain quality: a study under elevated atmospheric CO2 levels and drought conditions. Food Chem, 276, 285–290, doi:10.1016/j.foodchem.2018.09.145.
Fetzel, T., et al., 2017: Quantification of uncertainties in global grazing systems assessment. Glob. Biogeochem. Cycles, 31 (7), 1089–1102, doi:10.1002/2016gb005601.
Fidelman, P., T.V. Tuyen, K. Nong and M. Nursey-Bray, 2017: The institutions-adaptive capacity nexus: Insights from coastal resources co-management in Cambodia and Vietnam. Environ Sci Policy, 76, 103–112, doi:10.1016/j.envsci.2017.06.018.
Figueroa-Helland, L., C. Thomas and A.P. Aguilera, 2018: Decolonizing food systems: food sovereignty, indigenous revitalization, and agroecology as counter-hegemonic movements. Perspect. Glob. Dev. Technol. , 17 (1), 173–201, doi:10.1163/15691497-12341473.
Fikir, D., W. Tadesse and A. Gure, 2016: Economic contribution to local livelihoods and households dependency on dry land forest products in Hammer District, Southeastern Ethiopia. Int. J. For. Res. , 2016, 5474680, doi:10.1155/2016/547468.
Filipe, J.F., et al., 2020: Floods, hurricanes, and other catastrophes: a challenge for the immune system of livestock and other animals. Front. Vet. Sci. , 7, 16, doi:10.3389/fvets.2020.00016.
Fink, S., 2017: Loss of habitat: impacts on pinnipeds and their welfare. In: Marine Mammal Welfare: Human Induced Change in the Marine Environment and its Impacts on Marine Mammal Welfare[Butterworth, A.(ed.)]. Springer International Publishing, Cham, Switzerland, pp. 241–252. ISBN 978-3319469935.
Finkbeiner, E.M., et al., 2018: Exploring trade-offs in climate change response in the context of Pacific Island fisheries. Mar. Policy. , 88, 359–364, doi:10.1016/j.marpol.2017.09.032.
Fiore, A.M., et al., 2012: Global air quality and climate. Chem. Soc. Rev. , 41 (19), 6663–6683, doi:10.1039/c2cs35095e.
Fischer, J., et al., 2007: Mind the sustainability gap. Trends Ecol. Evol. , 22 (12), 621–624, doi:10.1016/j.tree.2007.08.016.
Fischer, K., 2016: Why new crop technology is not scale-neutral - a critique of the expectations for a crop-based African Green Revolution. Res Policy, 45 (6), 1185–1194, doi:10.1016/j.respol.2016.03.007.
Fischer, K., J. Van den Berg and C. Mutengwa, 2015: Is Bt maize effective in improving South African smallholder agriculture?S. Afr. J. Sci. , 111 (1/2), a92, doi:10.17159/sajs.2015/a0092.
Fischler, C., 1988: Food, self and identity. Soc. Sci. Inf. , 27 (2), 275–292, doi:10.1177/053901888027002005.
Fisher, B., et al., 2017: Integrating fisheries and agricultural programs for food security. Agric. Food Secur. , 6 (1), 1, doi:10.1186/s40066-016-0078-0.
Fitzer, S.C., et al., 2019: Selectively bred oysters can alter their biomineralization pathways, promoting resilience to environmental acidification. Glob Chang Biol, 25 (12), 4105–4115, doi:10.1111/gcb.14818.
Fitzgerald, G.J., et al., 2016: Elevated atmospheric [CO2 ] can dramatically increase wheat yields in semi-arid environments and buffer against heat waves. Glob Chang Biol, 22 (6), 2269–2284, doi:10.1111/gcb.13263.
Fleisher, D.H., et al., 2017: A potato model intercomparison across varying climates and productivity levels. Glob. Chang. Biol. , 23 (3), 1258–1281, doi:10.1111/gcb.13411.
Fleming, Z.L., et al., 2018: Tropospheric ozone assessment report: present-day ozone distribution and trends relevant to human health. Elem. Sci. Anthropocene, 6, 12, doi:10.1525/elementa.273.
Flörke, M., et al., 2019: Analysing trade-offs between SDGs related to water quality using salinity as a marker. Curr. Opin. Environ. Sustain. , 36, 96–104, doi:10.1016/j.cosust.2018.10.005.
Flörke, M., C. Schneider and R.I. McDonald, 2018: Water competition between cities and agriculture driven by climate change and urban growth. Nat. Sustain. , 1, 51–58, doi:10.1016/j.cosust.2018.10.005.
Flouris, A.D., et al., 2018: Workers’ health and productivity under occupational heat strain: a systematic review and meta-analysis. Lancet Planet Health, 2 (12), e521–e531, doi:10.1016/S2542-5196(18)30237-7.
Flynn, J. and J. Sumberg, 2021: Are Africa’s rural youth abandoning agriculture?[Sumberg, J. (ed.)]. Youth and the rural economy in Africa: hard work and hazard CABI, CABI, 43--57 pp. Available at: http://www.cabi.org/cabebooks/ebook/20210138606 (accessed 31/8//2021)
Fodor, N., et al., 2018: Spatially explicit estimation of heat stress-related impacts of climate change on the milk production of dairy cows in the United Kingdom. PLoS ONE, 13 (5), e197076, doi:10.1371/journal.pone.0197076.
Folberth, C., et al., 2019: Parameterization-induced uncertainties and impacts of crop management harmonization in a global gridded crop model ensemble. PLoS ONE, 14 (9), e221862.
Fonjong, L., I. Sama-Lang, L. Fombe and C. Abonge, 2016: Land governance and women’s rights in large-scale land acquisitions in Cameroon. Dev. Pract. , 26 (4), 420–430, doi:10.1080/09614524.2016.1162285.
Food and Land Use Coalition, 2019: Growing Better: Ten Critical Transitions to Transform Food and Land Use. The Global Consultation Report of the Food and Land Use Coalition. Food and Land Use Coalition, 237 pp. Available at: https://www.foodandlandusecoalition.org/wp-content/uploads/2019/09/FOLU-GrowingBetter-GlobalReport.pdf (accessed July 26, 2021).
Food Security Information Network, 2020: 2020 Global report on food crises: Joint analysis for better decisions. Food and Agriculture Organization (FAO); World Food Programme (WFP); and International Food Policy Research Institute (IFPRI), Rome, Italy and Washington DC, https://www.fsinplatform.org/global-report-food-crises-2020. (233 pp).
Forbes, B.C., et al., 2016: Sea ice, rain-on-snow and tundra reindeer nomadism in Arctic Russia. Biol. Lett. , 12 (11), 20160466, doi:10.1098/rsbl.2016.0466.
Forbes, B.C., et al., 2020: Changes in mountain birch forests and reindeer management: comparing different knowledge systems in Sápmi, northern Fennoscandia. Polar. Rec. , 55 (6), 507–521, doi:10.1017/s0032247419000834.
Forbis, Jr., R. and K. Hayhoe, 2018: Does Arctic governance hold the key to achieving climate policy targets?Environ. Res. Lett. , 13, 1–4, doi:10.1088/1748-9326/aaa359.
Ford, J.D., 2009: Vulnerability of Inuit food systems to food insecurity as a consequence of climate change: a case study from Igloolik, Nunavut. Reg Environ Change, 9 (2), 83–100, doi:10.1007/s10113-008-0060-x.
Ford, J.D., et al., 2015: The status of climate change adaptation in Africa and Asia. Reg Environ Change, 15 (5), 801–814, doi:10.1007/s10113-014-0648-2.
Ford, J.D., et al., 2019: Changing access to ice, land and water in Arctic communities. Nat. Clim. Chang. , 9 (4), 335–339, doi:10.1038/s41558-019-0435-7.
Ford, J.D., N. Couture, T. Bell and D.G. Clark, 2018: Climate change and Canada’s north coast: research trends, progress, and future directions. Environ. Rev. , 26 (1), 82–92, doi:10.1139/er-2017-0027.
Forest Trends, 2014: Consumer goods and deforestation: An analysis of the extent and nature of illegality in forest conversation for agriculture and timber plantations. Forest Trade and Finance Series, Washington, D.C.
Fort, J., et al., 2016: Does temporal variation of mercury levels in Arctic seabirds reflect changes in global environmental contamination, or a modification of Arctic marine food web functioning?Environ. Pollut. , 211, 382–388, doi:10.1016/j.envpol.2015.12.061.
Foster, J., et al., 2021: An advanced empirical model for quantifying the impact of heat and climate change on human physical work capacity. Int. J. Biometeorol. , 65 (7), 1215–1229, doi:10.1007/s00484-021-02105-0.
Foster, S., et al., 2018: Impact of irrigated agriculture on groundwater-recharge salinity: a major sustainability concern in semi-arid regions. Hydrogeol J, 26 (8), 2781–2791, doi:10.1007/s10040-018-1830-2.
Fox-Kemper, B., H.T. Hewitt, C. Xiao, G. Aðalgeirsdóttir, S.S. Drijfhout, T.L. Edwards, N.R. Golledge, M. Hemer, R.E. Kopp, G. Krinner, A. Mix, D. Notz, S. Nowicki, I.S. Nurhati, L. Ruiz, J.-B. Sallée, A.B.A. Slangen, and Y. Yu, 2021: Ocean, Cryosphere and Sea Level Change. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change[Masson-Delmotte, V., P. Zhai, A. Pirani, S.L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M.I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J.B.R. Matthews, T.K. Maycock, T. Waterfield, O. Yelekçi, R. Yu, and B. Zhou (eds.)]. Cambridge University Press.
Fox, J., J.C. Castella and A.D. Ziegler, 2014: Swidden, rubber and carbon: can REDD+ work for people and the environment in Montane Mainland Southeast Asia?Glob. Environ. Chang. , 29, 318–326, doi:10.1016/j.gloenvcha.2013.05.011.
Foyer, C.H., et al., 2016: Neglecting legumes has compromised human health and sustainable food production. Nat. Plants, 2, 16112, doi:10.1038/nplants.2016.112.
Fraga, H., I. Garcia de Cortazar Atauri, A.C. Malheiro and J.A. Santos, 2016a: Modelling climate change impacts on viticultural yield, phenology and stress conditions in Europe. Glob Chang Biol, 22 (11), 3774–3788, doi:10.1111/gcb.13382.
Fraga, H., N. Guimaraes and J.A. Santos, 2019a: Future changes in rice bioclimatic growing conditions in Portugal. Agronomy, 9 (11), 674, doi:10.3390/agronomy9110674.
Fraga, H., J.G. Pinto and J.A. Santos, 2019b: Climate change projections for chilling and heat forcing conditions in European vineyards and olive orchards: a multi-model assessment. Clim Change, 152 (1), 179–193, doi:10.1007/s10584-018-2337-5.
Fraga, H., et al., 2016b: Climatic suitability of Portuguese grapevine varieties and climate change adaptation. Int. J. Clim. , 36 (1), 1–12, doi:10.1002/joc.4325.
Fragaszy, S. and A. Closas, 2016: Cultivating the desert: irrigation expansion and groundwater abstraction in Northern State. Sudan. water Altern. , 9 (1), 139–161.
Franck, C., S.M. Grandi and M.J. Eisenberg, 2013: Agricultural subsidies and the American obesity epidemic. Am. J. Prev. Med. , 45 (3), 327–333, doi:10.1016/j.amepre.2013.04.010.
Franco, J., C.M.Y. Park and R. Herre, 2017: Just standards: international regulatory instruments and social justice in complex resource conflicts. Can. J. Dev. Stud. / Revue Can. D’études Du Développement , 0 (0), 1–19, doi:10.1080/02255189.2017.1298520.
Franco, J.C. and S.M. Borras, 2019: Grey areas in green grabbing: subtle and indirect interconnections between climate change politics and land grabs and their implications for research. Land Use Policy, 84 (October 2018), 192–199, doi:10.1016/j.landusepol.2019.03.013.
Franco, J.C., S. Monsalve and S.M. Borras, 2015: Democratic land control and human rights. Curr. Opin. Environ. Sustain. , 15, 66–71, doi:10.1016/j.cosust.2015.08.010.
Freduah, G., P. Fidelman and T.F. Smith, 2019: Adaptive capacity of small-scale coastal fishers to climate and non-climate stressors in the Western region of Ghana. Geogr J, 185 (1), 96–110, doi:10.1111/geoj.12282.
Free, C.M., et al., 2020: Realistic fisheries management reforms could mitigate the impacts of climate change in most countries. PLoS ONE, 15 (3), e224347, doi:10.1371/journal.pone.0224347.
Free, C.M., et al., 2019: Impacts of historical warming on marine fisheries production. Science, 363 (6430), 979–983, doi:10.1126/science.aau1758.
Freed, S., et al., 2020: Maintaining Diversity of Integrated Rice and Fish Production Confers Adaptability of Food Systems to Global Change. Front. Sustain. Food Syst. , 4, doi:10.3389/fsufs.2020.576179.
Freitas, C., et al., 2008: Ringed seal post-moulting movement tactics and habitat selection. Oecologia, 155 (1), 193–204, doi:10.1007/s00442-007-0894-9.
Friant, S., et al., 2020: Eating bushmeat improves food security in a biodiversity and infectious disease “hotspot”. EcoHealth, 17 (1), 125–138, doi:10.1007/s10393-020-01473-0.
Friedrichs, S., et al., 2019: Meeting report of the OECD conference on “Genome editing: applications in agriculture-implications for health, environment and regulation”. Transgenic Res. , 28 (3-4), 419–463, doi:10.1007/s11248-019-00154-1.
Frieler, K., et al., 2017: Understanding the weather signal in national crop-yield variability. Earths Future, 5 (6), 605–616, doi:10.1002/2016EF000525.
Froehlich, H.E., R.R. Gentry and B.S. Halpern, 2018a: Global change in marine aquaculture production potential under climate change. Nat Ecol Evol, 2 (11), 1745–1750, doi:10.1038/s41559-018-0669-1.
Froehlich, H.E., et al., 2018b: Comparative terrestrial feed and land use of an aquaculture-dominant world. Proc. Natl. Acad. Sci. U. S. A. , 115 (20), 5295–5300, doi:10.1073/pnas.1801692115.
Froese, R. and J. Schilling, 2019: The nexus of climate change, land use, and conflicts. Curr Clim Change Rep, 5 (1), 24–35, doi:10.1007/s40641-019-00122-1.
Frolicher, T.L. and C. Laufkotter, 2018: Emerging risks from marine heat waves. Nat Commun, 9 (1), 650, doi:10.1038/s41467-018-03163-6.
Fry, J.P., et al., 2016: Environmental health impacts of feeding crops to farmed fish. Environ Int , 91, 201–214, doi:10.1016/j.envint.2016.02.022.
Fry, J.P., et al., 2018a: Corrigendum: Feed conversion efficiency in aquaculture: do we measure it correctly? (2018 Environ. Res. Lett. 13 024017). Environ. Res. Lett. , 13 (7), 79502, doi:10.1088/1748-9326/aad007.
Fry, J.P., et al., 2018b: Feed conversion efficiency in aquaculture: do we measure it correctly?Environ. Res. Lett. , 13 (2), 24017, doi:10.1088/1748-9326/aaa273.
FSIN, 2017: Global Report on Food Crises 2017. FSIN, Rome, Italy, 145-145 pp. Available at: https://documents.wfp.org/stellent/groups/public/documents/ena/wfp291271.pdf?_ga=2.269203057.1077894310.1641273785-1517166772.1641273785 (accessed August 31, 2021)
FSIN, 2021: 2021 Global Report on Food Crises: Joint analysis for better decisions. DC. Food and Agriculture Organization (FAO); World Food Programme (WFP); and International Food Policy Research Institute (IFPRI), Rome, Italy; and Washington, DO., https://www.fsinplatform.org/global-report-food-crises-2021.(last accessed January 3, 2021).
Fuchs, R., C. Brown and M. Rounsevell, 2020: Europe’s Green Deal offshores environmental damage to other nations. Nature, 586 (7831), 671–673, doi:10.1038/d41586-020-02991-1.
Fuhrer, J., P. Smith and A. Gobiet, 2014: Implications of climate change scenarios for agriculture in alpine regions--a case study in the Swiss Rhone catchment. Sci. Total Environ. , 493, 1232–1241, doi:10.1016/j.scitotenv.2013.06.038.
Fujimori, S., et al., 2018: Macroeconomic impacts of climate change driven by changes in crop yields. Sustainability, 10, 3673, doi:10.3390/su10103673.
Fujisaki, K., et al., 2018: Soil carbon stock changes in tropical croplands are mainly driven by carbon inputs: A synthesis. Agric. Ecosyst. Environ. , 259, 147–158, doi:10.1016/j.agee.2017.12.008.
Funes, I., et al., 2020: Soil water availability as a tool for adapting viticulture to climate change. Quaderns Agrar. , 49, 35–52, doi:10.2436/20.1503.01.116.
Funge-Smith, S. and A. Bennett, 2019: A fresh look at inland fisheries and their role in food security and livelihoods. Fish. Fish. , 20 (6), 1176–1195.
Funk, C., et al., 2018: Anthropogenic enhancement of moderate-to-strong el nino events likely contributed to drought and poor harvests in Southern Africa during 2016. Bull. Am. Meterol. Soc. , 99 (1), 91–S96, doi:10.1175/Bams-D-17-0112.1.
Furman, C., et al., 2014: Social justice in climate services: engaging African American farmers in the American South. Clim. Risk Manag. , 2, 11–25, doi:10.1016/j.crm.2014.02.002.
Furze, J.R., et al., 2017: Resistance and resilience of root fungal communities to water limitation in a temperate agroecosystem. Ecol Evol, 7 (10), 3443–3454, doi:10.1002/ece3.2900.
Fuss, S., et al., 2015: Global food security & adaptation under crop yield volatility. Technol Forecast Soc Change, 98, 223–233, doi:10.1016/j.techfore.2015.03.019.
Gabay, M. and M. Alam, 2017: Community forestry and its mitigation potential in the Anthropocene: The importance of land tenure governance and the threat of privatization. Policy Econ, 79, 26–35, doi:10.1016/j.forpol.2017.01.011.
Gabriel, A.G. and T.L.S. Mangahas, 2017: Indigenous people’s contribution to the mitigation of climate variation, their perception, and organizing strategy for sustainable community based forest resources management in Caraballo mountain, Philippines. Open J. Ecol. , 7 (2), 85–100, doi:10.4236/oje.2017.72007.
Gaines, S.D., et al., 2018: Improved fisheries management could offset many negative effects of climate change. Sci. Adv. , 4 (8), eaao1378, doi:10.1126/sciadv.aao1378.
Gaira, K.S., R.S. Rawal, B. Rawat and I.D. Bhatt, 2014: Impact of climate change on the flowering of Rhododendron arboreum in central Himalaya, India. Curr. Sci. , 106 (12), 1735–1738.
Gairola, S., N.M. Shariff, A. Bhatt and C.P. Kala, 2010: Influence of climate change on production of secondary chemicals in high altitude medicinal plants: issues needs immediate attention. J. Med. Plants Res. , 4 (18), 1825–1829, doi:10.5897/JMPR10.354.
Gajjar, S.P., C. Singh and T. Deshpande, 2019: Tracing back to move ahead: a review of development pathways that constrain adaptation futures. Clim. Dev. , 11 (3), 223–237, doi:10.1080/17565529.2018.1442793.
Galappaththi, E.K., J.D. Ford and E.M. Bennett, 2020a: Climate change and adaptation to social-ecological change: the case of indigenous people and culture-based fisheries in Sri Lanka. Clim Change, 162 (2), 279–300, doi:10.1007/s10584-020-02716-3.
Galappaththi, E.K., J.D. Ford, E.M. Bennett and F. Berkes, 2021: Adapting to climate change in small-scale fisheries: Insights from indigenous communities in the global north and south. Environ Sci Policy, 116, 160–170, doi:10.1016/j.envsci.2020.11.009.
Galappaththi, E.K., et al., 2020b: Climate change adaptation in aquaculture. Rev. Aquac. , 12 (4), 2160–2176, doi:10.1111/raq.12427.
Galford, G.L., et al., 2020: Agricultural development addresses food loss and waste while reducing greenhouse gas emissions. Sci. Total Environ. , 699, 134318–134318, doi:10.1016/j.scitotenv.2019.134318.
Gallo, N.D., et al., 2019: Bridging from monitoring to solutions-based thinking: lessons from CalCOFI for understanding and adapting to marine climate change Impacts. Front. Mar. Sci. , 6, 695, doi:10.3389/fmars.2019.00695.
Galvin, K. A., T.A. Beeton, R.B. Boone and S.B. BurnSilver, 2015: Nutritional status of Maasai pastoralists under change. Hum. Ecol. Interdiscip. J. , 43 (3), 411–424, doi:10.1007/s10745-015-9749-x.
Gandini, G.C. and E. Villa, 2003: Analysis of the cultural value of local livestock breeds: a methodology. J. Anim. Breed. Genet. , 120 (1), 1–11, doi:10.1046/j.1439-0388.2003.00365.x.
Gantner, V., et al., 2017: The differences in heat stress resistance due to dairy cattle breed. Mljekarstvo, 67 (2), 112–122, doi:10.15567/mljekarstvo.2017.0203.
García-Martínez, Y.G., et al., 2016: Traditional agroecosystems and global change implications in Mexico. Bulg. J. Agric. Sci. , 22 (4), 548–565.
García de Cortázar-Atauri, I., et al., 2017: Grapevine phenology in France: from past observations to future evolutions in the context of climate change. Oeno One, 51 (2), 115–126, doi:10.20870/oeno-one.2017.51.2.1622.
Garcia, A., et al., 2021: Emancipatory spaces: Opportunities for (re)negotiating gendered subjectivities and enhancing adaptive capacities. Geoforum, 119, 190–205, doi:10.1016/j.geoforum.2020.09.018.
Garibaldi, L.A., et al., 2016: Mutually beneficial pollinator diversity and crop yield outcomes in small and large farms. Science, 351 (6271), 388–391, doi:10.1126/science.aac7287.
Garnett, E.E., et al., 2021: Price of change: does a small alteration to the price of meat and vegetarian options affect their sales?J Environ Psychol, 101589, doi:10.1016/j.jenvp.2021.101589.
Garnett, E.E., et al., 2019: Impact of increasing vegetarian availability on meal selection and sales in cafeterias. Proc. Natl. Acad. Sci. U. S. A. , 116 (42), 20923–20929, doi:10.1073/pnas.1907207116.
Garnett, E.E., et al., 2020: Order of meals at the counter and distance between options affect student cafeteria vegetarian sales. Nat. Food, 1 (8), 485–488, doi:10.1038/s43016-020-0132-8.
Garnett, S.T., et al., 2018: A spatial overview of the global importance of Indigenous lands for conservation. Nat. Sustain. , 1, 369–374, doi:10.1038/s41893-018-0100-6.
Garnett, T., E. Röös and D. Little, 2015: Lean, green, mean, obscene...? What is efficiency? And is it sustainable? Animal production and consumption reconsidered. https://www.fcrn.org.uk/sites/default/files/fcrn_lmgo.pdf. (last accessed January 3, 2021).
Garrabou, J., et al., 2019: Collaborative database to track mass mortality events in the Mediterranean Sea. Front. Mar. Sci. , 6, 707, doi:10.3389/fmars.2019.00707.
Gasco, L., et al., 2018: Fishmeal alternative protein sources for aquaculture feeds. In: Feeds for the Aquaculture Sector: Current Situation and Alternative Sources. Springer International Publishing, Cham, pp. 1–28. ISBN 978-3319779416.
Gattuso, J.P., et al., 2018: Ocean solutions to address climate change and its effects on marine ecosystems. Front. Mar. Sci. , 5, 337, doi:10.3389/fmars.2018.00337.
Gaupp, F., J. Hall, S. Hochrainer-Stigler and S. Dadson, 2020: Changing risks of simultaneous global breadbasket failure. Nat. Clim. Chang. , 10 (1), 54–57, doi:10.1038/s41558-019-0600-z.
Gaupp, F., J. Hall, D. Mitchell and S. Dadson, 2019: Increasing risks of multiple breadbasket failure under 1.5 and 2 degrees C global warming. Agric Syst , 175, 34–45, doi:10.1016/j.agsy.2019.05.010.
Gauthier, S., et al., 2014: Climate change vulnerability and adaptation in the managed Canadian boreal forest. Environ. Rev. , 22 (3), 256–285, doi:10.1139/er-2013-0064.
Gautier, D., B. Locatelli, C. Corniaux and V. Alary, 2016: Global changes, livestock and vulnerability: the social construction of markets as an adaptive strategy. Geogr J, 182 (2), 153–164, doi:10.1111/geoj.12115.
Gebhardt, T., et al., 2014: The more, the better? Water relations of Norway spruce stands after progressive thinning. Agric For Meteorol, 197, 235–243, doi:10.1016/j.agrformet.2014.05.013.
Génova, M., L. Caminero and J. Dochao, 2014: Resin tapping in Pinus pinaster: effects on growth and response function to climate. Eur J For Res, 133, 323–333, doi:10.1007/s10342-013-0764-4.
Gentle, P. and R. Thwaites, 2016: Transhumant pastoralism in the context of socioeconomic and climate change in the mountains of Nepal. Mt Res Dev, 36 (2), 173–182, doi:10.1659/Mrd-Journal-D-15-00011.1.
Gentry, R.R., et al., 2020: Exploring the potential for marine aquaculture to contribute to ecosystem services. Rev. Aquac. , 12 (2), 499–512, doi:10.1111/raq.12328.
Gentry, R.R., et al., 2017: Offshore aquaculture: spatial planning principles for sustainable development. Ecol Evol, 7 (2), 733–743, doi:10.1002/ece3.2637.
Gepts, P., 2014: The contribution of genetic and genomic approaches to plant domestication studies. Curr. Opin. Plant Biol. , 18, 51–59, doi:10.1016/j.pbi.2014.02.001.
Gerard, F., F. Affholder, N. F. Mane and M. Sali, 2020: Impact de différentes politiques publiques sur l’intensification agroécologique et les inégalités de revenu dans le Bassin arachidier du Sénégal. AFD Research Papers.
Gerten, D., et al., 2020: Feeding ten billion people is possible within four terrestrial planetary boundaries. Nat. Sustain. , 3, 200–208, doi:10.1038/s41893-019-0465-1.
Gevaña, D.T., L.D. Camacho and J.M. Pulhin, 2018: Conserving mangroves for their blue carbon: insights and prospects for community-based mangrove management in southeast Asia. In: Threats to Mangrove Forests: Hazards, Vulnerability, and Management [Makowski, C. and C.W. Finkl(eds.)]. Springer, Cham, Switzerland, pp. 579–588. ISBN 978-3319730158.
Ghahramani, A. and D. Bowran, 2018: Transformative and systemic climate change adaptations in mixed crop livestock farming systems. Agric Syst , 164, 236–251, doi:10.1016/j.agsy.2018.04.011.
Ghamkhar, R., C. Hartleb, F. Wu and A. Hicks, 2020: Life cycle assessment of a cold weather aquaponic food production system. J Clean Prod, 244, 118767, doi:10.1016/j.jclepro.2019.118767.
Ghanem, M. E., H. Marrou and T.R. Sinclair, 2015: Physiological phenotyping of plants for crop improvement. Trends Plant Sci. , 20, 139–144, doi:10.1016/j.tplants.2014.11.006.
Ghezzo, M., et al., 2018: Natural resources and climate change: a study of the potential impact on Manila clam in the Venice lagoon. Sci. Total Environ. , 645, 419–430, doi:10.1016/j.scitotenv.2018.07.060.
Ghosh-Jerath, S., et al., 2015: Traditional knowledge and nutritive value of indigenous foods in the Oraon tribal community of Jharkhand: An exploratory cross-sectional study. Ecol. Food. Nutr. , 54 (5), 493–519, doi:10.1080/03670244.2015.1017758.
Giannini, T.C., et al., 2017: Projected climate change threatens pollinators and crop production in Brazil. PLoS ONE, 12 (8), e182274, doi:10.1371/journal.pone.0182274.
Giger, M., et al., 2019: Impacts of large-scale land acquisitions on common-pool resources. In: The Commons in a Glocal World[Haller, T., T. Breu, T. De Moor, C. Rohr and H. Znoj (eds.)]. Routledge, Abingdon, pp. 257-279. ISBN ISBN 978-1-138-48481-8.
Gil, J.D.B., et al., 2017: The resilience of integrated agricultural systems to climate change. Wiley. Interdiscip. Rev. Clim. Change. , 8 (4), e461, doi:10.1002/wcc.461.
Gilbert, M., et al., 2018: Global distribution data for cattle, buffaloes, horses, sheep, goats, pigs, chickens and ducks in 2010. Sci Data, 5, 180227, doi:10.1038/sdata.2018.227.
Giller, K.E., J.A. Andersson, J. Sumberg and J. Thompson, 2017: A golden age for agronomy? In: Agronomy for Development: The Politics of Knowledge in Agricultural Research[Sumberg, J.(ed.)]. Routledge, Oxon, UK, and New York, NY, USA, pp. 150–160. ISBN 978-1138240278.
Gitea, M. A., et al., 2019: Orchard management under the effects of climate change: implications for apple, plum, and almond growing. Environ. Sci. Pollut. Res. Int. , 26 (10), 9908–9915, doi:10.1007/s11356-019-04214-1.
Gjedrem, T. and M. Rye, 2018: Selection response in fish and shellfish: a review. Rev. Aquac. , 10 (1), 168–179, doi:10.1111/raq.12154.
Glemarec, Y., 2017: Addressing the gender differentiated investment risks to climate-smart agriculture. AIMS Agric. Food, 2 (1), 56–74, doi:10.3934/agrfood.2017.1.56.
Glenn, D.M., S.H. Kim, J. Ramirez-Villegas and P. Laderach, 2013: Response of perennial horticultural crops to climate change. In: Horticultural Reviews[Janick, J.(ed.)]. Wiley-Blackwell, pp. 47–130. ISBN 978-1118707371.
Glibert, P.M., 2020: Harmful algae at the complex nexus of eutrophication and climate change. Harmful Algae, 91, 101583, doi:10.1016/j.hal.2019.03.001.
Gliessman, S., 2014: Agroecology: The Ecology of Sustainable Food Systems, 3rd edn., CRC Press, Boca Raton.
Global Commission on Adaptation, 2019: Adapt Now: A Global Call for Leadership on Climate Resilience. Global Center on Adaptation, Rotterdam, the Netherlands, https://gca.org/reports/adapt-now-a-global-call-for-leadership-on-climate-resilience (accessed January 3, 2022). (81 pp).
Glover, D., 2014: Of yield gaps and yield ceilings: making plants grow in particular places. Geoforum, 53, 184–194, doi:10.1016/j.geoforum.2013.06.001.
Godde, C., et al., 2019: Climate change and variability impacts on grazing herds: insights from a system dynamics approach for semi-arid Australian rangelands. Glob Chang Biol, 25 (9), 3091–3109, doi:10.1111/gcb.14669.
Godde, C.M., et al., 2020: Global rangeland production systems and livelihoods at threat under climate change and variability. Environ. Res. Lett. , 15 (4), 44021–44021, doi:10.1088/1748-9326/ab7395.
Godde, C.M., et al., 2021: Impacts of climate change on the livestock food supply chain; a review of the evidence. Glob Food Sec, 28, 100488, doi:10.1016/j.gfs.2020.100488.
Goddek, S., et al., 2015: Challenges of sustainable and commercial aquaponics. Sustainability, 7 (4), 4199–4224, doi:10.3390/su7044199.
Godden, L. and M. Tehan, 2016: REDD+: climate justice and indigenous and local community rights in an era of climate disruption. J. Energy Nat. Resour. Law, 34 (1), 95–108, doi:10.1080/02646811.2016.1121620.
Godfray, H.C., et al., 2010: The future of the global food system. Philos. Trans. R. Soc. Lond. B Biol. Sci. , 365 (1554), 2769–2777, doi:10.1098/rstb.2010.0180.
Golden, C.D., et al., 2016: Nutrition: fall in fish catch threatens human health. Nature, 534 (7607), 317–320, doi:10.1038/534317a.
Golden, C.D., et al., 2011: Benefits of wildlife consumption to child nutrition in a biodiversity hotspot. Proc. Natl. Acad. Sci. U. S. A. , 108 (49), 19653–19656, doi:10.1073/pnas.1112586108.
Goldsmith, K. A., et al., 2019: Scientific considerations for acidification monitoring in the U.S. Mid-Atlantic Region. Estuar. Coast. Shelf Sci. , 225, 106189, doi:10.1016/j.ecss.2019.04.023.
Goldstein, B., M. Hauschild, J. Fernandez and M. Birkved, 2016: Urban versus conventional agriculture, taxonomy of resource profiles: a review. Agron. Sustain. Dev. , 36 (1), 9, doi:10.1007/s13593-015-0348-4.
Goldstein, J.H., et al., 2012: Integrating ecosystem-service tradeoffs into land-use decisions. Proc. Natl. Acad. Sci. U. S. A. , 109 (19), 7565–7570, doi:10.1073/pnas.1201040109.
Gondhalekar, D. and T. Ramsauer, 2017: Nexus city: operationalizing the urban water-energy-food nexus for climate change adaptation in Munich, Germany. Urban Clim, 19, 28–40, doi:10.1016/j.uclim.2016.11.004.
Gonzal, A., T. Pongthanapanic and P. Bueno, 2019: Strengthening Small Aquaculture Entrepreneurs: The Case of a Women’s Association in the Philippines. FAO Fisheries and Aquaculture Circular, Food and Agricultural Organization of the United Nations, Rome, Italy, ISBN 978-9251315446. 26 pp.
Gonzalez-Alzaga, B., et al., 2014: A systematic review of neurodevelopmental effects of prenatal and postnatal organophosphate pesticide exposure. Toxicol. Lett. , 230 (2), 104–121, doi:10.1016/j.toxlet.2013.11.019.
Gonzalez-Varo, J.P., et al., 2013: Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. , 28 (9), 524–530, doi:10.1016/j.tree.2013.05.008.
González de Molina, M., 2020: Strategies for scaling up Agroecological experiences in Europe. Int. J. Agric. Nat. Resour. , 47 (3), 187–203, doi:10.7764/ijanr.v47i3.2257.
Gonzalez, P., C.J. Tucker and H. Sy, 2012: Tree density and species decline in the African Sahel attributable to climate. J. Arid Environ. , 78, 55–64, doi:10.1016/j.jaridenv.2011.11.001.
Goodman, W. and J. Minner, 2019: Will the urban agricultural revolution be vertical and soilless? A case study of controlled environment agriculture in New York City. Land Use Policy, 83, 160–173, doi:10.1016/j.landusepol.2018.12.038.
Gopal, N., et al., 2020: Expanding the horizons for women in fisheries and aquaculture. Gend. Technol. Dev. , 24 (1), 1–9, doi:10.1080/09718524.2020.1736353.
Gorn, L., J. Kleemann and C. Fürst, 2018: Improving the matrix-assessment of ecosystem services provision—the case of regional land use planning under climate change in the region of Halle, Germany. Land, 7 (2), 76, doi:10.3390/land7020076.
Gosling, S., J. Zaherpour and D. Ibarreta, 2018: PESETA III: Climate change impacts on labour productivity. Joint Research Centre (Seville site), Publications Office of the European Union, Luxembourg. Available at: https://publications.jrc.ec.europa.eu/repository/handle/JRC113740 (accessed Juanuary 3, 2021)
Gosnell, H., N. Gill and M. Voyer, 2019: Transformational adaptation on the farm: processes of change and persistence in transitions to ‘climate-smart’ regenerative agriculture. Glob. Environ. Change. , 59, 101965, doi:10.1016/j.gloenvcha.2019.101965.
Gouel, C. and D. Laborde, 2021: The crucial role of domestic and international market-mediated adaptation to climate change. J Environ Econ Manage, 106, doi:10.1016/j.jeem.2020.102408.
Gouse, M., D. Sengupta, P. Zambrano and J.F. Zepeda, 2016: Genetically modified maize: less drudgery for her, more maize for him? Evidence from smallholder maize farmers in South Africa. World Dev, 83, 27–38, doi:10.1016/j.worlddev.2016.03.008.
Govers, G., R. Merckx, B. van Wesemae and K. Van Oost, 2017: Soil conservation in the 21st century: why we need smart agricultural intensification. Soil, 3 (1), 45–59, doi:10.5194/soil-3-45-2017.
Gravert, C. and V. Kurz, 2019: Nudging à la carte: a field experiment on climate-friendly food choice. Behav. Public Policy, 5 (3), 378–395, doi:10.1017/bpp.2019.11.
Gray, S.B., et al., 2016: Intensifying drought eliminates the expected benefits of elevated carbon dioxide for soybean. Nat. Plants, 2 (9), 16132, doi:10.1038/nplants.2016.132.
Greatrex, H., et al., 2015: Scaling Up Index Insurance for Smallholder Farmers: Recent Evidence and Insights. CCAFS Report, CGIAR Research Program on Climate Change, Agriculture and Food Security, Copenhagen, Denmark, https://hdl.handle.net/10568/53101. Accessed 2021 . (32 pp).
Green, R., et al., 2013: The effect of rising food prices on food consumption: systematic review with meta-regression. BMJ, 346, f3703, doi:10.1136/bmj.f3703.
Green, T.J., et al., 2019: Simulated marine heat wave alters abundance and structure of Vibrio populations associated with the Pacific oyster resulting in a mass mortality event. Microb Ecol, 77 (3), 736–747, doi:10.1007/s00248-018-1242-9.
Greene, C., 2018: Broadening understandings of drought - the climate vulnerability of farmworkers and rural communities in California (USA). Environ Sci Policy, 89, 283–291, doi:10.1016/j.envsci.2018.08.002.
Greenhill, L., J.O. Kenter and H. Dannevig, 2020: Adaptation to climate change-related ocean acidification: An adaptive governance approach. Ocean. Coast. Manag. , 191, 105176, doi:10.1016/j.ocecoaman.2020.105176.
Gregorich, E. G., et al., 2017: Litter decay controlled by temperature, not soil properties, affecting future soil carbon. Glob Chang Biol, 23 (4), 1725–1734, doi:10.1111/gcb.13502.
Griffin, T., et al., 2015: Regional self-reliance of the Northeast food system. Renew. Agric. Food Syst. , 30 (4), 349–363, doi:10.1017/S1742170514000027.
Griffith, A.W., O.M. Doherty and C.J. Gobler, 2019: Ocean warming along temperate western boundaries of the Northern Hemisphere promotes an expansion of Cochlodinium polykrikoides blooms. Proc. R. Soc. B. , 286 (1904), 20190340, doi:10.1098/rspb.2019.0340.
Griffith, A.W. and C.J. Gobler, 2017: Transgenerational exposure of North Atlantic bivalves to ocean acidification renders offspring more vulnerable to low pH and additional stressors. Sci Rep, 7 (1), 11394, doi:10.1038/s41598-017-11442-3.
Griffith, A.W. and C.J. Gobler, 2020: Harmful algal blooms: a climate change co-stressor in marine and freshwater ecosystems. Harmful Algae, 91, 101590–101590, doi:10.1016/j.hal.2019.03.008.
Groner, M.L., et al., 2016: Quantifying the influence of salinity and temperature on the population dynamics of a marine ectoparasite. Can. J. Fish. Aquat. Sci. , 73 (8), 1281–1291, doi:10.1139/cjfas-2015-0444.
Gros, C., et al., 2019: Household-level effects of providing forecast-based cash in anticipation of extreme weather events: Quasi-experimental evidence from humanitarian interventions in the 2017 floods in Bangladesh. Int. J. Disaster Risk Reduct. , 41, 101275–101275, doi:10.1016/j.ijdrr.2019.101275.
Grosso, G., et al., 2020: Nutrition in the context of the Sustainable Development Goals. Eur. J. Public. Health. , 30 (Supplement_1), i19–i23, doi:10.1093/eurpub/ckaa034.
Guerra, J., J. Blesh, A.L. Schmitt and H. Wittman, 2017: Pathways to agroecological management through mediated markets in Santa Catarina, Brazil. Elem. Sci. Anthropocene, 5, 67, doi:10.1525/elementa.248.
Guido, Z., C. Knudson, D. Campbell and J. Tomlinson, 2019: Climate information services for adaptation: what does it mean to know the context?Clim. Dev. , 1–13, doi:10.1080/17565529.2019.1630352.
Gulev, S.K., P.W. Thorne, J. Ahn, F.J. Dentener, C.M. Domingues, S. Gerland, D. Gong, D.S. Kaufman, H.C. Nnamchi, J. Quaas, J.A. Rivera, S. Sathyendranath, S.L. Smith, B. Trewin, K. von Schuckmann, and R.S. Vose, 2021: Changing State of the Climate System. In: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. [Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press.
Gumucio, T., J. Hansen, S. Huyer and T. van Huysen, 2020: Gender-responsive rural climate services: a review of the literature. Clim. Dev. , 12 (3), 241–254, doi:10.1080/17565529.2019.1613216.
Gunathilaka, R.P.D., J.C.R. Smart and C.M. Fleming, 2018: Adaptation to climate change in perennial cropping systems: options, barriers and policy implications. Environ Sci Policy, 82, 108–116, doi:10.1016/j.envsci.2018.01.011.
Guo, H.B., et al., 2018: Estimating realized heritability for growth in Zhikong scallop (Chlamys farreri) using genome-wide complex trait analysis. Aquaculture, 497, 103–108, doi:10.1016/j.aquaculture.2018.07.046.
Guo, Y., et al., 2017: Prediction of the potential geographic distribution of the ectomycorrhizal mushroom Tricholoma matsutake under multiple climate change scenarios. Sci Rep, 7, 46221, doi:10.1038/srep46221.
Guo, Y. and Y.J. Shen, 2015: Quantifying water and energy budgets and the impacts of climatic and human factors in the Haihe River Basin, China: 2. Trends and implications to water resources. J Hydrol, 527, 251–261, doi:10.1016/j.jhydrol.2015.04.071.
Guodaar, L., et al., 2019: How do climate change adaptation strategies result in unintended maladaptive outcomes? Perspectives of tomato farmers. Int. J. Veg. Sci. , 26 (1), 15–31, doi:10.1080/19315260.2019.1573393.
Gupta, A., 1998: Postcolonial Developments: Agriculture in the Making of Modern India. Duke University Press, Durham, NC, USA, ISBN 978-0822322139. 432 pp.
Gupta, P. and N. Sen, 1980: Food consumption and nutrition of regional tribes of India. Ecol. Food. Nutr. , 9 (2), 93–108, doi:10.1080/03670244.1980.9990587.
Gupta, R., E. Somanathan and S. Dey, 2016: Global warming and local air pollution have reduced wheat yields in India. Clim. Change, 140 (3-4), 593–604, doi:10.1007/s10584-016-1878-8.
Gurung, K., H. Bhandari and T. Paris, 2016: Transformation from Rice Farming to Commercial Aquaculture in Bangladesh: Implications for Gender, Food Security, and Livelihood. Gend. Technol. Dev. , 20 (1), 49–80, doi:10.1177/0971852415618747.
Gutierrez, A.P., O. Matika, T.P. Bean and R.D. Houston, 2018: Genomic selection for growth traits in Pacific oyster (Crassostrea gigas): potential of low-density Marker Panels for breeding value prediction. Front. Genet. , 9, 391, doi:10.3389/fgene.2018.00391.
Gutsch, M., et al., 2018: Balancing trade-offs between ecosystem services in Germany’s forests under climate change. Environ. Res. Lett. , 13 (4), 45012, doi:10.1088/1748-9326/aab4e5.
Gyapong, A.Y., 2020: How and why large scale agricultural land investments do not create long-term employment benefits: A critique of the ‘state’ of labour regulations in Ghana. Land Use Policy, 95 (April), 104651–104651, doi:10.1016/j.landusepol.2020.104651.
Haapala, A., 2018: A feminist analysis of oppressive and emancipatory potentialities within technology-prioritized climate change adaptation intervention: a case study from Phailom community in Laos. In: Gender dimension of climate change research in agriculture (Case studies in Southeast Asia) [Paris, T.R. and M. E. Rola-Rubzen(eds.)]. Southeast Asian Regional Center for Graduate Study and Research in Agriculture, CGIAR Research Program on Climate Change, Agriculture, and Food Security, Laguna, Philippines and Wageningen, the Netherlands, pp. 123–140. ISBN 978-9715602716.
Haasnoot, M., et al., 2020: Defining the solution space to accelerate climate change adaptation. Reg Environ Change, 20 (2), doi:10.1007/s10113-020-01623-8.
Häberli, C. and F. Smith, 2014: Food Security and Agri-Foreign Direct Investment in Weak States: Finding the Governance Gap to Avoid ‘Land Grab’. Mod Law Rev, 77 (2), 189–222, doi:10.1111/1468-2230.12062.
Habermann, E., et al., 2019: Warming and water deficit impact leaf photosynthesis and decrease forage quality and digestibility of a C4 tropical grass. Physiol Plant , 165 (2), 383–402, doi:10.1111/ppl.12891.
Hadjichambis, A., et al., 2008: Wild and semi-domesticated food plant consumption in seven circum-Mediterranean areas. Int. J. Food Sci. Nutr. , 59 (5), 383–414, doi:10.1080/09637480701566495.
Hagenlocher, M., et al., 2019: Drought vulnerability and risk assessments: state of the art, persistent gaps, and research agenda. Environ. Res. Lett. , 14 (8), doi:10.1088/1748-9326/ab225d.
Haglund, E., J. Ndjeunga, L. Snook and D. Pasternak, 2011: Dry land tree management for improved household livelihoods: farmer managed natural regeneration in Niger. J Environ Manage, 92 (7), 1696–1705, doi:10.1016/j.jenvman.2011.01.027.
Hai, N.T., B. Dell, V.T. Phuong and R.J. Harper, 2020: Towards a more robust approach for the restoration of mangroves in Vietnam. Ann. For. Sci. , 77 (1), 18, doi:10.1007/s13595-020-0921-0.
Hailegiorgis, A., A. Crooks and C. Cioffi-Revilla, 2018: An agent-based model of rural households’ adaptation to climate change. J. Artif. Soc. Soc. Simul. , 21 (4), doi:10.18564/jasss.3812.
Haileslassie, A., et al., 2009: Livestock water productivity in mixed crop-livestock farming systems of the Blue Nile basin: assessing variability and prospects for improvement. Agric Syst , 102 (1), 33–40, doi:10.1016/j.agsy.2009.06.006.
Haji, J. and B. Legesse, 2017: Impact of sedentarization program on the livelihood and food security of Ethiopian pastoralists. J. Arid Environ. , 136, 45–53, doi:10.1016/j.jaridenv.2016.10.007.
Hajjar, R., G. Engbring and K. Kornhauser, 2021: The impacts of REDD plus on the social-ecological resilience of community forests. Environ. Res. Lett. , 16 (2), doi:10.1088/1748-9326/abd7ac.
Hak, S., J. McAndrew and A. Neef, 2018: Impact of government policies and corporate land grabs on indigenous people’s access to common lands and livelihood resilience in Northeast Cambodia. Land, 7 (4), doi:10.3390/land7040122.
Hakkenberg, R., et al., 2008: Temperature sensitivity of the turnover times of soil organic matter in forests. Ecol Appl, 18 (1), 119–131, doi:10.1890/06-1034.1.
Hall, A., A.J. Mathews and B.P. Holzapfel, 2016: Potential effect of atmospheric warming on grapevine phenology and post-harvest heat accumulation across a range of climates. Int. J. Biometeorol. , 60 (9), 1405–1422, doi:10.1007/s00484-016-1133-z.
Hallegatte, S., et al., 2018: The Economics of (and Obstacles to) Aligning Development and Climate Change Adaptation. Global Center on Adaptation, Rotterdam and Washington, DC, Available at: https://gca.org/reports/the-economics-of-and-obstacles-to-aligning-development-and-climate-change-adaptation/ (accessed January 3, 2021) Haller, T., 2019: The different meanings of land in the age of neoliberalism: Theoretical reflections on commons and resilience grabbing from a social anthropological perspective. Land, 8 (7), doi:10.3390/land8070104.
Haller, T., F. Käser and M. Ngutu, 2020: Does commons grabbing lead to resilience grabbing? The anti-politics machine of neo-liberal agrarian development and local responses. Land, 9 (7), doi:10.3390/land9070220.
Halpern, B.S., et al., 2019: Putting all foods on the same table: achieving sustainable food systems requires full accounting. Proc. Natl. Acad. Sci. U. S. A. , 116 (37), 18152–18156, doi:10.1073/pnas.1913308116.
Halvey, M.R., et al., 2020: Beyond backyard chickens: A framework for understanding municipal urban agriculture policies in the United States. Food Policy, 102013–102013, doi:10.1016/j.foodpol.2020.102013.
Hambrey, J., 2017: The 2030 Agenda and the Sustainable Development Goals: The Challenge for Aquaculture Development and Management . FAO Fisheries and Aquaculture Circular, Vol. 1141. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978925109928. 62 pp.
Hamilton, L.C., et al., 2016: Alaska. Popul Environ, 38 (2), 115–133, doi:10.1007/s11111-016-0259-6.
Hammill, M.O., 2009: Ringed seal: Pusa hispida. In: Encyclopedia of Marine Mammals[Perrin, W.F., B. Würsig and J.G.M. Thewissen(eds.)], 2nd edn. Academic, Burlington, MA, USA, pp. 972–974. ISBN 97801237355539.
Hanaček, K. and B. Rodríguez-Labajos, 2018: Impacts of land-use and management changes on cultural agroecosystem services and environmental conflicts—a global review. Glob. Environ. Chang. , 50, 41–59, doi:10.1016/j.gloenvcha.2018.02.016.
Handisyde, N., T.C. Telfer and L.G. Ross, 2017: Vulnerability of aquaculture-related livelihoods to changing climate at the global scale. Fish. Fish. , 18 (3), 466–488, doi:10.1111/faf.12186.
Hanich, Q., et al., 2018: Small-scale fisheries under climate change in the Pacific Islands region. Mar. Policy. , 88, 279–284, doi:10.1016/j.marpol.2017.11.011.
Hank, T.B., et al., 2019: Spaceborne imaging spectroscopy for sustainable agriculture: contributions and challenges. Surv Geophys, 40 (3), 515–551, doi:10.1007/s10712-018-9492-0.
Hannah, L., et al., 2017: Regional modeling of climate change impacts on smallholder agriculture and ecosystems in Central America. Clim Change, 141 (1), 29–45, doi:10.1007/s10584-016-1867-y.
Hansel, M.C., et al., 2020: Ocean warming and acidification may drag down the commercial Arctic cod fishery by 2100. PLoS ONE, 15 (4), e231589, doi:10.1371/journal.pone.0231589.
Hansen, B.B., et al., 2013: Climate events synchronize the dynamics of a resident vertebrate community in the high Arctic. Science, 339 (6117), 313–315, doi:10.1126/science.1226766.
Hansen, J., et al., 2019a: Climate risk management and rural poverty reduction. Agric Syst , 172, 28–46, doi:10.1016/j.agsy.2018.01.019.
Hansen, P.G., M. Schilling and M.S. Malthesen, 2019b: Nudging healthy and sustainable food choices: three randomized controlled field experiments using a vegetarian lunch-default as a normative signal. J. Public Health, 43 (2), 392–397, doi:10.1093/pubmed/fdz154.
Haque, E., et al., 2018: Application of CRISPR/Cas9 genome editing technology for the improvement of crops cultivated in tropical climates: recent progress, prospects, and challenges. Front. Plant Sci. , 9, 617, doi:10.3389/fpls.2018.00617.
Hariharan, V.K., et al., 2018: Does climate-smart village approach influence gender equality in farming households? A case of two contrasting ecologies in India. Clim Change, 158 (1), 77–90, doi:10.1007/s10584-018-2321-0.
Harland, J., 2019: The origins of aquaculture. Nat Ecol Evol, 3 (10), 1378–1379, doi:10.1038/s41559-019-0966-3.
Harle, K.J., S.M. Howden, L.P. Hunt and M. Dunlop, 2007: The potential impact of climate change on the Australian wool industry by 2030. Agric Syst , 93 (1-3), 61–89, doi:10.1016/j.agsy.2006.04.003.
Harper, S., et al., 2020: Valuing invisible catches: Estimating the global contribution by women to small-scale marine capture fisheries production. PLoS ONE, 15 (3), e228912, doi:10.1371/journal.pone.0228912.
Harris, J., et al., 2021: Fruits and Vegetables for Healthy Diets: Priorities for Food System Research and Action. Food Systems Summit Briefs, The Scientific Group for the Food Systems Summit, (11 pp) Available at: https://sc-fss2021.org/wp-content/uploads/2021/04/FSS_Brief_Fruits_Vegetables.pdf (accessed August 3, 2021)Harrison, H.L., S. Rybraten and O. Aas, 2018: Hatching knowledge: a case study on the hybridization of local ecological knowledge and scientific knowledge in small-scale Atlantic salmon (Salmo salar) cultivation in Norway. Hum Ecol, 46 (4), 449–459, doi:10.1007/s10745-018-0001-3.
Harrison, M.T., B.R. Cullen and D. Armstrong, 2017: Management options for dairy farms under climate change: Effects of intensification, adaptation and simplification on pastures, milk production and profitability. Agric Syst , 155, 19–32, doi:10.1016/j.agsy.2017.04.003.
Harrison, M.T., B.R. Cullen and R.P. Rawnsley, 2016: Modelling the sensitivity of agricultural systems to climate change and extreme climatic events. Agric Syst , 148, 135–148, doi:10.1016/j.agsy.2016.07.006.
Harrod, C., A. Ramírez, J. Valbo-Jørgensen and S. Funge-Smith, 2018a: Current anthropogenic stress and projected effect of climate change on global inland fisheries. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agricultural Organization of the United Nations, Rome, Italy, pp. 393–448. ISBN 978-9251306079.
Harrod, C., A. Ramírez, J. Valbo-Jørgensen and S. Funge-Smith, 2018b: How climate change impacts inland fisheries. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 375–392. ISBN 978-9251306079.
Harrod, C., F. Simmance, S. Funge-Smith and J. Valbo-Jørgensen, 2018c: Options and opportunities for supporting inland fisheries to cope with climate change adaptation in other sectors. In: Impacts of climate change on fisheries and aquaculture: synthesis of current knowledge, adaptation and mitigation options[Barange, M., et al.(ed.)]. FAO, Rome, pp. 567–584.
Hartiningtias, D., P.Z. Fulé and A.A. Gunawan, 2020: Wildfire effects on forest structure of Pinus merkusii in Sumatra, Indonesia. For. Ecol. Manag. , 457, doi:10.1016/j.foreco.2019.117660.
Hasan, M.R. and S. Soto, 2017: Improving Feed Conversion Ratio and Its Impact on Reducing Greenhouse Gas Emissions in Aquaculture. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251098868. 33 pp.
Hasegawa, T., et al., 2018: Risk of increased food insecurity under stringent global climate change mitigation policy. Nat. Clim. Chang. , 8 (8), 699–703, doi:10.1038/s41558-018-0230-x.
Hasegawa, T., et al., 2016: Rice free-air carbon dioxide enrichment studies to improve assessment of climate change effects on rice agriculture. In: Improving Modeling Tools to Assess Climate Change Effects on Crop Response[Hatfield, J.L. and D. Fleisher(eds.)]. American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, Madison, WI, USA, pp. 45–68. ISBN 978-0891183518.
Hasegawa, T., et al., 2021a: Extreme climate events increase risk of global food insecurity and adaptation needs. Nat. Food, 2 (8), 587–595, doi:10.1038/s43016-021-00335-4.
Hasegawa, T., et al., 2021b: A global dataset for the projected impacts of climate change on four major crops. Biorxiv, doi:10.1101/2021.05.27.444762.
Hassan, B., et al., 2020: Impact of climate change on freshwater ecosystem and its sustainable management. In: Fresh Water Pollution Dynamics and Remediation[Qadri, H., R.A. Bhat, M. A. Mehmood and G.H. Dar(eds.)]. Springer Nature, Singapore, Singapore. ISBN 978-9811382785.
Hasselberg, A.E., et al., 2020: Fish for food and nutrition security in Ghana: challenges and opportunities. Glob. Food Secur. Policy Econ. Environ. , 26, 100380, doi:10.1016/j.gfs.2020.100380.
Hassen, A., et al., 2017: Potential use of forage-legume intercropping technologies to adapt to climate-change impacts on mixed crop-livestock systems in Africa: a review. Reg Environ Change, 17 (6), 1713–1724, doi:10.1007/s10113-017-1131-7.
Havemann, T., C. Negra and F. Werneck, 2020: Blended finance for agriculture: exploring the constraints and possibilities of combining financial instruments for sustainable transitions. Agric Human Values, 1, 1–12, doi:10.1007/s10460-020-10131-8.
Havet, A., et al., 2014: Review of livestock farmer adaptations to increase forages in crop rotations in western France. Agric Ecosyst Environ, 190, 120–127, doi:10.1016/j.agee.2014.01.009.
Havlik, P., et al., 2014: Climate change mitigation through livestock system transitions. Proc. Natl. Acad. Sci. U. S. A. , 111 (10), 3709–3714, doi:10.1073/pnas.1308044111.
Haworth, M., Y. Hoshika and D. Killi, 2016: Has the impact of rising CO2 on plants been exaggerated by meta-analysis of free air CO2 enrichment studies?Front. Plant Sci. , 7, 1153, doi:10.3389/fpls.2016.01153.
He, W., et al., 2019: Assessing alfalfa production under historical and future climate in eastern Canada: DNDC model development and application. Environ. Model. Softw. , 122, 104540–104540, doi:10.1016/j.envsoft.2019.104540.
Heck, V., D. Gerten, W. Lucht and A. Popp, 2018: Biomass-based negative emissions difficult to reconcile with planetary boundaries. Nat. Clim. Chang. , 8 (2), 151–155, doi:10.1038/s41558-017-0064-y.
Heffernan, C., 2018: Climate change and multiple emerging infectious diseases. Vet. J. , 234, 43–47, doi:10.1016/j.tvjl.2017.12.021.
Heide-Jørgensen, M.P., 2018: Narwhal: Monodon monoceros. In: Encyclopedia of Marine Mammals[Perrin, W.F., B. Würsig and J.G.M. Thewissen(eds.)], 2nd edn. Academic, Burlington, MA, USA, pp. 627–631. ISBN 97801237355539.
Heinemann, J., D.S. Coray and D.S. Thaler, 2018: Exploratory fact-finding scoping study on “digital sequence information” on genetic resources for food and agriculture. Background Study Paper, Vol. 68. UN FAO, Rome.
Heinemann, J.A., et al., 2014: Sustainability and innovation in staple crop production in the US Midwest. Int. J. Agric. Sustain. , 12 (1), 71–88, doi:10.1080/14735903.2013.806408.
Heino, M., et al., 2020: A multi-model analysis of teleconnected crop yield variability in a range of cropping systems. Earth Syst. Dyn. , 11, 113–128, doi:10.5194/esd-11-113-2020.
Hejazi, M., et al., 2014: Long-term global water projections using six socioeconomic scenarios in an integrated assessment modeling framework. Technol Forecast Soc Change, 81, 205–226, doi:10.1016/j.techfore.2013.05.006.
Hellberg, R.S. and E. Chu, 2016: Effects of climate change on the persistence and dispersal of foodborne bacterial pathogens in the outdoor environment: A review. Crit. Rev. Microbiol. , 42 (4), 548–572, doi:10.3109/1040841X.2014.972335.
Hempel, S. and C. Menz, 2019: Indoor climate projections for European cattle barns, V1. Mendeley Data, https://doi.org/10.17632/tjp8h523p7.1. Accessed 2021.
Henchion, M., et al., 2017: Future protein supply and demand: strategies and factors influencing a sustainable equilibrium. Foods, 6 (7), 53, doi:10.3390/foods6070053.
Henders, S., U.M. Persson and T. Kastner, 2015: Trading forests: land-use change and carbon emissions embodied in production and exports of forest-risk commodities. Environ. Res. Lett. , 10 (12), 125012, doi:10.1088/1748-9326/10/12/125012.
Henderson, B., et al., 2018: The economic potential of residue management and fertilizer use to address climate change impacts on mixed smallholder farmers in Burkina Faso. Agric Syst , 167, 195–205, doi:10.1016/j.agsy.2018.09.012.
Hendrix, C.S., 2018: Searching for climate-conflict links. Nat. Clim. Chang. , 8 (3), 190–191, doi:10.1038/s41558-018-0083-3.
Hennon, G.M.M. and S.T. Dyhrman, 2020: Progress and promise of omics for predicting the impacts of climate change on harmful algal blooms. Harmful Algae, 91, 101587–101587, doi:10.1016/j.hal.2019.03.005.
Henry, B.K., R.J. Eckard and K. A. Beauchemin, 2018: Review: Adaptation of ruminant livestock production systems to climate changes. Animal, 12 (s2), s445–s456, doi:10.1017/S1751731118001301.
Henry, R.C., et al., 2019: The role of global dietary transitions for safeguarding biodiversity. Glob. Environ. Chang. , 58, 101956–101956, doi:10.1016/j.gloenvcha.2019.101956.
Herath, S., K. Tsusaka and J. Diwa, 2015: Assessment on the Feasibility of REDD+ in Nagacadan Rice Terraces of Ifugao and Its Muyong Forest . Rice Terrace Farming Systems Working Paper Series, United Nations University Institute for the Advanced Study of Sustainability, Tokyo, Japan, http://collections.unu.edu/eserv/UNU:3335/Assessment_on_the_feasibility_of_REDD.pdf. (accessed December 28, 2021) . (8 pp).
Herbert, E.R., et al., 2015: A global perspective on wetland salinization: ecological consequences of a growing threat to freshwater wetlands. Ecosphere, 6 (10), 206, doi:10.1890/Es14-00534.1.
Herbut, P., S. Angrecka, D. Godyń and G. Hoffmann, 2019: The Physiological and Productivity Effects of Heat Stress in Cattle: A Review. Ann. Animal Sci. , 19 (3), 579–593, doi:10.2478/aoas-2019-0011.
Herforth, A. and S. Ahmed, 2015: The food environment, its effects on dietary consumption, and potential for measurement within agriculture-nutrition interventions. Food Secur, 7 (3), 505–520, doi:10.1007/s12571-015-0455-8.
Herman-Mercer, N.M., et al., 2019: Vulnerability of subsistence systems due to social and environmental change: a case study in the Yukon-Kuskokwim Delta, Alaska. Arctic, 72 (3), 258–272, doi:10.14430/arctic68867.
Herman-Mercer, N.M., R.A. Loehman, R.C. Toohey and C. Paniyak, 2020: Climate- and disturbance-driven changes in subsistence berries in coastal Alaska: indigenous knowledge to inform ecological inference. Hum Ecol, 48 (1), 85–99, doi:10.1007/s10745-020-00138-4.
Hernández-Morcillo, M., et al., 2018: Scanning agroforestry-based solutions for climate change mitigation and adaptation in Europe. Environ Sci Policy, 80, 44–52, doi:10.1016/j.envsci.2017.11.013.
Hernroth, B.E. and S.P. Baden, 2018: Alteration of host-pathogen interactions in the wake of climate change - Increasing risk for shellfish associated infections?Environ. Res. , 161, 425–438, doi:10.1016/j.envres.2017.11.032.
Herrero, M., et al., 2020: Innovation can accelerate the transition towards a sustainable food system. Nat. Food, 1 (5), 266–272, doi:10.1038/s43016-020-0074-1.
Herrero, M., et al., 2017: Farming and the geography of nutrient production for human use: a transdisciplinary analysis. Lancet Planet. Health, 1 (1), E33–E42, doi:10.1016/s2542-5196(17)30007-4.
Herrero, M., et al., 2015: Livestock and the environment: what have we learned in the past decade?Annu. Rev. Environ. Resour. , 40 (1), 177–202, doi:10.1146/annurev-environ-031113-093503.
Herring, S.C., et al., 2018: Explaining Extreme Events of 2016 from a Climate Perspective. Bull. Am. Meterol. Soc. , 99 (1), S1–S157, doi:10.1175/BAMS-ExplainingExtremeEvents2016.1.
Hertel, T.W. and U.L.C. Baldos, 2016: Attaining food and environmental security in an era of globalization. Glob. Environ. Change. , 41, 195–205, doi:10.1016/j.gloenvcha.2016.10.006.
Hertel, T.W. and C.Z. de Lima, 2020: Viewpoint: climate impacts on agriculture: searching for keys under the streetlight. Food Policy, 95, doi:10.1016/j.foodpol.2020.101954.
Heslin, A., 2021: Riots and resources: How food access affects collective violence. J Peace Res, 58 (2), 199–214, doi:10.1177/0022343319898227.
Hickey, G.M. and N. Unwin, 2020: Addressing the triple burden of malnutrition in the time of COVID-19 and climate change in small island developing states: what role for improved local food production?Food Secur, 12, 1–5, doi:10.1007/s12571-020-01066-3.
Hickman, J.E., et al., 2017: Nonlinear response of nitric oxide fluxes to fertilizer inputs and the impacts of agricultural intensification on tropospheric ozone pollution in Kenya. Glob Chang Biol, 23 (8), 3193–3204, doi:10.1111/gcb.13644.
Hicks, C.C., et al., 2019: Harnessing global fisheries to tackle micronutrient deficiencies. Nature, 574 (7776), 95–98, doi:10.1038/s41586-019-1592-6.
Hicks Pries, C.E., et al., 2018: Response to comment on “The whole-soil carbon flux in response to warming”. Science, 359 (6378), eaao457, doi:10.1126/science.aao0457.
Hicks Pries, C.E., C. Castanha, R.C. Porras and M.S. Torn, 2017: The whole-soil carbon flux in response to warming. Science, 355 (6332), 1420–1423, doi:10.1126/science.aal1319.
Hidalgo, H.A., 2015: Vulnerability assessment of agri-ecotourism communities as influenced by climate change. Int. J. Adv. Sci. Eng. Inf. Technol. , 5 (6), 379, doi:10.18517/ijaseit.5.6.553.
Hidalgo, M., V. Mihneva, M. Vasconcellos and M. Bernal, 2018: Climate change impacts, vulnerabilities and adaptations: Mediterranean Sea and the Black Sea marine fisheries. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 139–158. ISBN 978-9251306079.
Hill, S., 2014: Considerations for enabling the ecological redesign of organic and conventional agriculture: A social ecology and psychosocial perspective. In: Organic Farming, Prototype for Sustainable Agricultures[Bellon, S. and S. Penvern(eds.)]. Springer Science+Business Media, Dordrecht, the Netherlands, pp. 401–422. ISBN 978-94-007-7927-3.
Hilmi, N., et al., 2014: Exposure of Mediterranean countries to ocean acidification. Water, 6 (6), 1719–1744, doi:10.3390/w6061719.
Hiltner, U., et al., 2021: Climate change alters the ability of neotropical forests to provide timber and sequester carbon. For. Ecol. Manag. , 492, 119166, doi:10.1016/j.foreco.2021.119166.
Hitchcock, R., 2016: From Local to Global: Perceptions and Realities of Environmental Change Among Kalahari San. In: Anthropology and Climate Change: From Encounters to Actions[Crate, S.A. and M. Nuttall(eds.)]. Routledge, Oxon, UK and New York, NY, USA, pp. 250–262. ISBN 978-1598743333.
Hixson, S.M. and M.T. Arts, 2016: Climate warming is predicted to reduce omega-3, long-chain, polyunsaturated fatty acid production in phytoplankton. Glob Chang Biol, 22 (8), 2744–2755, doi:10.1111/gcb.13295.
Hlásny, T., et al., 2021: Devastating outbreak of bark beetles in the Czech Republic: drivers, impacts, and management implications. For. Ecol. Manag. , 490, doi:10.1016/j.foreco.2021.119075.
HLPE, 2013: Investing in Smallholder Agriculture for Food Security. HLPE Report, High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security, Rome, Italy, <Go to WoS>://WOS:000406485000003. Accessed 2021 . (112 pp).
HLPE, 2014: Sustainable Fisheries and Aquaculture for Food Security and Nutrition. HLPE Report, High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security, Rome, Italy, http://www.fao.org/3/i3844e/i3844e.pdf. Accessed 2021 . (119 pp).
HLPE, 2017a: Nutrition and Food Systems. HLPE Report, High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security, http://www.fao.org/3/i7846e/i7846e.pdf. Accessed 2021 . (152 pp).
HLPE, 2017b: Sustainable Forestry for Food Security and Nutrition. HLPE Report, High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security, Rome, Italy, http://www.fao.org/3/i7395e/i7395e.pdf. Accessed 2021 . (137 pp).
HLPE, 2019: Agroecological and Other Innovative Approaches for Sustainable Agriculture and Food Systems that Enhance Food Security and Nutrition. HLPE Report, High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security, Rome, Italy, http://www.fao.org/3/ca5602en/ca5602en.pdf. Accessed 2021 . (163 pp).
HLPE, 2020: Impacts of COVID-19 on Food Security and Nutrition: Developing Effective Policy Responses to Address the Hunger and Malnutrition Pandemic. HLPE Issues Paper, High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security, Rome, Italy, https://doi.org/10.4060/cb1000en. Accessed 2021 . (22 pp).
HLPE, 2021: Promoting youth engagement and employment in agriculture and food systems. High Level Panel of Experts (HLPE) of UN Committee for World Food Security Food and Agrcultural Organization (FAO).
Hobbs, N.T., et al., 2008: Fragmentation of rangelands: implications for humans, animals, and landscapes. Glob. Environ. Change. , 18 (4), 776–785, doi:10.1016/j.gloenvcha.2008.07.011.
Hobbs, R.J., L.E. Valentine, R.J. Standish and S.T. Jackson, 2018: Movers and stayers: novel assemblages in changing environments. Trends Ecol. Evol. , 33 (2), 116–128, doi:10.1016/j.tree.2017.11.001.
Hobday, A.J., et al., 2017: National climate change adaptation research plan marine biodiversity and resources: update 2017. National Climate Change Adaptation Research Facility, Gold Coast. 75 pp.
Hobday, A.J., et al., 2018: A framework for combining seasonal forecasts and climate projections to aid risk management for fisheries and aquaculture. Front. Mar. Sci. , 5, doi:10.3389/fmars.2018.00137.
Hochman, Z., D.L. Gobbett and H. Horan, 2017: Climate trends account for stalled wheat yields in Australia since 1990. Glob. Change Biol. , 23 (5), 2071–2081, doi:10.1111/gcb.13604.
Hochrainer-Stigler, S. and K. Reiter, 2021: Risk-Layering for Indirect Effects. Int. J. Disaster Risk Sci. , 12(5), 770-778, doi:10.1007/s13753-021-00366-2.
Hodges, R.J., J.C. Buzby and B. Bennett, 2011: Postharvest losses and waste in developed and less developed countries: opportunities to improve resource use. J. Agric. Sci. , 149, 37–45, doi:10.1017/S0021859610000936.
Hoegh-Guldberg, O., D. Jacob, M. Taylor, M. Bindi, S. Brown, I. Camilloni, A. Diedhiou, R. Djalante, K.L. Ebi, F. Engelbrecht, J. Guiot, Y. Hijioka, S. Mehrotra, A. Payne, S.I. Seneviratne, A. Thomas, R. Warren, and G. Zhou, 2018: Impacts of 1.5°C Global Warming on Natural and Human Systems. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty[Masson-Delmotte, V., P. Zhai, H. O. Pörtner, D. Roberts, J. Skea, P. R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J. B. R. Matthews, Y. Chen, X. Zhou, M. I. Gomis, E. Lonnoy, T. Maycock, M. Tignor and T. Waterfield (eds.)].
Hoffman, J.S., V. Shandas and N. Pendleton, 2020: The effects of historical housing policies on resident exposure to intra-urban heat: a study of 108 US urban areas. Climate, 8 (1), 12, doi:10.3390/cli8010012.
Hofmeijer, I., et al., 2013: Community vulnerability to the health effects of climate change among indigenous populations in the Peruvian Amazon: a case study from Panaillo and Nuevo Progreso. Mitig Adapt Strateg Glob Change, 18 (7), 957–978, doi:10.1007/s11027-012-9402-6.
Holland, R.A., et al., 2015: A synthesis of the ecosystem services impact of second generation bioenergy crop production. Renew. Sustain. Energy Rev. , 46, 30–40, doi:10.1016/j.rser.2015.02.003.
Holmes, I., K.R. Kirby and C. Potvin, 2017: Agroforestry within REDD plus : experiences of an indigenous Embera community in Panama. Agrofor. Syst. , 91 (6), 1181–1197, doi:10.1007/s10457-016-0003-3.
Holsman, K.K., et al., 2020: Ecosystem-based fisheries management forestalls climate-driven collapse. Nat Commun, 11 (1), 4579, doi:10.1038/s41467-020-18300-3.
Holsman, K.K., et al., 2019: Towards climate resiliency in fisheries management. ICES J. Mar. Sci. , 76 (5), 1368–1378, doi:10.1093/icesjms/fsz031.
Holt-Giménez, E., A. Shattuck and I. Van Lammeren, 2021: Thresholds of resistance: agroecology, resilience and the agrarian question. J Peasant Stud, 48 (4), 715–733, doi:10.1080/03066150.2020.1847090.
Honkaniemi, J., W. Rammer and R. Seidl, 2020: Norway spruce at the trailing edge: the effect of landscape configuration and composition on climate resilience. Landsc Ecol, 35 (3), 591–606, doi:10.1007/s10980-019-00964-y.
Hopping, K. A., S.M. Chignell and E.F. Lambin, 2018: The demise of caterpillar fungus in the Himalayan region due to climate change and overharvesting. Proc. Natl. Acad. Sci. U.S.A. , 115 (45), 11489–11494, doi:10.1073/pnas.1811591115.
Hopping, K. A., C.R. Yangzong and J.A. Klein, 2016: Local knowledge production, transmission, and the importance of village leaders in a network of Tibetan pastoralists coping with environmental change. Ecol. Soc. , 21 (1), 25, doi:10.5751/Es-08009-210125.
Horst, M., N. McClintock and L. Hoey, 2017: The intersection of planning, urban agriculture, and food justice: a review of the literature. J. Am. Plan. Assoc. , 83 (3), 277–295, doi:10.1080/01944363.2017.1322914.
Horstkotte, T., et al., 2017: Human-animal agency in reindeer management: Sami herders’ perspectives on vegetation dynamics under climate change. Ecosphere, 8 (9), e1931, doi:10.1002/ecs2.1931.
Hosonuma, N., et al., 2012: An assessment of deforestation and forest degradation drivers in developing countries. Environ. Res. Lett. , 7 (4), 44009, doi:10.1088/1748-9326/7/4/044009.
Hossain, M.A.R., M. Ahmed, E. Ojea and J.A. Fernandes, 2018: Impacts and responses to environmental change in coastal livelihoods of south-west Bangladesh. Sci. Total Environ. , 637-638, 954–970, doi:10.1016/j.scitotenv.2018.04.328.
Houser, M. and D. Stuart, 2019: An accelerating treadmill and an overlooked contradiction in industrial agriculture: climate change and nitrogen fertilizer. J. Agrar. Chang. , 20 (2), 215–237, doi:10.1111/joac.12341.
Houston, R.D., et al., 2020: Harnessing genomics to fast-track genetic improvement in aquaculture. Nat. Rev. Genet. , 21 (7), 389–409, doi:10.1038/s41576-020-0227-y.
Hristov, A.N., et al., 2018: Climate change effects on livestock in the Northeast US and strategies for adaptation. Clim Change, 146 (1-2), 33–45, doi:10.1007/s10584-017-2023-z.
Hsiang, S., et al., 2017: Estimating economic damage from climate change in the United States. Science, 356 (6345), 1362–1369, doi:10.1126/science.aal4369.
Hu, H. and L. Xiong, 2014: Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. , 65 (1), 715–741, doi:10.1146/annurev-arplant-050213-040000.
Hua, K., et al., 2019: The Future of Aquatic Protein: Implications for Protein Sources in Aquaculture Diets. One Earth, 1 (3), 316–329, doi:10.1016/j.oneear.2019.10.018.
Huaman, E.S. and L.A. Valdiviezo, 2014: Indigenous knowledge and education from the Quechua community to school: beyond the formal/non-formal dichotomy. Int. J. Qual. Stud. Educ. , 27 (1), 65–87, doi:10.1080/09518398.2012.737041.
Huang, J. and H. Hao, 2020: Effects of climate change and crop planting structure on the abundance of cotton bollworm, Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). Ecol Evol, 10 (3), 1324–1338, doi:10.1002/ece3.5986.
Huang, J., A.E. Hartemink and C.J. Kucharik, 2021: Soil-dependent responses of US crop yields to climate variability and depth to groundwater. Agric Syst , 190, 103085, doi:10.1016/j.agsy.2021.103085.
Hubau, W., et al., 2020: Asynchronous carbon sink saturation in African and Amazonian tropical forests. Nature, 579 (7797), 80–87, doi:10.1038/s41586-020-2035-0.
Hules, M. and S.J. Singh, 2017: India’s land grab deals in Ethiopia: Food security or global politics?Land Use Policy, 60, 343–351, doi:10.1016/j.landusepol.2016.10.035.
Humphries, S., et al., 2015: Synergies at the interface of farmer–scientist partnerships: agricultural innovation through participatory research and plant breeding in Honduras. Agric. Food Secur. , 4 (1), 1–17, doi:10.1186/s40066-015-0046-0.
Hunsberger, C., et al., 2017: Climate change mitigation, land grabbing and conflict: towards a landscape-based and collaborative action research agenda. Can. J. Dev. Stud. , 38 (3), 305–324, doi:10.1080/02255189.2016.1250617.
Huntington, H.P., et al., 2015: Vessels, risks, and rules: planning for safe shipping in Bering Strait. Mar. Policy. , 51, 119–127, doi:10.1016/j.marpol.2014.07.027.
Huntington, H.P., L.T. Quakenbush and M. Nelson, 2017: Evaluating the effects of climate change on indigenous marine mammal hunting in northern and western Alaska using traditional knowledge. Front. Mar. Sci. , 4, 319, doi:10.3389/fmars.2017.00319.
Hupp, J., M. Brubaker, K. Wilkinson and J. Williamson, 2015: How are your berries? Perspectives of Alaska’s environmental managers on trends in wild berry abundance. Int. J. Circumpolar Health, 74, 28704, doi:10.3402/ijch.v74.28704.
Huq, N., A. Bruns, L. Ribbe and S. Huq, 2017: Mainstreaming ecosystem services based climate change adaptation (EbA) in Bangladesh: status, challenges and opportunities. Sustainability, 9 (6), 926, doi:10.3390/su9060926.
Huq, N. and A. Stubbings, 2016: How is the role of ecosystem services considered in local level flood management policies: case study in Cumbria, England. J. Environ. Assess. Policy Manag. , 17 (04), 1550032, doi:10.1142/s1464333215500325.
Hurlbert, M., J. Krishnaswamy, E. Davin, F.X. Johnson, C.F. Mena, J. Morton, S. Myeong, D. Viner, K. Warner, A. Wreford, S. Zakieldeen, Z. Zommers, 2019: Risk management and decision-making in relation to sustainable development. In: Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems[Shukla, P. R., J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H. O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi and J. Malley (eds.)]. In press.
Hussain, A., G. Rasul, B. Mahapatra and S. Tuladhar, 2016: Household food security in the face of climate change in the Hindu-Kush Himalayan region. Food Secur, 8 (5), 921–937, doi:10.1007/s12571-016-0607-5.
Hussain, M.I., M. Farooq, A. Muscolo and A. Rehman, 2020: Crop diversification and saline water irrigation as potential strategies to save freshwater resources and reclamation of marginal soils-a review. Environ. Sci. Pollut. Res. Int. , 27 (23), 28695–28729, doi:10.1007/s11356-020-09111-6.
Hutter, S.E., et al., 2018: Assessing changing weather and the El Nino Southern Oscillation impacts on cattle rabies outbreaks and mortality in Costa Rica (1985-2016). BMC Vet Res, 14 (1), 285, doi:10.1186/s12917-018-1588-8.
Huyer, S., et al., 2019: Learning and action for gender-transformative climate-smart agriculture. Wageningen, the Netherlands, 26–26. CCAFS Working Paper No. 279. CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS).
Huyer, S. and S. Partey, 2020: Weathering the storm or storming the norms? Moving gender equality forward in climate-resilient agriculture Introduction to the Special Issue on Gender Equality in Climate-Smart Agriculture: Approaches and Opportunities. Clim Change, 158 (1), 1–12, doi:10.1007/s10584-019-02612-5.
Hvas, M. and F. Oppedal, 2019: Physiological responses of farmed Atlantic salmon and two cohabitant species of cleaner fish to progressive hypoxia. Aquaculture, 512, 734353, doi:10.1016/j.aquaculture.2019.734353.
Ickowitz, A., et al., 2019: Agricultural intensification, dietary diversity, and markets in the global food security narrative. Glob. Food Secur. Policy Econ. Environ. , 20, 9–16, doi:10.1016/j.gfs.2018.11.002.
IFAD, 2019: Creating opportunities for rural youth. 2019 Rural Development Report . International Fund for Agricultural Development, https://www.ifad.org/documents/38714170/41133075/RDR_report.pdf/7282db66-2d67-b514-d004-5ec25d9729a0.
IGAD, 2020: Food Security and Nutrition Response Strategy. Intergovernmental Authority on Development, https://www.icpac.net/fsnwg/igad-food-security-and-nutrition-response-strategy/ (accessed January 3, 2022) . (62 pp).
Iiyama, M., et al., 2017: Understanding patterns of tree adoption on farms in semi-arid and sub-humid Ethiopia. Agrofor. Syst. , 91 (2), 271–293, doi:10.1007/s10457-016-9926-y.
Iiyama, M., et al., 2018: Addressing the paradox - the divergence between smallholders’ preference and actual adoption of agricultural innovations. Int. J. Agric. Sustain. , 16 (6), 472–485, doi:10.1080/14735903.2018.1539384.
Iizumi, T., 2019: Emerging adaptation to climate change in agriculture. In: Adaptation to Climate Change in Agriculture: Research and Practices[Iizumi, T., R. Hirata and R. Matsuda(eds.)]. Springer Nature, Singapore, Singapore, pp. 3–16. ISBN 978-9811392344.
Iizumi, T., et al., 2021a: Rising temperatures and increasing demand challenge wheat supply in Sudan. Nat. Food, 2 (1), 19–27, doi:10.1038/s43016-020-00214-4.
Iizumi, T., et al., 2017: Responses of crop yield growth to global temperature and socioeconomic changes. Sci Rep, 7 (1), 7800, doi:10.1038/s41598-017-08214-4.
Iizumi, T. and N. Ramankutty, 2016: Changes in yield variability of major crops for 1981–2010 explained by climate change. Environ. Res. Lett. , 11 (3), 34003, doi:10.1088/1748-9326/11/3/034003.
Iizumi, T., et al., 2020: Climate change adaptation cost and residual damage to global crop production. Clim. Res. , 80 (3), 203–218, doi:10.3354/cr01605.
Iizumi, T., et al., 2018: Crop production losses associated with anthropogenic climate change for 1981–2010 compared with preindustrial levels. Int. J. Climatol. , 38 (14), 5405–5417, doi:10.1002/joc.5818.
Iizumi, T., et al., 2021b: Global within-season yield anomaly prediction for major crops derived using seasonal forecasts of large-scale climate indices and regional temperature and precipitation. Weather. Forecast. , 36 (1), 285–299, doi:10.1175/waf-d-20-0097.1.
Iizumi, T. and R. Wagai, 2019: Leveraging drought risk reduction for sustainable food, soil and climate via soil organic carbon sequestration. Sci Rep, 9, 19744, doi:10.1038/s41598-019-55835-y.
Ilboudo Nébié, E.K., D. Ba and A. Giannini, 2021: Food security and climate shocks in Senegal: Who and where are the most vulnerable households?Glob. Food Secur. , 29, 100513–100513, doi:10.1016/j.gfs.2021.100513.
Im, E.S., J.S. Pal and E.A.B. Eltahir, 2017: Deadly heat waves projected in the densely populated agricultural regions of South Asia. Sci. Adv. , 3 (8), e1603322, doi:10.1126/sciadv.1603322.
Imbach, P., et al., 2017: Coupling of pollination services and coffee suitability under climate change. Proc. Natl. Acad. Sci. USA, 114 (39), 10438–10442, doi:10.1073/pnas.1617940114.
Imbong, J.D., 2021: “Bungkalan” and the Manobo-Pulangihon tribe’s resistance to corporate land-grab in Bukidnon, Mindanao. AlterNative, 17 (1), 23–31, doi:10.1177/1177180120967724.
Inaotombi, S. and P.C. Mahanta, 2019: Pathways of socio-ecological resilience to climate change for fisheries through indigenous knowledge. Hum. Ecol. Risk Assess. , 25 (8), 2032–2044, doi:10.1080/10807039.2018.1482197.
Ingalls, M.L., et al., 2018: The transboundary displacement of deforestation under REDD+: Problematic intersections between the trade of forest-risk commodities and land grabbing in the Mekong region. Glob. Environ. Chang. , 50 (November 2017), 255–267, doi:10.1016/j.gloenvcha.2018.04.003.
Ingram, D.J., 2020: Wild meat in changing times. J. Ethnobiol, 40 (2), 117–130, doi:10.2993/0278-0771-40.2.117.
Ioannou, L.G., et al., 2017: Time-motion analysis as a novel approach for evaluating the impact of environmental heat exposure on labor loss in agriculture workers. Temperature, 4 (3), 330–340, doi:10.1080/23328940.2017.1338210.
IPBES, 2019: Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services[Díaz, S., J. Settele, E. S. Brondízio E.S., H. T. Ngo, M. Guèze, J. Agard, A. Arneth, P. Balvanera, K. A. Brauman, S. H. M. Butchart, K. M. A. Chan, L. A. Garibaldi, K. Ichii, J. Liu, S. M. Subramanian, G. F. Midgley, P. Miloslavich, Z. Molnár, D. Obura, A. Pfaff, S. Polasky, A. Purvis, J. Razzaque, B. Reyers, R. Roy Chowdhury, Y. J. Shin, I. J. Visseren-Hamakers, K. J. Willis and C. N. Zayas (eds.)]. IPBES secretariat, Bonn, Germany.
IPC Global Partners, 2019: Integrated Food Security Phase Classification Technical Manual. Version 3.0, Evidence and Standards for Better Food Security and Nutrition Decisions., FAO, Rome. Available at: https://www.ipcinfo.org/fileadmin/user_upload/ipcinfo/docs/IPC_Technical_Manual_3_Final.pdf (accessed January 28, 2022).
IPCC, 2018: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty[Masson-Delmotte, V., P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P.R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J.B.R. Matthews, Y. Chen, X. Zhou, M.I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, and T. Waterfield (eds.)]. In Press.
IPCC, 2019a: Annex I: Glossary [van Diemen, R. (ed.)]. In: Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems[Shukla, P. R., J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H. O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi and J. Malley (eds.)], pp. In press.
IPCC, 2019b: Climate Change and Land: An IPCC Special Report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems[P.R. Shukla, J. S., E. Calvo Buendia, V. Masson-Delmotte, H.- O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi, J. Malley (ed.)]. In press.
IPCC, 2019c: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate[H.-O. Pörtner, D.C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegría, M. Nicolai, A. Okem, J. Petzold, B. Rama, N.M. Weyer (eds.)]. In press.
IPCC, 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. [Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press, Cambridge, UK.
IPPC Secretariat, 2021: Scientific review of the impact of climate change on plant pests- A global challenge to prevent and mitigate plant pest risks in agriculture, forestry and ecosystems. FAO on behalf of the IPPC Secretariat, Rome, Italy. ISBN 978-92-5-134435-4.
Irimia, L.M., C.V. Patriche and O.C. Murariu, 2018: The impact of climate change on viticultural potential and wine grape varieties of a temperate wine growing region. Appl. Ecol. Environ. Res. , 16 (3), 2663–2680, doi:10.15666/aeer/1603_26632680.
Isakson, S.R., 2014: Food and finance: the financial transformation of agro-food supply chains. J Peasant Stud, 41 (5), 749–775, doi:10.1080/03066150.2013.874340.
Isakson, S.R., 2015: Derivatives for development? Small-farmer vulnerability and the financialization of climate risk management. J. Agrar. Chang. , 15 (4), 569–580, doi:10.1111/joac.12124.
Isbell, F., et al., 2017: Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. , 105 (4), 871–879, doi:10.1111/1365-2745.12789.
Ishigooka, Y., et al., 2021: Revision of estimates of climate change impacts on rice yield and quality in Japan by considering the combined effects of temperature and CO2 concentration. J. Agric. Meteorol, 77 (2), 139–149, doi:10.2480/agrmet.D-20-00038.
Islam, M.M., et al., 2019: Vulnerability of inland and coastal aquaculture to climate change: evidence from a developing country. Aquac. Fish. , 4 (5), 183–189, doi:10.1016/j.aaf.2019.02.007.
Istomin, K.V. and J.O. Habeck, 2016: Permafrost and indigenous land use in the northern Urals: Komi and Nenets reindeer husbandry. Polar Sci. , 10 (3), 278–287, doi:10.1016/j.polar.2016.07.002.
Iticha, B. and A. Husen, 2019: Adaptation to climate change using indigenous weather forecasting systems in Borana pastoralists of southern Ethiopia. Clim. Dev. , 11 (7), 564–573, doi:10.1080/17565529.2018.1507896.
Ito, A., et al., 2018: Comparative phenology of dormant Japanese pear (Pyrus pyrifolia) flower buds: a possible cause of ‘flowering disorder’. Tree Physiol, 38 (6), 825–839, doi:10.1093/treephys/tpx169.
Iversen, M., et al., 2014: Detection of transgenes in local maize varieties of small-scale farmers in eastern cape, South Africa. Plos One, 9 (12), e116147, doi:10.1371/journal.pone.0116147.
Iverson, L.R., A.M. Prasad, M.P. Peters and S.N. Matthews, 2019: Facilitating adaptive forest management under climate change: a spatially specific synthesis of 125 species for habitat changes and assisted migration over the eastern United States. Forests, 10 (11), 989, doi:10.3390/f10110989.
Jachnik, R., R.M. Mirabile and A. Dobrinevski, 2019: Tracking Finance Flows Towards Assessing their Consistency with Climate Objectives. OECD Environment Working Papers, Organization for Economic Co-operation and Development, Paris, France, https://doi.org/10.1787/82cc3a4c-en (accessed 05/08/2021) . (42 pp).
Jacka, J.K., 2016: Correlating local knowledge with climatic data: Porgeran experiences of climate change in Papua New Guinea. In: Anthropology and Climate Change: From Actions to Transformations[Crate, S.A. and M. Nuttall(eds.)], 2nd edn. Routledge, New York, NY, USA and Oxon, UK, pp. 186–199. ISBN 978-1629580005.
Jackley, J., L. Gardner, A. F. Djunaedi and A.K. Salomon, 2016: Ancient clam gardens, traditional management portfolios, and the resilience of coupled human-ocean systems. Ecol. Soc. , 21 (4).
Jacobi, J., S. Rist and M. A. Altieri, 2017: Incentives and disincentives for diversified agroforestry systems from different actors’ perspectives in Bolivia. Int. J. Agric. Sustain. , 15 (4), 365–379, doi:10.1080/14735903.2017.1332140.
Jacobson, C., S. Crevello, C. Chea and B. Jarihani, 2019: When is migration a maladaptive response to climate change?Reg Environ Change, 19 (1), 101–112, doi:10.1007/s10113-018-1387-6.
Jactel, H., et al., 2018: Positive biodiversity-productivity relationships in forests: climate matters. Biol. Lett. , 14 (4), 20170747, doi:10.1098/rsbl.2017.0747.
Jaeger, C., et al., 2019: Mass balanced based LCA of a common carp-lettuce aquaponics system. Aquac Eng, 84, 29–41, doi:10.1016/j.aquaeng.2018.11.003.
Jagermeyr, J. and K. Frieler, 2018: Spatial variations in crop growing seasons pivotal to reproduce global fluctuations in maize and wheat yields. Sci. Adv. , 4 (11), eaat4517, doi:10.1126/sciadv.aat4517.
Jägermeyr, J., et al., 2021: Climate change signal in global agriculture emerges earlier in new generation of climate and crop models. Nat. Food, 2 (11), 873-885, doi:10.1038/s43016-021-00400-y.
Jägermeyr, J., A. Pastor, H. Biemans and D. Gerten, 2017: Reconciling irrigated food production with environmental flows for Sustainable Development Goals implementation. Nat Commun, 8, 15900, doi:10.1038/ncomms15900.
Jain, N. and R.S. Nadgauda, 2013: Commiphora wight (Arnott) Bhandari—a natural source of guggulsterone: facing a high risk of extinction in its natural habitat. Am J Plant Sci, 4 (6), 57–68, doi:10.4236/ajps.2013.46A009.
Jalloh, A., et al., 2013: West African agriculture and climate change: A comprehensive analysis. IFPRI Issue Brief, International Food Policy Research Institute, Washington, DC, USA, http://dx.doi.org/10.2499/9780896292048. Accessed 2021 . (408 pp).
Jamero, M.L., M. Onuki, M. Esteban and N. Tan, 2018: Community-based adaptation in low-lying islands in the Philippines: challenges and lessons learned. Reg Environ Change, 18 (8), 2249–2260, doi:10.1007/s10113-018-1332-8.
James, S.J. and C. James, 2010: The food cold-chain and climate change. Food. Res. Int. , 43 (7), 1944–1956, doi:10.1016/j.foodres.2010.02.001.
Jane, S.F., et al., 2021: Widespread deoxygenation of temperate lakes. Nature, 594 (7861), 66–70, doi:10.1038/s41586-021-03550-y.
Janetos, A., et al., 2017: The Risks of Multiple Breadbasket Failures in the 21st Century: A Science Research Agenda. In: Pardee Center Research Report [Pardee Center for the Study of the Longer-Range Future(ed.)]. Boston University, Boston, MA, USA. ISBN 978-1936727148 22 pp).
Janhiainen, J.S. and L. Hooli, 2017: Indigenous knowledge and developing countries’ innovation systems: the case of Namibia. Int. J. Innov. Stud. , 1, 89–106, doi:10.3724/SP.J.1440.101007.
Jans, Y., W. von Bloh, S. Schaphoff and C. Muller, 2021: Global cotton production under climate change - Implications for yield and water consumption. Hydrol. Earth Syst. Sci. , 25 (4), 2027–2044, doi:10.5194/hess-25-2027-2021.
Janssens, C., et al., 2020: Global hunger and climate change adaptation through international trade. Nat. Clim. Chang. , 10, 829–835, doi:10.1038/s41558-020-0847-4.
Jantarasami, L., et al., 2018: Chapter 15 : Tribal and Indigenous Communities. Impacts, Risks, and Adaptation in the United States: The Fourth National Climate Assessment, Volume II. Washington, DC, https://nca2018.globalchange.gov/chapter/15/.
Jarecki, M., et al., 2018: Long-term Trends in Corn Yields and Soil Carbon under Diversified Crop Rotations. J. Environ. Qual. , 47 (4), 635–643, doi:10.2134/jeq2017.08.0317.
Jayanthi, M., et al., 2020: Assessment of land and water ecosystems capability to support aquaculture expansion in climate-vulnerable regions using analytical hierarchy process based geospatial analysis. J. Environ. Manag. , 270, 110952, doi:10.1016/j.jenvman.2020.110952.
Jeliazkov, A., et al., 2016: Impacts of agricultural intensification on bird communities: new insights from a multi-level and multi-facet approach of biodiversity. Agric. Ecosyst. Environ. , 216, 9–22, doi:10.1016/j.agee.2015.09.017.
Jennings, S.A., et al., 2020: Global Potato Yields Increase Under Climate Change With Adaptation and CO2Fertilisation. Front. Sustain. Food Syst. , 4, 248.
Jensen, N. and C. Barrett, 2017: Agricultural index insurance for development. Appl. Econ. Perspect. Policy. , 39 (2), 199–219, doi:10.1093/aepp/ppw022.
Jeon, J.-Y., P. Adhikari and C. Seo, 2020: Impact of climate change on potential dispersal of Paeonia obovata (paeoniaceae), a critically endangered medicinal plant of South Korea. Eco Env Cons, 26, S145–S155.
Jha, S., et al., 2014: Shade coffee: update on a disappearing refuge for biodiversity. BioScience, 64 (5), 416–428, doi:10.1093/biosci/biu038.
Jiménez-Quiroz, M.C., et al., 2019: Impact of “The Blob” and “El Niño” in the SW Baja California Peninsula: Plankton and Environmental Variability of Bahia Magdalena. Front. Mar. Sci, 6, 25.
Jin, J., R. Armstrong and C. Tang, 2019: Impact of elevated CO2 on grain nutrient concentration varies with crops and soils - A long-term FACE study. Sci. Total Environ. , 651 (Pt 2), 2641–2647, doi:10.1016/j.scitotenv.2018.10.170.
Jin, Z., E.A. Ainsworth, A.D.B. Leakey and D.B. Lobell, 2018: Increasing drought and diminishing benefits of elevated carbon dioxide for soybean yields across the US Midwest. Glob Chang Biol, 24 (2), e522–e533, doi:10.1111/gcb.13946.
Jiri, O., P.L. Mafongoya and P. Chivenge, 2017: Building climate change resilience through adaptation in smallholder farming systems in semi-arid Zimbabwe. Int. J. Clim. Chang. Strateg. Manag. , 9 (2), 151–165, doi:10.1108/IJCCSM-07-2016-0092.
Jobin Poirier, E., G. Pickering and R. Plummer, 2019: Doom, gloom, or boom? Perceptions of climate change among Canadian winegrowers. Int. J. Wine Res. , 11, 1–11, doi:10.2147/ijwr.S188787.
Joffre, O.M., L. Klerkx, M. Dickson and M. Verdegem, 2017: How is innovation in aquaculture conceptualized and managed? A systematic literature review and reflection framework to inform analysis and action. Aquaculture, 470, 129–148, doi:10.1016/j.aquaculture.2016.12.020.
Johansson, E., S. Olin and J. Seaquist, 2020: Foreign demand for agricultural commodities drives virtual carbon exports from Cambodia. Environ. Res. Lett. , 15 (6), doi:10.1088/1748-9326/ab8157.
Johansson, E.L. and E. Isgren, 2017: Local perceptions of land-use change: using participatory art to reveal direct and indirect socioenvironmental effects of land acquisitions in Kilombero Valley, Tanzania. Ecol. Soc. , 22 (1), doi:10.5751/Es-08986-220103.
Johnson, A., K. Koranteng, S. Petersen and M. Tandstad, 2014: Establishing a Baseline for the Implementation of an Ecosystem Approach to Marine Fisheries Management in West and Central Africa. FAO-EAF-Nansen Project Report, Food and Agriculture Organization of the United Nations, Rome, Italy, http://www.fao.org/fishery/docs/DOCUMENT/eaf_nansen/Reports/EAF-NansenReportNo20_en.pdf. Accessed 2021 . (44 pp).
Johnson, J.E., et al., 2020: Impacts of climate change on marine resources in the Pacific Island region. In: Climate Change and Impacts in the Pacific[Kumar, L.(ed.)]. Springer Nature, Cham, Switzerland, pp. 359–402. ISBN 978-3030328771.
Johnson, J.S., E.D. Nobmann, E. Asay and A.P. Lanier, 2009: Dietary intake of Alaska Native people in two regions and implications for health: the Alaska Native dietary and subsistence food assessment project. Int. J. Circumpolar Health, 68 (2), 109–122, doi:10.3402/ijch.v68i2.18320.
Johnson, L., B. Wandera, N. Jensen and R. Banerjee, 2019: Competing expectations in an index-based livestock insurance project. J. Dev. Stud. , 55 (6), 1221–1239, doi:10.1080/00220388.2018.1453603.
Jones, A.D., 2017a: Critical review of the emerging research evidence on agricultural biodiversity, diet diversity, and nutritional status in low- and middle-income countries. Nutr. Rev. , 75 (10), 769–782, doi:10.1093/nutrit/nux040.
Jones, A.D., 2017b: On-farm crop species richness is associated with household diet diversity and quality in subsistence- and market-oriented farming households in Malawi. J. Nutr. , 147 (1), 86–96, doi:10.3945/jn.116.235879.
Jones, A.D., et al., 2018: Farm-level agricultural biodiversity in the Peruvian Andes is associated with greater odds of women achieving a minimally diverse and micronutrient adequate diet. J. Nutr. , 148 (10), 1625–1637, doi:10.1093/jn/nxy166.
Jones, A.E., et al., 2019: Bluetongue risk under future climates. Nat. Clim. Chang. , 9 (2), 153–157, doi:10.1038/s41558-018-0376-6.
Jones, M.B., 2019: Projected climate change and the global distribution of grasslands. In: Grasslands and Climate Change[Gibson, D. and J. Newman(eds.)]. Cambridge University Press, Cambridge, UK, pp. 67–81. ISBN 978-1107195264.
Jones, R.A.C. and M. Barbetti, 2012: Influence of climate change on plant disease infections and epidemics caused by viruses and bacteria. Cab Rev. Perspect. Agric. Veterinary Sci. Nutr. Nat. Resour. , 7 (022), doi:10.1079/pavsnnr20127022.
Jönsson, A.M., F. Lagergren and B. Smith, 2015: Forest management facing climate change – an ecosystem model analysis of adaptation strategies. Mitig. Adapt. Strateg. Glob. Change, 20 (2), 201–220, doi:10.1007/s11027-013-9487-6.
Joris, I., J. Bronders, B. van der Grift and P. Seuntjens, 2014: Model-based scenario analysis of the impact of remediation measures on metal leaching from soils contaminated by historic smelter emissions. J. Environ. Qual. , 43 (3), 859–868, doi:10.2134/jeq2013.07.0287.
Jost, C., et al., 2016: Understanding gender dimensions of agriculture and climate change in smallholder farming communities. Clim. Dev. , 8 (2), 133–144, doi:10.1080/17565529.2015.1050978.
Ju, R.T., et al., 2020: Emerging risks of non-native species escapes from aquaculture: call for policy improvements in China and other developing countries. J Appl Ecol, 57 (1), 85–90, doi:10.1111/1365-2664.13521.
Juhola, S., E. Glaas, B.O. Linner and T.S. Neset, 2016: Redefining maladaptation. Environ Sci Policy, 55, 135–140, doi:10.1016/j.envsci.2015.09.014.
Jurjonas, M. and E. Seekamp, 2019: Balancing carbon dioxide: a case study of forest preservation, out-migration, and afforestation in the Pueblos Mancomunados of Oaxaca, Mexico. J. Sustain. For. , 38 (7), 697–714, doi:10.1080/10549811.2019.1602058.
Kagunyu, A.W. and J. Wanjohi, 2014: Camel rearing replacing cattle production among the Borana community in Isiolo County of Northern Kenya, as climate variability bites. Pastoralism, 4 (1), 13, doi:10.1186/s13570-014-0013-6.
Kais, S.M. and M.S. Islam, 2018: Impacts of and resilience to climate change at the bottom of the shrimp commodity chain in Bangladesh: a preliminary investigation. Aquaculture, 493, 406–415, doi:10.1016/j.aquaculture.2017.05.024.
Kais, S.M. and M.S. Islam, 2021: Resilience to Climate Change in Industrial Shrimping in Bangladesh: Assessing the Comparative Role of the Stakeholders. Sustainability, 13 (1), 307.
Kaky, E. and F. Gilbert, 2017: Predicting the distributions of Egypt’s medicinal plants and their potential shifts under future climate change. PLoS ONE, 12 (11), e187714, doi:10.1371/journal.pone.0187714.
Kalikoski, D.C., et al., 2018: Understanding the impacts of climate change for fisheries and aquaculture: applying a poverty lens. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 19–39. ISBN 978-9251306079.
Kaminski, A.M., S. Genschick, A.S. Kefi and F. Kruijssen, 2018: Commercialization and upgrading in the aquaculture value chain in Zambia. Aquaculture, 493, 355–364, doi:10.1016/j.aquaculture.2017.12.010.
Kane, D.A., et al., 2021: Soil organic matter protects US maize yields and lowers crop insurance payouts under drought. Environ. Res. Lett. , 16 (4), 44018, doi:10.1088/1748-9326/abe492.
Kangmennaang, J., et al., 2017: Impact of a participatory agroecological development project on household wealth and food security in Malawi. Food Secur, 9 (3), 561–576, doi:10.1007/s12571-017-0669-z.
Kansanga, M.M., et al., 2021: Does participatory farmer-to-farmer training improve the adoption of sustainable land management practices?Land Use Policy, 108, 105477–105477, doi:10.1016/j.landusepol.2021.105477.
Kanter, D.R., et al., 2018: Evaluating agricultural trade-offs in the age of sustainable development. Agric Syst , 163, 73–88, doi:10.1016/j.agsy.2016.09.010.
Kao, Y.C., et al., 2020: Effects of climate and land-use changes on fish catches across lakes at a global scale. Nat Commun, 11 (1), 2526, doi:10.1038/s41467-020-14624-2.
Karavani, A., et al., 2018: Effect of climatic and soil moisture conditions on mushroom productivity and related ecosystem services in Mediterranean pine stands facing climate change. Agric For Meteorol, 248, 432–440, doi:10.1016/j.agrformet.2017.10.024.
Kareem, B., et al., 2020: Pathways for resilience to climate change in African cities. Environ. Res. Lett. , 15 (7), 73002, doi:10.1088/1748-9326/ab7951.
Karg, H., et al., 2016: Foodsheds and city region food systems in two West African cities. Sustainability, 8 (12), 1175, doi:10.3390/su8121175.
Karlsson, L., L.O. Naess, A. Nightingale and J. Thompson, 2018: “Triple wins” or “triple faults”? Analysing the equity implications of policy discourses on climate-smart agriculture (CSA). J Peasant Stud, 45 (1), 150–174, doi:10.1080/03066150.2017.1351433.
Karttunen, K., J. Wolf, C. Garcia and A. Meybeck, 2017: Addressing Agriculture, Forestry and Fisheries in National Adaptation Plans [Supplementary Guidelines. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN 978-9251097557. 104 pp.
Kassi, J.B., et al., 2018: Remotely sensing the biophysical drivers of Sardinella aurita wariability in Ivorian waters. Remote Sens, 10 (5), 785, doi:10.3390/rs10050785.
Kathiresan, R. and G. Gualbert, 2016: Impact of climate change on the invasive traits of weeds. Weed Biol. Manag. , 16 (2), 59–66, doi:10.1111/wbm.12096.
Kaushal, N. and F.M. Muchomba, 2015: How consumer price subsidies affect nutrition. World Dev, 74, 25–42, doi:10.1016/j.worlddev.2015.04.006.
Kawall, K., 2019: New possibilities on the horizon: genome editing makes the whole genome accessible for changes. Front. Plant Sci. , 10, 525, doi:10.3389/fpls.2019.00525.
Keairns, D.L., R.C. Darton and A. Irabien, 2016: The energy-water-food nexus. Annu. Rev. Chem. Biomol. Eng. , 7, 239–262, doi:10.1146/annurev-chembioeng-080615-033539.
Keatinge, J.D.H., et al., 2016: Assessing air temperature trends in Mesoamerica and their implications for the future of horticulture. Eur. J. Hortic. Sci. , 81 (2), 63–77, doi:10.17660/eJHS.2016/81.2.1.
Keatinge, J.D.H., et al., 2015: Indigenous vegetables worldwide: their importance and future development. Acta Hortic. , 1102, 1–20, doi:10.17660/ActaHortic.2015.1102.1.
Keenan, R.J., 2015: Climate change impacts and adaptation in forest management: a review. Ann. For. Sci. , 72 (2), 145–167, doi:10.1007/s13595-014-0446-5.
Kehoe, L., et al., 2017: Biodiversity at risk under future cropland expansion and intensification. Nat Ecol Evol, 1 (8), 1129–1135, doi:10.1038/s41559-017-0234-3.
Kellett, J. and K. Peters, 2014: Dare to Prepare: Taking Risk Seriously. Financing Emergency Preparedness: From Fighting Crisis to Managing Risk. Overseas Development Institute, London, UK, <Go to WoS>://WOS:000412260400023. Accessed 2021 . (133 pp).
Kelley, C., et al., 2017: Commentary on the Syria case: climate as a contributing factor. Polit. Geogr. , 60, 245–247, doi:10.1016/j.polgeo.2017.06.013.
Kellogg, J., et al., 2010: Alaskan wild berry resources and human health under the cloud of climate change. J. Agric. Food Chem. , 58 (7), 3884–3900, doi:10.1021/jf902693r.
Kenny, T.A., et al., 2018: Caribou (Rangifer tarandus) and Inuit nutrition security in Canada. EcoHealth, 15 (3), 590–607, doi:10.1007/s10393-018-1348-z.
Kent, R. and R. Hannay, 2020: Explaining “carbon” in community sequestration projects: a key element in the creation of local carbon knowledges. Environ. Commun. , 14 (3), 364–377, doi:10.1080/17524032.2019.1673459.
Keppen, D. and T. Dutcher, 2015: The 2014 drought and water management policy impacts on California’s Central Valley food production. J Environ Stud Sci, 5 (3), 362–377, doi:10.1007/s13412-015-0283-3.
Kerr, A., et al., 2018: Vulnerability of California specialty crops to projected mid-century temperature changes. Clim. Change, 148 (3), 419–436, doi:10.1007/s10584-017-2011-3.
Kerr, J.T., et al., 2015: Climate change impacts on bumblebees converge across continents. Science, 349 (6244), 177–180, doi:10.1126/science.aaa7031.
Khalili Tilami, S. and S. Sampels, 2018: Nutritional value of fish: lipids, proteins, vitamins, and minerals. Rev. Fish. Sci. Aquac. , 26 (2), 243–253, doi:10.1080/23308249.2017.1399104.
Khatri-Chhetri, A., P.P. Regmi, N. Chanana and P.K. Aggarwal, 2020: Potential of climate-smart agriculture in reducing women farmers’ drudgery in high climatic risk areas. Clim Change, 158 (1), 29–42, doi:10.1007/s10584-018-2350-8.
Kibblewhite, M.G., K. Ritz and M.J. Swift, 2008: Soil health in agricultural systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. , 363 (1492), 685–701, doi:10.1098/rstb.2007.2178.
Kibet, S., M. Nyangito, L. MacOpiyo and D. Kenfack, 2016: Tracing innovation pathways in the management of natural and social capital on Laikipia Maasai Group Ranches, Kenya. Pastoralism, 6, doi:10.1186/s13570-016-0063-z.
Kibria, G., A.K. Haroon and D. Nugegoda, 2017: Climate change impacts on tropical and temperate fisheries, aquaculture, and seafood security and implications - A review. Livest. Res. Rural. Dev. , 29 (1), 1-29.
Kifani, S., et al., 2018: Climate change impacts, vulnerabilities and adaptations: Eastern Central Atlantic marine fisheries. In: Impacts of Climate Change on Fisheries and Aquaculture: synthesis of Current Knowledge, Adaptation and Mitigation Options[Barange, M., et al.(ed.)]. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 159–184. ISBN 978-9251306079.
Kihila, J.M., 2018: Indigenous coping and adaptation strategies to climate change of local communities in Tanzania: a review. Clim. Dev. , 10 (5), 406–416, doi:10.1080/17565529.2017.1318739.
Kim, B.T., C.L. Brown and D.H. Kim, 2019a: Assessment on the vulnerability of Korean aquaculture to climate change. Mar. Policy. , 99, 111–122, doi:10.1016/j.marpol.2018.10.009.
Kim, W., T. Iizumi and M. Nishimori, 2019b: Global patterns of crop production losses associated with droughts from 1983 to 2009. J. Appl. Meteorol. Clim. , 58 (6), 1233–1244, doi:10.1175/Jamc-D-18-0174.1.
Kimathi, E., et al., 2020: Prediction of breeding regions for the desert locust Schistocerca gregaria in East Africa. Sci Rep, 10 (1), 11937, doi:10.1038/s41598-020-68895-2.
Kimball, B.A., 2016: Crop responses to elevated CO2 and interactions with H2O, N, and temperature. Curr. Opin. Plant Biol. , 31, 36–43, doi:10.1016/j.pbi.2016.03.006.
Kiptot, E., S. Franzel and A. Degrande, 2014: Gender, agroforestry and food security in Africa. Curr. Opin. Environ. Sustain. , 6, 104–109, doi:10.1016/j.cosust.2013.10.019.
Kirchherr, J., D. Reike and M. Hekkert, 2017: Conceptualizing the circular economy: An analysis of 114 definitions. Resour. Conserv. Recycl. , 127, 221–232, doi:10.1016/j.resconrec.2017.09.005.
Kirchner, M., et al., 2015: Ecosystem services and economic development in Austrian agricultural landscapes - the impact of policy and climate change scenarios on trade-offs and synergies. Ecol. Econ. , 109, 161–174, doi:10.1016/j.ecolecon.2014.11.005.
Kissinger, G., M. Brockhaus and S.R. Bush, 2021: Policy integration as a means to address policy fragmentation: Assessing the role of Vietnam’s national REDD+ action plan in the central highlands. Environ Sci Policy, 119, 85–92, doi:10.1016/j.envsci.2021.02.011.
Kitalyi, A., et al., 2005: Why keep livestock if you are poor. In: Livestock and Wealth Creation. Improving the Husbandry of Animals Kept by Resource-Poor People in Developing Countries[Owen, E., A. Kitalyi, N. Jayasuriya and T. Smith(eds.)]. Nottingham University Press, Nottingham, UK, pp. 13–27. ISBN 978-1904761327.
Kizildeniz, T., J.J. Irigoyen, I. Pascual and F. Morales, 2018: Simulating the impact of climate change (elevated CO2 and temperature, and water deficit) on the growth of red and white Tempranillo grapevine in three consecutive growing seasons (2013-2015). Agric. Water Manag. , 202, 220–230, doi:10.1016/j.agwat.2018.02.006.
Kizildeniz, T., et al., 2015: Effects of climate change including elevated CO2 concentration, temperature and water deficit on growth, water status, and yield quality of grapevine (Vitis vinifera L.) cultivars. Agric. Water Manag. , 159, 155–164, doi:10.1016/j.agwat.2015.06.015.
Kjellstrom, T., et al., 2016: Heat, human performance, and occupational health: a key issue for the assessment of global climate change impacts. Annu. Rev. Public Health, 37 (1), 97–112, doi:10.1146/annurev-publhealth-032315-021740.
Kjellstrom, T., et al., 2018: Estimating population heat exposure and impacts on working people in conjunction with climate change. Int. J. Biometeorol. , 62 (3), 291–306, doi:10.1007/s00484-017-1407-0.
Kjellström, T., et al., 2019: Working on a Warmer Planet : The Effect of Heat Stress on Labour Productivity and Decent Work, 1st edn., International Labor Organization, Geneva, Switzerland, ISBN 978-9221329671. 97 pp.
Klein, J.A., et al., 2014: Unexpected climate impacts on the Tibetan Plateau: Local and scientific knowledge in findings of delayed summer. Glob. Environ. Change. , 28, 141–152, doi:10.1016/j.gloenvcha.2014.03.007.
Klemm, T., D.D. Briske and M.C. Reeves, 2020: Vulnerability of rangeland beef cattle production to climate-induced NPP fluctuations in the US Great Plains. Glob. Chang. Biol. , 26 (9), 4841–4853, doi:10.1111/gcb.15202.
Klenk, N., A. Fiume, K. Meehan and C. Gibbes, 2017: Local knowledge in climate adaptation research: moving knowledge frameworks from extraction to co-production. Wiley. Interdiscip. Rev. Clim. Change. , 8 (5), e475, doi:10.1002/wcc.475.
Klinger, D.H., S.A. Levin and J.R. Watson, 2017: The growth of finfish in global open-ocean aquaculture under climate change. Proc. Biol. Sci. , 284 (1864), doi:10.1098/rspb.2017.0834.
Knox, J., A. Daccache, T. Hess and D. Haro, 2016: Meta-analysis of climate impacts and uncertainty on crop yields in Europe. Environ. Res. Lett. , 11 (11), 113004, doi:10.1088/1748-9326/11/11/113004.
Kohler, F. and E.S. Brondizio, 2017: Considering the needs of indigenous and local populations in conservation programs. Conserv. Biol. , 31 (2), 245–251, doi:10.1111/cobi.12843.
Kohler, I.H., S.C. Huber, C.J. Bernacchi and I.R. Baxter, 2019: Increased temperatures may safeguard the nutritional quality of crops under future elevated CO2 concentrations. Plant J. , 97 (5), 872–886, doi:10.1111/tpj.14166.
Koirala, A. and K.B. Shrestha, 2017: Effects of climate change on the livestock population in Mustang District, Nepal. Asian J. Agric. Dev. , 14 (1), 37–49, doi:10.22004/ag.econ.259247.
Koks, E.E., et al., 2019: A global multi-hazard risk analysis of road and railway infrastructure assets. Nat Commun, 10 (1), 2677, doi:10.1038/s41467-019-10442-3.
Kole, C., et al., 2015: Application of genomics-assisted breeding for generation of climate resilient crops: progress and prospects. Front. Plant Sci. , 6, 563, doi:10.3389/fpls.2015.00563.
Kolström, M., et al., 2011: Reviewing the science and implementation of climate change adaptation measures in European forestry. Forests, 2 (4), 961–982, doi:10.3390/f2040961.
Konchar, K.M., et al., 2015: Adapting in the shadow of Annapurna: a climate tipping point. J. Ethnobiol, 35 (3), 449–471, doi:10.2993/0278-0771-35.3.449.
Konforti, L., 2018: Ungovernable? Indigenous Campesino Resistance to Land Grabbing in Guatemala’s Polochic Valley. Lat. Am. Perspect. , 1–18, doi:10.1177/0094582X18813565.
Kongsager, R., 2017: Barriers to the adoption of alley cropping as a climate-smart agriculture practice: lessons from maize cultivation among the Maya in southern Belize. Forests, 8 (7), 260, doi:10.3390/f8070260.
Kongsager, R., 2018: Linking climate change adaptation and mitigation: A review with evidence from the land-use sectors. Land, 7 (4), doi:10.3390/land7040158.
Koontz, M.J., et al., 2020: Local forest structure variability increases resilience to wildfire in dry western U.S. coniferous forests. Ecol Lett , 23 (3), 483–494, doi:10.1111/ele.13447.
Kopáček, M., 2021: Land-Use Planning and the Public: Is There an Optimal Degree of Civic Participation?Land, 10 (1), 90–90, doi:10.3390/LAND10010090.
Koren, O., B.E. Bagozzi and T.S. Benson, 2021: Food and water insecurity as causes of social unrest : Evidence from geolocated Twitter data. J Peace Res, doi:10.1177/0022343320975091.
Korres, N.E., et al., 2016: Cultivars to face climate change effects on crops and weeds: a review. Agron. Sustain. Dev. , 36 (1), 12, doi:10.1007/s13593-016-0350-5.
Koshiol, J., et al., 2017: Association of aflatoxin and gallbladder cancer. Gastroenterology, 153 (2), 488–494, doi:10.1053/j.gastro.2017.04.005.
Kosmowski, F., A. Leblois and B. Sultan, 2016: Perceptions of recent rainfall changes in Niger: a comparison between climate-sensitive and non-climate sensitive households. Clim Change, 135, 227–241, doi:10.1007/s10584-015-1562-4.
Koufos, G., et al., 2014: Viticulture-climate relationships in Greece: the impacts of recent climate trends on harvest date variation. Int. J. Clim. , 34 (5), 1445–1459, doi:10.1002/joc.3775.
Koufos, G.C., T. Mavromatis, S. Koundouras and G.V. Jones, 2020: Adaptive capacity of winegrape varieties cultivated in Greece to climate change. Curr. Trends Future Proj. , 1201–1219, doi:10.20870/oeno-one.2020.54.4.3129.
Kramer, B., et al., 2019: Building Resilience Through Climate Risk Insurance: Insights from Agricultural Research for Development . CCAFS Working Paper, CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS), Wageningen, the Netherlands, https://hdl.handle.net/10568/106171. Accessed 2021 . (38 pp).
Krčmářová, J. and L. Jeleček, 2016: Czech traditional agroforestry: historic accounts and current status. Agrofor. Syst. , 91 (6), 1087–1100, doi:10.1007/s10457-016-9985-0.
Kremen, C., 2015: Reframing the land-sparing/land-sharing debate for biodiversity conservation. Ann. NY Acad. Sci. , 1355, 52–76, doi:10.1111/nyas.12845.
Kremen, C. and A.M. Merenlender, 2018: Landscapes that work for biodiversity and people. Science, 362 (6412), eaau6020, doi:10.1126/science.aau6020.
Kremen, C. and A. Miles, 2012: Ecosystem services in biologically diversified versus conventional farming systems: benefits, externalities, and trade-offs. Ecol. Soc. , 17 (4), 40, doi:10.5751/Es-05035-170440.
Kriewald, S., et al., 2019: Hungry cities: how local food self-sufficiency relates to climate change, diets, and urbanisation. Environ. Res. Lett. , 14 (9), 94007, doi:10.1088/1748-9326/ab2d56.
Kryza, M., et al., 2015: Observed changes in SAT and GDD and the climatological suitability of the Poland-Germany-Czech Republic transboundary region for wine grapes cultivation. Theor Appl Climatol, 122 (1-2), 207–218, doi:10.1007/s00704-014-1296-7.
Kueppers, L.M., et al., 2005: Modeled regional climate change and California endemic oak ranges. Proc. Natl. Acad. Sci. U. S. A. , 102 (45), 16281–16286, doi:10.1073/pnas.0501427102.
Kuhnlein, H.V. and D.H. Calloway, 1977: Contemporary Hopi food-intake patterns. Ecol. Food. Nutr. , 6 (3), 159–173, doi:10.1080/03670244.1977.9990495.
Kuhnlein, H.V. and O. Receveur, 2007: Local cultural animal food contributes high levels of nutrients for Arctic Canadian Indigenous adults and children. J. Nutr. , 137 (4), 1110–1114, doi:10.1093/jn/137.4.1110.
Kuhnlein, H.V., R. Soueida and O. Receveur, 1996: Dietary nutrient profiles of Canadian Baffin Island Inuit differ by food source, season, and age. J. Am. Diet. Assoc. , 96 (2), 155–162, doi:10.1016/S0002-8223(96)00045-4.
Kumar, P., et al., 2021: Agricultural sustainability in Indian Himalayan Region: Constraints and potentials. Indian J. Ecol. , 48 (3), 649–662.
Kumar, R., 2016: The perils of productivity: making ‘good farmers’ in Malwa, India. J. Agrar. Chang. , 16 (1), 70–93, doi:10.1111/joac.12084.
Kummu, M., et al., 2021: Climate change risks pushing one-third of global food production outside the safe climatic space. One Earth, doi:10.1016/j.oneear.2021.04.017.
Kurpis, J., M. A. Serrato-Cruz and T.P.F. Arroyo, 2019: Modeling the effects of climate change on the distribution of Tagetes lucida Cav. (Asteraceae). Glob. Ecol. Conserv. , 20, e747, doi:10.1016/j.gecco.2019.e00747.
Kurz, V., 2018: Nudging to reduce meat consumption: Immediate and persistent effects of an intervention at a university restaurant. J Environ Econ Manage, 90, 317–341, doi:10.1016/j.jeem.2018.06.005.
Kurz, W.A., et al., 2008: Mountain pine beetle and forest carbon feedback to climate change. Nature, 452 (7190), 987–990, doi:10.1038/nature06777.
Kuyah, S., et al., 2019: Agroforestry delivers a win-win solution for ecosystem services in sub-Saharan Africa. A meta-analysis. Agron. Sustain. Dev. , 39 (5), 47, doi:10.1007/s13593-019-0589-8.
Kyetere, D., et al., 2019: The Role and Contribution of Plant Breeding and Plant Biotechnology to Sustainable Agriculture in Africa. Afr. Focus, 32 (2), 83–108, doi:10.21825/af.v32i2.15772.
Labbé, J., et al., 2016: Vulnerability to the health effects of climate variability in rural southwestern Uganda. Mitig Adapt Strateg Glob Change, 21 (6), 931–953, doi:10.1007/s11027-015-9635-2.
LaCanne, C.E. and J.G. Lundgren, 2018: Regenerative agriculture: merging farming and natural resource conservation profitably. PeerJ, 6, e4428–e4428, doi:10.7717/peerj.4428.
Lade, S.J., et al., 2020: Human impacts on planetary boundaries amplified by Earth system interactions. Nat. Sustain. , 3 (2), 119–128, doi:10.1038/s41893-019-0454-4.
Läderach, P., et al., 2017: Climate change adaptation of coffee production in space and time. Clim. Change, 141 (1), 47–62, doi:10.1007/s10584-016-1788-9.
Ladio, A.H. and M. Lozada, 2004: Patterns of use and knowledge of wild edible plants in distinct ecological environments: a case study of a Mapuche community from northwestern Patagonia. Biodivers Conserv, 13 (6), 1153–1173, doi:10.1023/B:Bioc.0000018150.79156.50.
Lafond, V., G. Lagarrigues, T. Cordonnier and B. Courbaud, 2013: Uneven-aged management options to promote forest resilience for climate change adaptation: effects of group selection and harvesting intensity. Ann. For. Sci. , 71 (2), 173–186, doi:10.1007/s13595-013-0291-y.
Lahive, F., P. Hadley and A.J. Daymond, 2019: The physiological responses of cacao to the environment and the implications for climate change resilience. A review. Agron. Sustain. Dev. , 39 (1), 5, doi:10.1007/s13593-018-0552-0.
Lahmar, R., et al., 2012: Tailoring conservation agriculture technologies to West Africa semi-arid zones: building on traditional local practices for soil restoration. Field Crop. Res. , 132, 158–167, doi:10.1016/j.fcr.2011.09.013.
Laidler, G.J., et al., 2009: Travelling and hunting in a changing Arctic: assessing Inuit vulnerability to sea ice change in Igloolik, Nunavut. Clim Change, 94 (3), 363–397, doi:10.1007/s10584-008-9512-z.
Laidre, K.L. and M.P. Heide-Jorgensen, 2005: Arctic sea ice trends and narwhal vulnerability. Biol. Conserv. , 121 (4), 509–517, doi:10.1016/j.biocon.2004.06.003.
Lake, I.R. and G.C. Barker, 2018: Climate change, foodborne pathogens and illness in higher-income countries. Curr Environ Health Rep, 5 (1), 187–196, doi:10.1007/s40572-018-0189-9.
Lake, I.R., et al., 2014: The effects of river flooding on dioxin and PCBs in beef. Sci. Total Environ. , 491-492, 184–191, doi:10.1016/j.scitotenv.2014.01.080.
Lallo, C.H.O., et al., 2018: Characterizing heat stress on livestock using the temperature humidity index (THI) prospects for a warmer Caribbean. Reg Environ Change, 18 (8), 2329–2340, doi:10.1007/s10113-018-1359-x.
Lam, S., et al., 2019: Community-based monitoring of Indigenous food security in a changing climate: global trends and future directions. Environ. Res. Lett. , 14 (7), 73002, doi:10.1088/1748-9326/ab13e4.
Lam, V.W., W.W. Cheung, G. Reygondeau and U.R. Sumaila, 2016: Projected change in global fisheries revenues under climate change. Sci Rep, 6, 32607, doi:10.1038/srep32607.
Lam, V.W.Y., et al., 2020: Climate change, tropical fisheries and prospects for sustainable development. Nat. Rev. Earth Environ. , 1 (9), 440–454, doi:10.1038/s43017-020-0071-9.
Lamanna, C., et al., 2016: Evidence-based opportunities for out-scaling climate-smart agriculture in East Africa. CCAFS Working Paper, Copenhagen, Denmark, https://hdl.handle.net/10568/77180. Accessed 2021 . (63 pp).
Lamb, A., et al., 2016: The potential for land sparing to o set greenhouse gas emissions from agriculture. Nat. Clim. Chang. , doi:10.1038/NCLIMATE2910.
Lampkin, N., G. Schwarz and S. Bellon, 2020: Policies for agroecology in Europe, building on experiences in France, Germany and the UK. Landbauforsch J Sustainable Organic Agric Syst , 70 (2), 103–112, doi:10.3220/LBF1611684471000.
Land Matrix, 2021: The Land Matrix. https://landmatrix.org, (last accessed January 3, 2022).
Lassaletta, L., et al., 2019: Future global pig production systems according to the Shared Socioeconomic Pathways. Sci. Total Environ. , 665, 739–751, doi:10.1016/j.scitotenv.2019.02.079.
Laube, W., 2016: Climate Change Adaptation, Education, and Rural Transformation in Northern Ghana. Moving Beyond an Agricultural Focus, 121–145. doi:10.1007/978-3-319-31499-0_8.
Laufkotter, C., J. Zscheischler and T.L. Frolicher, 2020: High-impact marine heatwaves attributable to human-induced global warming. Science, 369 (6511), 1621–1625, doi:10.1126/science.aba0690.
Lauria, V., et al., 2018: Importance of fisheries for food security across three climate change vulnerable deltas. Sci. Total Environ. , 640-641, 1566–1577, doi:10.1016/j.scitotenv.2018.06.011.
Law, W. and J. Salick, 2005: Human-induced dwarfing of Himalayan snow lotus, Saussurea laniceps (Asteraceae). Proc. Natl. Acad. Sci. , 102 (29), 10218–10220, doi:10.1073/pnas.0502931102.
Lawanson, T., O. Orelaja and M. Simire, 2014: Effects of climate change on a peri-urban farming community in Lagos, Nigeria. Afr. J. Sci. Technol. Dev. , 6 (6), 511–517, doi:10.1080/20421338.2014.976990.
Lazarus, E.D., 2014: Land grabbing as a driver of environmental change. Area, 46 (1), 74–82, doi:10.1111/area.12072.
Lazoglou, G., C. Anagnostopoulou and S. Koundouras, 2018: Climate change projections for Greek viticulture as simulated by a regional climate model. Theor Appl Climatol, 133 (1-2), 551–567, doi:10.1007/s00704-017-2198-2.
Le Cornu, E., et al., 2018: Spatial management in small-scale fisheries: A potential approach for climate change adaptation in Pacific Islands. Mar. Policy. , 88, 350–358, doi:10.1016/j.marpol.2017.09.030.
Le Mouël, C., M. De Lattre-Gasquet and O. Mora (eds.), 2018: Land Use and Food Security in 2050: A Narrow Road. Éditions Quae, Versailles, France, ISBN 978-2759228805. 400 pp.
Le Mouël, C. and A. Forslund, 2017: How can we feed the world in 2050? A review of the responses from global scenario studies. Eur. Rev. Agric. Econ. , 44 (4), 541–591, doi:10.1093/erae/jbx006.
Le Tacon, F., et al., 2014: Climatic variations explain annual fluctuations in French Périgord black truffle wholesale markets but do not explain the decrease in black truffle production over the last 48 years. Mycorrhiza, 24 (1), 115–125.
Leakey, A.D., K. A. Bishop and E.A. Ainsworth, 2012: A multi-biome gap in understanding of crop and ecosystem responses to elevated CO2. Curr. Opin. Plant Biol. , 15 (3), 228–236, doi:10.1016/j.pbi.2012.01.009.
Lebel, L., et al., 2018: Managing the risks from the water-related impacts of extreme weather and uncertain climate change on inland aquaculture in Northern Thailand. Water Int. , 43 (2), 257–280, doi:10.1080/02508060.2017.1416446.
Lebel, L., et al., 2020: Aquaculture farmers’ perceptions of climate-related risks in the Mekong Region. Reg Environ Change, 20 (3), 95, doi:10.1007/s10113-020-01688-5.
Leck, H., D. Conway, M. Bradshaw and J. Rees, 2015: Tracing the water-energy-food nexus: description, theory and practice. Geogr. Compass, 9 (8), 445–460, doi:10.1111/gec3.12222.
Lee, D.M. and K.S. Lyon, 2004: Assessing climate change impacts, benefits of mitigation, and uncertainties on major global forest regions under multiple socioeconomic and emissions scenarios. South Econ J, 70, 467–489, doi:10.1088/1748-9326/aa63 fc.
Lee, H.L., Y.P. Lin and J.R. Petway, 2018: Global agricultural trade pattern in a warming world: regional realities. Sustainability, 10 (8), 2763, doi:10.3390/su10082763.
Legave, J.M., et al., 2015: Differentiated responses of apple tree floral phenology to global warming in contrasting climatic regions. Front. Plant Sci. , 6, 1054, doi:10.3389/fpls.2015.01054.
Leguizamón, A., 2014: Modifying Argentina: GM soy and socio-environmental change. Geoforum, 53, 149–160, doi:10.1016/j.geoforum.2013.04.001.
Lehnert, L.W., et al., 2016: Climate variability rather than overstocking causes recent large scale cover changes of Tibetan pastures. Sci Rep, 6, 24367, doi:10.1038/srep24367.
Leibar, U., et al., 2017: Grape yield and quality responses to simulated year 2100 expected climatic conditions under different soil textures. J. Sci. Food Agric, 97 (8), 2633–2640, doi:10.1002/jsfa.8086.
Leitão, S.T., et al., 2019: Maize open-pollinated populations physiological improvement: validating tools for drought response participatory selection. Sustainability, 11 (21), doi:10.3390/su11216081.
Lemasson, A.J., J.M. Hall-Spencer, V. Kuri and A.M. Knights, 2019: Changes in the biochemical and nutrient composition of seafood due to ocean acidification and warming. Mar. Environ. Res. , 143, 82–92, doi:10.1016/j.marenvres.2018.11.006.
Lemonnier, P. and E.A. Ainsworth, 2018: Crop responses to rising atmospheric [CO2] and global climate change. In: Food Security and Climate Change[Yadav, S.S., R.J. Redden, J.L. Hatfield, A.A. Ebert and D. Hunter(eds.)]. Wiley Blackwell, Hoboken, NJ, USA and Chichester, UK, pp. 51–69. ISBN 978-1119180647.
Lemos, M.C., et al., 2016: Linking development to climate adaptation: Leveraging generic and specific capacities to reduce vulnerability to drought in NE Brazil. Glob. Environ. Change. , 39, 170–179, doi:10.1016/j.gloenvcha.2016.05.001.
Lennox, E., 2015: Double exposure to climate change and globalization in a Peruvian highland community. Soc. Nat. Resour. , 28 (7), 781–796, doi:10.1080/08941920.2015.1024364.
Lenoir, J., et al., 2019: Species better track the shifting isotherms in the oceans than on lands. bioRxiv, 765776, doi:10.1101/765776.
Lenton, T.M., et al., 2019: Climate tipping points - too risky to bet against. Nature, 575 (7784), 592–595, doi:10.1038/d41586-019-03595-0.
Leolini, L., et al., 2019: Modelling sugar and acid content in Sangiovese grapes under future climates: an Italian case study. Clim. Res. , 78 (3), 211–224, doi:10.3354/cr01571.
Leon-Munoz, J., M. A. Urbina, R. Garreaud and J.L. Iriarte, 2018: Hydroclimatic conditions trigger record harmful algal bloom in western Patagonia (summer 2016). Sci Rep, 8 (1), 1330, doi:10.1038/s41598-018-19461-4.
Leon, J.X., et al., 2015: Supporting local and traditional knowledge with science for adaptation to climate change: lessons learned from participatory three-dimensional modeling in BoeBoe, Solomon Islands. Coast Manage, 43 (4), 424–438, doi:10.1080/08920753.2015.1046808.
Leonelli, G.C., 2018: GMO risks, food security, climate change and the entrenchment of neo-liberal legal narratives. Transnatl. Leg. Theory, 9, 302–315, doi:10.1080/20414005.2018.1572313.
Lepofsky, D., K. Lertzman, D. Hallett and R. Mathewes, 2005: Climate change and culture change on the southern coast of British Columbia 2400-1200 CAL. BP: an hypothesis. Am. antiq. , 70 (2), 267–293, doi:10.2307/40035704.
Lesk, C., et al., 2017: Threats to North American forests from southern pine beetle with warming winters. Nat Clim Chang, 7, 713–717, doi:10.1038/nclimate3375.
Lesk, C., P. Rowhani and N. Ramankutty, 2016: Influence of extreme weather disasters on global crop production. Nature, 529 (7584), 84–87, doi:10.1038/nature16467.
Lesnikowski, A., et al., 2016: National-level progress on adaptation. Nat. Clim. Chang. , 6 (3), 261–264, doi:10.1038/Nclimate2863.
Leßmeister, A., et al., 2015: Substitution of the most important and declining wild food species in southeast Burkina Faso. Flora Veg. Sudano-sambesica, 18, 11–20, doi:10.21248/fvss.18.29.
Lévesque, E., et al., 2012: Trends in vegetation dynamics and impacts on berry productivity. In: Nunavik and Nunatsiavut: From Science to Policy. An Integrated Regional Impact Study (IRIS) of Climate Change and Modernization[Allard, M. and M. Lemay(eds.)]. ArcticNet, Quebec City, Canada, pp. 223–247.
Levi, M., T. Kjellstrom and A. Baldasseroni, 2018: Impact of climate change on occupational health and productivity: a systematic literature review focusing on workplace heat. Med. Lav. , 109 (3), 163–179, doi:10.23749/mdl.v109i3.6851.
Levy, B.S. and C. Roelofs, 2019: Impacts of climate change on workers’ health and safety. In: Oxford Research Encyclopedia of Global Public Health. doi:10.1093/acrefore/9780190632366.013.39.
Lhorente, J.P., M. Araneda, R. Neira and J.M. Yanez, 2019: Advances in genetic improvement for salmon and trout aquaculture: the Chilean situation and prospects. Rev. Aquac. , 11 (2), 340–353, doi:10.1111/raq.12335.
Li, C., et al., 2020: Syndromes of production in intercropping impact yield gains. Nat. Plants, 6 (6), 653–660, doi:10.1038/s41477-020-0680-9.
Li, F.B., et al., 2019: Effects of rice-fish co-culture on oxygen consumption in intensive aquaculture pond. Rice Sci, 26 (1), 50–59, doi:10.1016/j.rsci.2018.12.004.
Li, M., et al., 2014: Efficient and heritable gene targeting in tilapia by CRISPR/Cas9. Nat. Genet. , 197 (2), 591–599, doi:10.1534/genetics.114.163667.
Li, T. et al., 2015: Uncertainties in predicting rice yield by current crop models under a wide range of climatic conditions. Glob Chang Biol, 21 (3), 1328-1341, doi:10.1111/gcb.12758. Li, W.H., et al., 2018a: Ecosystem structure, functioning and stability under climate change and grazing in grasslands: current status and future prospects. Curr. Opin. Environ. Sustain. , 33, 124–135, doi:10.1016/j.cosust.2018.05.008.
Li, X., et al., 2018b: Asparagopsis taxiformis decreases enteric methane production from sheep. Anim. Prod. Sci. , 58 (4), 681–681, doi:10.1071/an15883.
Li, X. and K.H.M. Siddique (eds.), 2018: Future Smart Food: Rediscovering Hidden Treasures of Neglected and Underutilized Species for Zero Hunger in Asia. Food and Agriculture Organization of the United Nations, Bangkok, Thailand, ISBN 978-9251304952. 216 pp.
Li, X.L., et al., 2017: Patterns of herders’ adaptation to changes in social-ecological systems across northern China’s grasslands over the past three decades. Rangel. J. , 39 (4), 317–328, doi:10.1071/Rj16070.
Liao, C. and D.G. Brown, 2018: ScienceDirect Assessments of synergistic outcomes from sustainable intensification of agriculture need to include smallholder livelihoods with food production and ecosystem services. Curr. Opin. Environ. Sustain. , 32 (December 2017), 53–59, doi:10.1016/j.cosust.2018.04.013.
Liao, C. and D. Fei, 2017: Sedentarization as constrained adaptation: evidence from pastoral regions in far Northwestern China. Hum Ecol, 45 (1), 23–35, doi:10.1007/s10745-016-9872-3.
Liao, C., S. Jung, D.G. Brown and A. Agrawal, 2020: Spatial patterns of large-scale land transactions and their potential socio-environmental outcomes in Cambodia, Ethiopia, Liberia, and Peru. Land Degrad Dev, 31 (10), 1241–1251, doi:10.1002/ldr.3544.
Liao, C., et al., 2021: Carbon emissions from the global land rush and potential mitigation. Nat. Food, 2 (1), 15–18, doi:10.1038/s43016-020-00215-3.
Liebman, M. and L.A. Schulte, 2015: Enhancing agroecosystem performance and resilience through increased diversification of landscapes and cropping systems. Elem. Sci. Anthropocene, 3, 41, doi:10.12952/journal.elementa.000041.
Lim-Camacho, L., et al., 2015: Facing the wave of change: stakeholder perspectives on climate adaptation for Australian seafood supply chains. Reg Environ Change, 15 (4), 595–606, doi:10.1007/s10113-014-0670-4.
Lin, B.B., et al., 2017: Adaptation pathways in coastal case studies: lessons learned and future directions. Coast Manage, 45 (5), 384–405, doi:10.1080/08920753.2017.1349564.
Lincoln, N.K., 2019: Learning from indigenous agriculture. Nat. Sustain. , 2 (3), 167–168, doi:10.1038/s41893-019-0234-1.
Lincoln, N.K., 2020: Agroforestry form and ecological adaptation in ancient Hawai’i: Extent of the pakukui swidden system of Hamakua, Hawai’i Island. Agric Syst , 181, 102808, doi:10.1016/j.agsy.2020.102808.
Link, J.S., R. Griffis and S. Busch (eds.), 2015: NOAA Fisheries Climate Science Strategy. NOAA Technical Memorandum NMFS-F/SPO, Vol. 155. U.S. Dept. of Commerce, Silver Spring, MD, USA. 70 pp.
Lipper, L., R. DeFries and L. Bizikova, 2020: Shedding light on the evidence blind spots confounding the multiple objectives of SDG 2. Nat. Plants, 6 (10), 1203–1210, doi:10.1038/s41477-020-00792-y.
Lipper, L., et al. (ed.), 2017: Climate Smart Agriculture: Building Resilience to Climate Change, Natural Resource Management and Policy. Springer Nature, Cham, Switzerland, ISBN 978-3319611938. 630 pp.
Lipper, L., et al., 2014: Climate-smart agriculture for food security. Nat. Clim. Chang. , 4 (12), 1068–1072, doi:10.1038/Nclimate2437.
Little, D.C., R.W. Newton and M.C. Beveridge, 2016: Aquaculture: a rapidly growing and significant source of sustainable food? Status, transitions and potential. Proc. Nutr. Soc. , 75 (3), 274–286, doi:10.1017/S0029665116000665.
Liu, C., N. Hofstra and E. Franz, 2013: Impacts of climate change on the microbial safety of pre-harvest leafy green vegetables as indicated by Escherichia coli O157 and Salmonella spp. Int. J. Food Microbiol. , 163 (2-3), 119–128, doi:10.1016/j.ijfoodmicro.2013.02.026.
Liu, C., et al., 2019a: Vertical redistribution of salt and layered changes in global ocean salinity. Nat Commun, 10 (1), 3445, doi:10.1038/s41467-019-11436-x.
Liu, S., et al., 2018a: Retrospective evaluation of marker-assisted selection for resistance to bacterial cold water disease in three generations of a commercial rainbow trout breeding population. Front. Genet. , 9 (286), 286, doi:10.3389/fgene.2018.00286.
Liu, S.L., et al., 2019b: Extreme stress threatened double rice production in Southern China during 1981–2010. Theor. Appl. Climatol. , 137 (3), 1987–1996, doi:10.1007/s00704-018-2719-7.
Liu, Y., et al., 2017: Do invasive alien plants benefit more from global environmental change than native plants?Glob Chang Biol, 23 (8), 3363–3370, doi:10.1111/gcb.13579.
Liu, Y., et al., 2018b: Impact of short-term climate effects and sea ice coverage variation on Japanese scallop aquaculture in Saroma Lake, Japan. Aquac Res, 49 (5), 1752–1767, doi:10.1111/are.13630.
Liu, Y., et al., 2020: Impact of climate extremes on suitability dynamics for Japanese scallop aquaculture in Shandong, China and Funka Bay, Japan. Sustainability, 12 (3), 833, doi:10.3390/su12030833.
Liu, Y.J., et al., 2018c: Modelling the impacts of climate change and crop management on phenological trends of spring and winter wheat in China. Agric For Meteorol, 248, 518–526, doi:10.1016/j.agrformet.2017.09.008.
Lkhagvadorj, D., M. Hauck, C. Dulamsuren and J. Tsogtbaatar, 2013: Pastoral nomadism in the forest-steppe of the Mongolian Altai under a changing economy and a warming climate. J. Arid Environ. , 88, 82–89, doi:10.1016/j.jaridenv.2012.07.019.
Lloyd, S.J., et al., 2018: A global-level model of the potential impacts of climate change on child stunting via income and food price in 2030. Environ. Health Perspect. , 126 (9), 97007, doi:10.1289/EHP2916.
Lo, A.Y., S. Liu, L.T.O. Cheung and F.K.S. Chan, 2019: Contested transformations: sustainable economic development and capacity for adapting to climate change. Ann. Am. Assoc. Geogr. , 110 (1), 223–241, doi:10.1080/24694452.2019.1625748.
Loaiza, T., U. Nehren and G. Gerold, 2016: REDD+ implementation in the Ecuadorian Amazon: Why land configuration and common-pool resources management matter. Policy Econ, 70, 67–79, doi:10.1016/j.forpol.2016.05.016.
Lobell, D.B. and C.B. Field, 2011: California perennial crops in a changing climate. Clim Change, 109, 317–333, doi:10.1007/s10584-011-0303-6.
Loboguerrero, A.M., et al., 2018: Bridging the gap between climate science and farmers in Colombia. Clim. Risk Manag. , 22, 67-81, doi:10.1016/j.crm.2018.08.001.
Loboguerrero, A.M., et al., 2020: Perspective article: actions to reconfigure food systems. Glob Food Sec, 26, 100432, doi:10.1016/j.gfs.2020.100432.
Lokuge, G. and D. Hilhorst, 2017: Outside the net: intersectionality and inequality in the fisheries of Trincomalee, Sri Lanka. Asian J. Women’s Stud. , 23 (4), 473–497, doi:10.1080/12259276.2017.1386839.
Loladze, I., 2014: Hidden shift of the ionome of plants exposed to elevated CO2 depletes minerals at the base of human nutrition. Elife, 3, e2245, doi:10.7554/eLife.02245.
Long, J.W., R.W. Goode and F.K. Lake, 2020: Recentering ecological restoration with tribal perspectives. Fremontia, 48 (1), 14–19.
López-i-Gelats, F., et al., 2015: Adaptation strategies of Andean pastoralist households to both climate and non-climate changes. Hum Ecol, 43 (2), 267–282, doi:10.1007/s10745-015-9731-7.
López-i-Gelats, F., E.D.G. Fraser, J.F. Morton and M.G. Rivera-Ferre, 2016: What drives the vulnerability of pastoralists to global environmental change? A qualitative meta-analysis. Glob. Environ. Chang. , 39, 258–274, doi:10.1016/j.gloenvcha.2016.05.011.
Loring, P.A. and S.C. Gerlach, 2009: Food, culture, and human health in Alaska: an integrative health approach to food security. Environ Sci Policy, 12 (4), 466–478, doi:10.1016/j.envsci.2008.10.006.
Lotter, D. and D. Maitre, 2014: Modelling the distribution of Aspalathus linearis (Rooibos tea): implications of climate change for livelihoods dependent on both cultivation and harvesting from the wild. Ecol Evol, 4 (8), 1209–1221, doi:10.1002/ece3.985.
Lotze, H.K., et al., 2019: Global ensemble projections reveal trophic amplification of ocean biomass declines with climate change. Proc. Natl. Acad. Sci. U. S. A. , 116 (26), 12907–12912, doi:10.1073/pnas.1900194116.
Love, D.C., M.S. Uhl and L. Genello, 2015: Energy and water use of a small-scale raft aquaponics system in Baltimore, Maryland, United States. Aquac Eng, 68, 19–27, doi:10.1016/j.aquaeng.2015.07.003.
Lovell, S.T., 2010: Multifunctional urban agriculture for sustainable land use planning in the United States. Sustainability, 2 (8), 2499–2522, doi:10.3390/su2082499.
Lovell, S.T., et al., 2018: Temperate agroforestry research: considering multifunctional woody polycultures and the design of long-term field trials. Agrofor. Syst. , 92 (5), 1397–1415, doi:10.1007/s10457-017-0087-4.
Lowder, S.K., M.V. Sánchez and R. Bertini, 2019: Farms, Family Farms, Farmland Distribution and Farm Labour: What do we Know Today?FAO Agricultural Development Economics Working Paper. Food and Agriculture Organization of the United Nations, ISBN 978-9251319703. 17/08/2021, 70 pp.
Lowe, B.S., et al., 2019: Adapting to change in inland fisheries: analysis from Lake Tanganyika, East Africa. Reg Environ Change, 19 (6), 1765–1776, doi:10.1007/s10113-019-01516-5.
Lu, M., et al., 2013: Responses of ecosystem carbon cycle to experimental warming: a meta-analysis. Ecology, 94 (3), 726–738, doi:10.1890/12-0279.1.
Luedeling, E., R. Kindt, N.I. Huth and K. Koenig, 2014: Agroforestry systems in a changing climate—challenges in projecting future performance. Curr. Opin. Environ. Sustain. , 6, 1–7, doi:10.1016/j.cosust.2013.07.013.
Lujan, E., et al., 2018: Climate Change in Port Heiden, Alaska-Strategies for Community Health. Alaska Native Tribal Health Consortium, https://anthc.org/wp-content/uploads/2018/10/ANTHC-Port-Heiden_Final.pdf. Accessed 2021 . (59 pp).
Luna-Gonzalez, D.V. and M. Sorensen, 2018: Higher agrobiodiversity is associated with improved dietary diversity, but not child anthropometric status, of Mayan Achi people of Guatemala. Public Health Nutr. , 21 (11), 2128–2141, doi:10.1017/S1368980018000617.
Lund, J.F., E. Sungusia, M.B. Mabele and A. Scheba, 2017: Promising Change, Delivering Continuity: REDD plus as Conservation Fad. World Dev, 89, 124–139, doi:10.1016/j.worlddev.2016.08.005.
Lunstrum, E., 2016: Green grabs, land grabs and the spatiality of displacement: Eviction from Mozambique’s Limpopo National Park. Area, 48 (2), 142–152, doi:10.1111/area.12121.
Lunstrum, E., P. Bose and A. Zalik, 2016: Environmental displacement: the common ground of climate change, extraction and conservation. Area, 48 (2), 130–133, doi:10.1111/area.12193.
Luo, Q.Y., M. Bange and D. Johnston, 2016: Environment and cotton fibre quality. Clim Change, 138 (1-2), 207–221, doi:10.1007/s10584-016-1715-0.
Lwasa, S., et al., 2014: Urban and peri-urban agriculture and forestry: transcending poverty alleviation to climate change mitigation and adaptation. Urban Clim, 7, 92–106, doi:10.1016/j.uclim.2013.10.007.
Lwasa, S., et al., 2015: A meta-analysis of urban and peri-urban agriculture and forestry in mediating climate change. Curr. Opin. Environ. Sustain. , 13, 68–73, doi:10.1016/j.cosust.2015.02.003.
Lydersen, C., 2018: Walrus: Odobenus rosmarus. In: Encyclopedia of Marine Mammals[Würsig, B., J.G.M. Thewissen and K.M. Kovacs(eds.)], 3rd edn. Academic, London, UK, San Diego, CA, USA, Cambridge, MA, USA, and Oxford, UK, pp. 1045–1048. doi:10.1016/B978-0-12-804327-1.00020-0.
Lyman, N.B., et al., 2013: Neglecting rice milling yield and quality underestimates economic losses from high-temperature stress. Plos One, 8 (8), e72157, doi:10.1371/journal.pone.0072157.
Lynch, A.J., et al., 2017: Inland fisheries–Invisible but integral to the UN Sustainable Development Agenda for ending poverty by 2030. Glob. Environ. Chang. , 47, 167–173.
Lynch, A.J., et al., 2016: Climate change effects on North American inland fish populations and assemblages. Fisheries, 41 (7), 346–361, doi:10.1080/03632415.2016.1186016.
Lynn, K., et al., 2013: The impacts of climate change on tribal traditional foods. In: Climate Change and Indigenous Peoples in the United States: Impacts, Experiences, and Actions[Maldonado, J.K., B. Colombi and R. Pandya(eds.)]. Springer, Cham, Switzerland, pp. 37–48. ISBN 978-3319052656.
Lyons, K., C. Richards and P. Westoby, 2014: The Darker Side of Green Plantation Forestry and Carbon Violence in Uganda: The Case of Green Resources’ Forestry-Based Carbon Markets. The Oakland Institute, Oakland, CA, https://www.oaklandinstitute.org/sites/oaklandinstitute.org/files/Report_DarkerSideofGreen_hirez.pdf. Accessed 2021 . (19 pp).
M’Kaibi, F.K., N.P. Steyn, S. Ochola and L. Du Plessis, 2015: Effects of agricultural biodiversity and seasonal rain on dietary adequacy and household food security in rural areas of Kenya. BMC Public Health, 15, 422, doi:10.1186/s12889-015-1755-9.
Müller, C., et al., 2017: Global gridded crop model evaluation: benchmarking, skills, deficiencies and implications. Geosci. Model Dev. , 10 (4), 1403–1422.
Müller, C., et al., 2021a: Exploring uncertainties in global crop yield projections in a large ensemble of crop models and CMIP5 and CMIP6 climate scenarios. Environ. Res. Lett. , 16 (3), doi:10.1088/1748-9326/abd8 fc.
Müller, M.F., et al., 2021b: Impact of transnational land acquisitions on local food security and dietary diversity. Proc. Natl. Acad. Sci. , 118 (4), e2020535118–e2020535118, doi:10.1073/pnas.2020535118.
Münke, C., et al., 2015: Wild ideas in food. In: The Routledge Handbook on Sustainable Food and Gastronomy[Sloan, P., W. Legrand and C. Hindley(eds.)]. Routledge, Abingdon, UK, and New York, NY, USA, pp. 206–213. ISBN 978-0415702553.
Mabe, F.N. and A. Asase, 2020: Climate change adaptation strategies and fish catchability: The case of inland artisanal fishers along the Volta Basin in Ghana. Fish. Res. , 230, 105675, doi:10.1016/j.fishres.2020.105675.
Mabe, F.N., S. Nashiru, E. Mummuni and V.F. Boateng, 2019: The nexus between land acquisition and household livelihoods in the Northern region of Ghana. Land Use Policy, 85 (April 2018), 357–367, doi:10.1016/j.landusepol.2019.03.043.
Macabuhay, A., et al., 2018: Can elevated CO2 buffer the effects of heat waves on wheat in a dryland cropping system?Environ. Exp. Bot. , 155, 578–588, doi:10.1016/j.envexpbot.2018.07.029.
Mach, K.J., et al., 2019: Climate as a risk factor for armed conflict. Nature, 571 (7764), 193–197, doi:10.1038/s41586-019-1300-6.
Machado, L., et al., 2016: Dose-response effects of Asparagopsis taxiformis and Oedogonium sp. on in vitro fermentation and methane production. J Appl Phycol, 28 (2), 1443–1452, doi:10.1007/s10811-015-0639-9.
Maciejczak, M. and J. Mikiciuk, 2019: Climate change impact on viticulture in Poland. Int. J. Clim. Chang. Strateg. Manag. , 11 (2), 254–264, doi:10.1108/Ijccsm-02-2018-0021.
Macko, S.A., C. Fantasia and G.J. Xue, 2017: Potential global economic impacts of ocean acidification. In: International Marine Economy: Law and Policy[Nordquist, M.H., J.N. Moore and R. Long(eds.)]. Brill Nijhoff, Leiden, the Netherlands and Boston, MA, USA, pp. 79–97. ISBN 978-9004323438.
Maeno, K.O. and M. A. Ould Babah Ebbe, 2018: Aggregation Site Choice by Gregarious Nymphs of the Desert Locust, Schistocerca gregaria, in the Sahara Desert of Mauritania. Insects, 9 (3), 99–99, doi:10.3390/insects9030099.
Maezumi, S.Y., et al., 2018: The legacy of 4,500 years of polyculture agroforestry in the eastern Amazon. Nat. Plants, 4 (8), 540–547, doi:10.1038/s41477-018-0205-y.
Magliocca, N.R., Q.V. Khuc, A. De Bremond and E.A. Ellicott, 2020: Direct and indirect land-use change caused by large-scale land acquisitions in Cambodia. Environ. Res. Lett. , 15 (2), 24010–24010, doi:10.1088/1748-9326/ab6397.
Magnan, A.K., 2014: Avoiding maladaptation to climate change: towards guiding principles. S.A.P. I. EN. S. , 7 (1), 1-12. http://journals.openedition.org/sapiens/1680.
Magni, G., 2017: Indigenous knowledge and implications for the sustainable development agenda. Eur. J. Educ. , 52 (4), 437–447, doi:10.1111/ejed.12238.
Magrini, M.-B., et al., 2016: Why are grain-legumes rarely present in cropping systems despite their environmental and nutritional benefits? Analyzing lock-in in the French agrifood system. Ecol. Econ. , 126, 152–162, doi:10.1016/j.ecolecon.2016.03.024.
Maharjan, S. and K. Maharjan, 2018: Roles and contributions of community seed banks in climate adaptation in Nepal. Dev. Pract. , 28 (2), 292–302, doi:10.1080/09614524.2018.1418838.
Maikhuri, R.K., et al., 2018: Assessment of climate change impacts and its implications on medicinal plants-based traditional healthcare system in Central Himalaya, India. Iran. J. Sci. Technol. Trans. A: Sci. , 42 (A4), 1827–1835, doi:10.1007/s40995-017-0354-2.
Maina, J., et al., 2016: Integrating social-ecological vulnerability assessments with climate forecasts to improve local climate adaptation planning for coral reef fisheries in Papua New Guinea. Reg Environ Change, 16 (3), 881–891, doi:10.1007/s10113-015-0807-0.
Makkar, H.P.S., 2018: Review: feed demand landscape and implications of food-not feed strategy for food security and climate change. Animal, 12 (8), 1744–1754, doi:10.1017/S175173111700324X.
Makondo, C.C. and D.S.G. Thomas, 2018: Climate change adaptation: linking indigenous knowledge with western science for effective adaptation. Environ Sci Policy, 88, 83–91, doi:10.1016/j.envsci.2018.06.014.
Mallory, C.D. and M.S. Boyce, 2018: Observed and predicted effects of climate change on Arctic caribou and reindeer. Environ. Rev. , 26 (1), 13–25, doi:10.1139/er-2017-0032.
Manderscheid, R., M. Erbs and H.J. Weigel, 2014: Interactive effects of free-air CO2 enrichment and drought stress on maize growth. Eur J Agron, 52, 11–21, doi:10.1016/j.eja.2011.12.007.
Mangi, S.C., et al., 2018: The economic impacts of ocean acidification on shellfish fisheries and aquaculture in the United Kingdom. Environ Sci Policy, 86, 95–105, doi:10.1016/j.envsci.2018.05.008.
Manners, R. and J. van Etten, 2018: Are agricultural researchers working on the right crops to enable food and nutrition security under future climates?Glob. Environ. Change, 53, 182–194.
Manners, R., et al., 2021: Suitability of root, tuber, and banana crops in Central Africa can be favoured under future climates. Agric Syst , 193, 103246, doi:10.1016/j.agsy.2021.103246.
Mantri, V.A., M.G. Kavale and M. A. Kazi, 2020: Seaweed biodiversity of India: reviewing current knowledge to identify gaps, challenges, and opportunities. Diversity, 12 (1), 13, doi:10.3390/d12010013.
Mantyka-Pringle, C.S., et al., 2016: Prioritizing management actions for the conservation of freshwater biodiversity under changing climate and land-cover. Biol. Conserv. , 197, 80–89, doi:10.1016/j.biocon.2016.02.033.
Maranz, S. and Z. Wiesman, 2003: Evidence for indigenous selection and distribution of the shea tree, Vitellaria paradoxa, and its potential significance to prevailing parkland savanna tree patterns in sub-Saharan Africa north of the equator. J. Biogeogr. , 30 (10), 1505–1516, doi:10.1046/j.1365-2699.2003.00892.x.
Maraseni, T.N., P.R. Neupane, F. Lopez-Casero and T. Cadman, 2014: An assessment of the impacts of the REDD+ pilot project on community forests user groups (CFUGs) and their community forests in Nepal. J Environ Manage, 136, 37–46, doi:10.1016/j.jenvman.2014.01.011.
Marcogliese, D.J., 2016: The distribution and abundance of parasites in aquatic ecosystems in a changing climate: more than just temperature. Integr. Comp. Biol. , 56 (4), 611–619, doi:10.1093/icb/icw036.
Marcos-Martinez, R., B.A. Bryan, J.D. Connor and D. King, 2017: Agricultural land-use dynamics: assessing the relative importance of socioeconomic and biophysical drivers for more targeted policy. Land Use Policy, 63, 53–66, doi:10.1016/j.landusepol.2017.01.011.
Marín, D., J. Armengol, J.M. Escalona and D. Gramaje, 2021: Challenges of viticulture adaptation to global change : tackling the issue from the roots. Aust J Grape Wine Res, 8–25, doi:10.1111/ajgw.12463.
Marini, L., et al., 2020: Crop rotations sustain cereal yields under a changing climate. Environ. Res. Lett. , 15 (12), 124011–124011, doi:10.1088/1748-9326/abc651.
Markandya, A. and M. González-Eguino, 2019: Integrated Assessment for Identifying Climate Finance Needs for Loss and Damage: A Critical Review. In: Loss and Damage from Climate Change: Concepts, Methods and Policy Options[Mechler, R., L.M. Bouwer, T. Schinko, S. Surminski and J. Linnerooth-Bayer(eds.)]. Springer International Publishing, Cham, pp. 343–362. ISBN 978-3319720265.
Markon, C., et al., 2018: Alaska. In: In Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment [Reidmiller, D.R., et al.(ed.)]. Vol.II, US Global Change Research Program, Washington, DC, USA, pp. 1185–1241.
Marselis, S.M., et al., 2017: Agricultural land displacement and undernourishment. J Clean Prod, 161, 619–628, doi:10.1016/j.jclepro.2017.05.125.
Marteau, T.M., G.J. Hollands and P.C. Fletcher, 2012: Changing human behavior to prevent disease: the importance of targeting automatic processes. Science, 337 (6101), 1492–1495, doi:10.1126/science.1226918.
Martellozzo, F., et al., 2014: Urban agriculture: a global analysis of the space constraint to meet urban vegetable demand. Environ. Res. Lett. , 9 (6), 64025, doi:10.1088/1748-9326/9/6/064025.
Martínez-Lüscher, J., et al., 2016: Sensitivity of grapevine phenology to water availability, temperature and CO2 concentration. Front. Environ. Sci. , 4, doi:10.3389/fenvs.2016.00048.
Martínez-Paz, J.M., I. Banos-González, J. Martínez-Fernández and M.Á. Esteve-Selma, 2019: Assessment of management measures for the conservation of traditional irrigated lands: The case of the Huerta of Murcia (Spain). Land Use Policy, 81, 382–391, doi:10.1016/j.landusepol.2018.10.050.
Martin, G., et al., 2020: Potential of multi-species livestock farming to improve the sustainability of livestock farms: a review. Agric Syst , 181, 102821, doi:10.1016/j.agsy.2020.102821.
Martin, G. and M. Willaume, 2016: A diachronic study of greenhouse gas emissions of French dairy farms according to adaptation pathways. Agric Ecosyst Environ, 221, 50–59, doi:10.1016/j.agee.2016.01.027.
Martin, M. and E. Molin, 2019: Environmental assessment of an urban vertical hydroponic farming system in Sweden. Sustain. (switzerland) , 11 (15), doi:10.3390/su11154124.
Martinez-Harms, M.J., et al., 2017: Scenarios for land use and ecosystem services under global change. Ecosyst. Serv. , 25, 56–68, doi:10.1016/j.ecoser.2017.03.021.
Martinez, M., et al., 2018: Measuring the effects of temperature rise on Mediterranean shellfish aquaculture. Ecol Indic, 88, 71–78, doi:10.1016/j.ecolind.2018.01.002.
Martins, I.M., L.C. Gammage, A. Jarre and M. A. Gasalla, 2019: Different but similar? Exploring vulnerability to climate change in Brazilian and South African small-scale fishing communities. Hum Ecol, 47 (4), 515–526, doi:10.1007/s10745-019-00098-4.
Martins, I.M. and M. A. Gasalla, 2018: Perceptions of climate and ocean change impacting the resources and livelihood of small-scale fishers in the South Brazil bight. Clim Change, 147 (3-4), 441–456, doi:10.1007/s10584-018-2144-z.
Martos-Sitcha, J.A., et al., 2019: Tissue-Specific Orchestration of Gilthead Sea Bream Resilience to Hypoxia and High Stocking Density. Front. Physiol. , 10 (840), doi:10.3389/fphys.2019.00840.
Marushka, L., et al., 2019: Potential impacts of climate-related decline of seafood harvest on nutritional status of coastal First Nations in British Columbia, Canada. PLoS ONE, 14 (2), e211473, doi:10.1371/journal.pone.0211473.
Mase, A.S. and L.S. Prokopy, 2014: Unrealized potential: a review of perceptions and use of weather and climate information in agricultural decision making. Weather Clim. Soc. , 6 (1), 47–61, doi:10.1175/Wcas-D-12-00062.1.
Masutomi, Y., et al., 2019: Rice grain quality degradation and economic loss due to global warming in Japan. Environ. Res. Commun. , 1 (12), 121003, doi:10.1088/2515-7620/ab52e7.
Matiu, M., D.P. Ankerst and A. Menzel, 2017: Interactions between temperature and drought in global and regional crop yield variability during 1961-2014. PLoS ONE, 12 (5), e178339, doi:10.1371/journal.pone.0178339.
Mattick, C.S., A.E. Landis, B.R. Allenby and N.J. Genovese, 2015: Anticipatory Life Cycle Analysis of In Vitro Biomass Cultivation for Cultured Meat Production in the United States. Environ. Sci. Technol. , 49 (19), 11941–11949, doi:10.1021/acs.est.5b01614.
Mattos, L.M., et al., 2014: Climate Changes and Potential Impacts on Quality of Fruit and Vegetable Crops. In: Emerging Technologies and Management of Crop Stress Tolerance[Ahmad, P. and S. Rasool(eds.)]. Academic, San Diego, CA, USA, London, UK, and Waltham, MA, USA, pp. 467–486. ISBN 978-0128008768.
Matzrafi, M., 2019: Climate change exacerbates pest damage through reduced pesticide efficacy. Pest. Manag. Sci. , 75 (1), 9–13, doi:10.1002/ps.5121.
Matzrafi, M., et al., 2016: Climate change increases the risk of herbicide-resistant weeds due to enhanced detoxification. Planta, 244 (6), 1217–1227, doi:10.1007/s00425-016-2577-4.
Mayer, A., et al., 2019a: Measuring Progress towards a Circular Economy: A Monitoring Framework for Economy-wide Material Loop Closing in the EU28. J Ind Ecol, 23 (1), 62–76, doi:10.1111/jiec.12809.
Mayer, D.G., K. A. Chandra and J.R. Burnett, 2019b: Improved crop forecasts for the Australian macadamia industry from ensemble models. Agric Syst , 173, 519–523, doi:10.1016/j.agsy.2019.03.018.
Mazaris, A.D. and B. Germond, 2018: Bridging the gap between climate change and maritime security: Towards a comprehensive framework for planning. Sci. Total Environ. , 635, 1076–1080, doi:10.1016/j.scitotenv.2018.04.136.
Mbow, C., C. Rosenzweig, L.G. Barioni, T.G. Benton, M. Herrero, M. Krishnapillai, E. Liwenga, P. Pradhan, M.G. Rivera-Ferre, T. Sapkota, F.N. Tubiello, Y. Xu, 2019: Food security. In: Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems[Shukla, P. R., J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H. O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi and J. Malley (eds.)], pp. In press.
Mbow, C., et al., 2014: Achieving mitigation and adaptation to climate change through sustainable agroforestry practices in Africa. Curr. Opin. Environ. Sustain. , 6, 8–14, doi:10.1016/j.cosust.2013.09.002.
McCabe, R.M., et al., 2016: An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. , 43 (19), 10366–10376, doi:10.1002/2016GL070023.
Mccall, M.K., 2016: Beyond “landscape” in REDD plus : the imperative for “territory”. World Dev, 85, 58–72, doi:10.1016/j.worlddev.2016.05.001.
McCoy, D., et al., 2017: Large-scale climatic effects on traditional Hawaiian fishpond aquaculture. PLoS ONE, 12 (11), e187951, doi:10.1371/journal.pone.0187951.
McDaniel, M.D., L.K. Tiemann and A.S. Grandy, 2014: Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol Appl, 24 (3), 560–570, doi:10.1890/13-0616.1.
McDonald, K.S., et al., 2019: Proactive, reactive, and inactive pathways for scientists in a changing world. Earths Future, 7 (2), 60–73, doi:10.1029/2018ef000990.
McDowell, N.G., et al., 2020: Pervasive shifts in forest dynamics in a changing world. Science, 368 (6494), doi:10.1126/science.aaz9463.
McGill, B.M., S.K. Hamilton, N. Millar and G.P. Robertson, 2018: The greenhouse gas cost of agricultural intensification with groundwater irrigation in a Midwest U.S. row cropping system. Glob Chang Biol, 24 (12), 5948–5960, doi:10.1111/gcb.14472.
McGrath, D. and L. Costello, 2015: Integrating fishers’ ecological knowledge and the ecosystem based mangement of tropical inland fisheries: an Amazon case study. In: Fishers’ knowledge and the ecosystem approach to fisheries: applications, experiences and lessons in Latin America[Fischer, J., J. Jorgensen, H. Josupeit, D. Kalikoski and C.M. Lucas(eds.)]. FAO, Rome, pp. 127–148.
McGrath, J.M., et al., 2015: An analysis of ozone damage to historical maize and soybean yields in the United States. Proc. Natl. Acad. Sci. U. S. A. , 112 (46), 14390–14395, doi:10.1073/pnas.1509777112.
McHunu, N., G. Lagerwall and A. Senzanje, 2018: Aquaponics in South Africa: Results of a national survey. Aquac. Reports, 12, 12–19, doi:10.1016/j.aqrep.2018.08.001.
McIntyre-Tamwoy, S., M. Fuary and A. Buhrich, 2013: Understanding climate, adapting to change: indigenous cultural values and climate change impacts in North Queensland. Local Environ, 18 (1), 91–109, doi:10.1080/13549839.2012.716415.
McIntyre, K.M., et al., 2017: Systematic assessment of the climate sensitivity of important human and domestic animals pathogens in Europe. Sci Rep, 7 (1), 7134, doi:10.1038/s41598-017-06948-9.
McIntyre, P.B., C.A. Reidy Liermann and C. Revenga, 2016: Linking freshwater fishery management to global food security and biodiversity conservation. Proc. Natl. Acad. Sci. U. S. A. , 113 (45), 12880–12885, doi:10.1073/pnas.1521540113.
Mcleod, E., et al., 2018: Raising the voices of Pacific Island women to inform climate adaptation policies. Mar. Policy. , 93, 178–185, doi:10.1016/j.marpol.2018.03.011.
McMillen, H., T. Ticktin and H.K. Springer, 2017: The future is behind us: traditional ecological knowledge and resilience over time on Hawai’i Island. Reg Environ Change, 17 (2), 579–592, doi:10.1007/s10113-016-1032-1.
McNamara, K.E. and L. Buggy, 2017: Community-based climate change adaptation: a review of academic literature. Local Environ, 22 (4), 443–460, doi:10.1080/13549839.2016.1216954.
McNamara, K.E., et al., 2020: An assessment of community-based adaptation initiatives in the Pacific Islands. Nat. Clim. Chang. , 10 (7), 628–639, doi:10.1038/s41558-020-0813-1.
McNamara, K.E. and S.S. Prasad, 2014: Coping with extreme weather: communities in Fiji and Vanuatu share their experiences and knowledge. Clim Change, 123 (2), 121–132, doi:10.1007/s10584-013-1047-2.
McNeeley, S.M., 2017: Sustainable Climate Change Adaptation in Indian Country. Weather. Clim. Soc. , 9 (3), 393–404, doi:10.1175/WCAS-D-16-0121.1.
McNeeley, S.M., et al., 2018: Anatomy of an interrupted irrigation season: Micro-drought at the Wind River Indian Reservation. Clim. Risk Manag. , 19, 61–82, doi:10.1016/j.crm.2017.09.004.
McNeeley, S.M., J.M. Friedman, T.A. Beeton and R.D. Thaxton, 2020: Cottonwoods, Water, and People-Integrating Analysis of Tree Rings with Observations of Elders from the Eastern Shoshone and Northern Arapaho Tribes of the Wind River Reservation. U.S. Geological Survey, Wyoming, 33 pp
McNeely, J.A. and G. Schroth, 2006: Agroforestry and biodiversity conservation - traditional practices, present dynamics, and lessons for the future. Biodivers Conserv, 15 (2), 549–554, doi:10.1007/s10531-005-2087-3.
Meade, E., M. A. Slattery and M. Garvey, 2019: Zoonotic antimicrobial resistance as a result of iatrogenic disease in companion animals - case study reports. J Infect Dis Epidemiol, 5, 73, doi:10.23937/2474-3658/1510073.
Mechiche-Alami, A., J. Yagoubi and K. A. Nicholas, 2021: Agricultural land acquisitions unlikely to address the food security needs of African countries. World Dev, 141, doi:10.1016/j.worlddev.2020.105384.
Medina, A., A. Rodriguez and N. Magan, 2015a: Climate change and mycotoxigenic fungi: impacts on mycotoxin production. Curr. Opin. Food Sci. , 5, 99–104, doi:10.1016/j.cofs.2015.11.002.
Medina, A., A. Rodriguez, Y. Sultan and N. Magan, 2015b: Climate change factors and Aspergillus flavus: effects on gene expression, growth and aflatoxin production. World Mycotoxin J. , 8 (2), 171–179, doi:10.3920/Wmj2014.1726.
Medina Hidalgo, D., et al., 2020: Sustaining healthy diets in times of change: linking climate hazards, food systems and nutrition security in rural communities of the Fiji Islands. Reg Environ Change, 20 (3), 73–73, doi:10.1007/s10113-020-01653-2.
Megersa, B., et al., 2014: Impacts of climate change and variability on cattle production in southern Ethiopia: Perceptions and empirical evidence. Agric Syst , 130, 23–34, doi:10.1016/j.agsy.2014.06.002.
Mehrabi, Z. and N. Ramankutty, 2019: Synchronized failure of global crop production. Nat. Ecol. Evol. , 3 (5), 780–786, doi:10.1038/s41559-019-0862-x.
Mehvar, S., et al., 2019: Climate change-driven losses in ecosystem services of coastal wetlands: A case study in the West coast of Bangladesh. Ocean. Coast. Manag. , 169, 273–283, doi:10.1016/j.ocecoaman.2018.12.009.
Meier, M., J. Fuhrer and A. Holzkamper, 2018: Changing risk of spring frost damage in grapevines due to climate change? A case study in the Swiss Rhone Valley. Int. J. Biometeorol. , 62 (6), 991–1002, doi:10.1007/s00484-018-1501-y.
Meijer, S.S., et al., 2015: The role of knowledge, attitudes and perceptions in the uptake of agricultural and agroforestry innovations among smallholder farmers in sub-Saharan Africa. Int. J. Agric. Sustain. , 13 (1), 40–54, doi:10.1080/14735903.2014.912493.
Mekonnen, M.M. and A.Y. Hoekstra, 2010: A global and high-resolution assessment of the green, blue and grey water footprint of wheat. Hydrol. Earth Syst. Sci. , 14 (7), 1259–1276, doi:10.5194/hess-14-1259-2010.
Mekuyie, M., A. Jordaan and Y. Melka, 2018: Understanding resilience of pastoralists to climate change and variability in the Southern Afar Region, Ethiopia. Clim. Risk Manag. , 20, 64–77, doi:10.1016/j.crm.2018.02.004.
Melbourne-Thomas, J., et al., 2021: Poleward bound: adapting to climate-driven species redistribution. Rev. Fish Biol. Fish. , doi:10.1007/s11160-021-09641-3.
Meldrum, G., et al., 2018: Climate change and crop diversity: farmers’ perceptions and adaptation on the Bolivian Altiplano. Environ. Dev. Sustain. , 20 (2), 703–730, doi:10.1007/s10668-016-9906-4.
Melillo, J.M., et al., 2017: Long-term pattern and magnitude of soil carbon feedback to the climate system in a warming world. Science, 358 (6359), 101–105, doi:10.1126/science.aan2874.
Mellin, C., et al., 2016: Forecasting marine invasions under climate change: Biotic interactions and demographic processes matter. Biol. Conserv. , 204, 459–467, doi:10.1016/j.biocon.2016.11.008.
Melvin, A.M., et al., 2017: Estimating wildfire response costs in Alaska’s changing climate. Clim Change, 141 (4), 783–795, doi:10.1007/s10584-017-1923-2.
Memmott, P., et al., 2013: Aboriginal Responses to Climate Change in Arid Zone Australia: Regional Understandings and Capacity Building for Adaptation. National Climate Change Adaptation Research Facility, Gold Coast, Australia.
Menchaca, A., P.C. Dos Santos-Neto, A.P. Mulet and M. Crispo, 2020: CRISPR in livestock: from editing to printing. Theriogenology, 150, 247–254, doi:10.1016/j.theriogenology.2020.01.063.
Mendenhall, E., et al., 2020: Climate change increases the risk of fisheries conflict. Mar. Policy. , 117, doi:10.1016/j.marpol.2020.103954.
Mendez, M., G. Flores-Haro and L. Zucker, 2020: The (in)visible victims of disaster: understanding the vulnerability of undocumented Latino/a and indigenous immigrants. Geoforum, 116, 50–62, doi:10.1016/j.geoforum.2020.07.007.
Menyuka, N.N., M. Sibanda and U. Bob, 2020: Perceptions of the challenges and opportunities of utilising organic waste through urban agriculture in the Durban South Basin. Int J Environ Res Public Health, 17 (4), doi:10.3390/ijerph17041158.
Mercure, J.F., et al., 2019: System complexity and policy integration challenges: the Brazilian energy-water-food nexus. Renew. Sustain. Energy Rev. , 105, 230–243, doi:10.1016/j.rser.2019.01.045.
Meredith, M., M. Sommerkorn, S. Cassotta, C. Derksen, A. Ekaykin, A. Hollowed, G. Kofinas, A. Mackintosh, J. Melbourne-Thomas, M.M.C. Muelbert, G. Ottersen, H. Pritchard, and E.A.G. Schuur, 2019: Polar Regions. In: IPCC Special Report on the Ocean and Crysosphere in a Changing Climate[Pörtner, H. O., D. C. Roberts, V. Vmasson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegría, M. Nicolai, A. Okem, J. Petzold, B. Rama and N. M. Weyer (eds.)], pp. 203–320. In press.
Meridian Institute, 2020: Food Systems of the Future: A Synthesis of Reports on Food Systems Transformation. Global Alliance for the Future of Food, https://futureoffood.org/wp-content/uploads/2020/03/GA-Meridian-Synthesis-Reports-FINAL-2020-03-16.pdf?fbclid=IwAR1QWVm77pNWD02MDZ6G2Jq3ayNw2LQyH0GuJLe46ZfXuCkirtCm37wJLaQ. Accessed 2021 . (31 pp).
Merlín-Uribe, Y., et al., 2012: Environmental and socio-economic sustainability of chinampas(raised beds) in Xochimilco, Mexico City. Int. J. Agric. Sustain. , 11 (3), 216–233, doi:10.1080/14735903.2012.726128.
Merloni, E., L. Camanzi, L. Mulazzani and G. Malorgio, 2018: Adaptive capacity to climate change in the wine industry: a Bayesian network approach. Wine Econ. Policy, 7 (2), 165–177, doi:10.1016/j.wep.2018.11.002.
Merow, C., et al., 2017: Climate change both facilitates and inhibits invasive plant ranges in New England. Proc. Natl. Acad. Sci. U.S.A. , 114 (16), E3276–E3284, doi:10.1073/pnas.1609633114.
Merrey, D.J., et al., 2018: Evolving high altitude livelihoods and climate change: a study from Rasuwa District, Nepal. Food Secur, 10 (4), 1055–1071, doi:10.1007/s12571-018-0827-y.
Mertz, O., A. Mbow, A. Reenberg and A. Diouf, 2009: Farmers’ perceptions of climate change and agricultural adaptation strategies in rural Sahel. Environ. Manag. , 45, 804–816.
Mertz, O., 2012: Climate variability and environmental stress in the Sudan-Sahel zone of West Africa. AMBIO, 41, 380–392.
Messer, E., 1972: Patterns of “wild” plant consumption in Oaxaca, Mexico. Ecol. Food. Nutr. , 1 (4), 325–332.
Metian, M., et al., 2020: Mapping diversity of species in global aquaculture. Rev. Aquac. , 12 (2), 1090–1100, doi:10.1111/raq.12374.
Metternicht, G., 2018: Land Use and Spatial Planning. SpringerBriefs in Earth Sciences. Springer International Publishing, Cham.
Meynard, C.N., M. Lecoq, M.P. Chapuis and C. Piou, 2020: On the relative role of climate change and management in the current desert locust outbreak in East Africa. Glob Chang Biol, 26 (7), 3753–3755, doi:10.1111/gcb.15137.
Meza, I., et al., 2020: Global-scale drought risk assessment for agricultural systems. Nat. Hazards Earth Syst. Sci. , 20 (2), 695–712, doi:10.5194/nhess-20-695-2020.
Michael, K., T. Deshpande and G. Ziervogel, 2019: Examining vulnerability in a dynamic urban setting: the case of Bangalore’s interstate migrant waste pickers. Clim. Dev. , 11 (8), 667–678, doi:10.1080/17565529.2018.1531745.
Mie, A., et al., 2017: Human health implications of organic food and organic agriculture: a comprehensive review. Environ. Health. , 16, 1–22, doi:10.1186/s12940-017-0315-4.
Mier y Terán Giménez Cacho, M., et al., 2018: Bringing agroecology to scale: key drivers and emblematic cases. Agroecol. Sustain. Food Syst. , 42 (6), 637–665, doi:10.1080/21683565.2018.1443313.
Migliorini, P., et al., 2016: Agronomic and quality characteristics of old, modern and mixture wheat varieties and landraces for organic bread chain in diverse environments of northern Italy. Eur J Agron, 79, 131–141, doi:10.1016/j.eja.2016.05.011.
Mikovits, C., et al., 2019: Impacts of global warming on confined livestock systems for growing-fattening pigs: simulation of heat stress for 1981 to 2017 in Central Europe. Int. J. Biometeorol. , 63 (2), 221–230, doi:10.1007/s00484-018-01655-0.
Mikulewicz, M., 2020: Disintegrating labour relations and depoliticised adaptation to climate change in rural São Tomé and Príncipe. Area, 53 (3), doi:10.1111/area.12630.
Milcu, A.I., J. Hanspach, D. Abson and J. Fischer, 2013: Cultural ecosystem services: a literature review and prospects for future research. Ecol. Soc. , 18 (3), 44, doi:10.5751/Es-05790-180344.
Milder, J.C., et al., 2014: Integrated landscape initiatives for African agriculture, development, and conservation: a region-wide assessment. World Dev, 54, 68–80, doi:10.1016/j.worlddev.2013.07.006.
Milicevic, D., et al., 2019: Impact of climate change on aflatoxin M1 contamination of raw milk with special focus on climate conditions in Serbia. J. Sci. Food Agric. , 99 (11), 5202–5210, doi:10.1002/jsfa.9768.
Millan, A., B. Limketkai and S. Guarnaschelli, 2019: Financing the Transformation of Food Systems Under a Changing Climate. CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS), Wageningen, the Netherlands, https://hdl.handle.net/10568/101132.
Millennium Ecosystem Assessment, 2005: Ecosystems and Human Well-being: Synthesis. Press Island, Washington, D.C., USA, ISBN 978-1597260404. 137 pp.
Miller-Struttmann, N.E., et al., 2015: Functional mismatch in a bumble bee pollination mutualism under climate change. Science, 349 (6255), 1541–1544, doi:10.1126/science.aab0868.
Miller, J.D., 2016: Mycotoxins in food and feed: a challenge for the twenty-first century. In: Biology of Microfungi[Li, D.W.(ed.)]. Springer, Cham, Switzerland, pp. 469–493. ISBN 978-3319291352.
Miller, K. A., G.R. Munro, U.R. Sumaila and W.W.L. Cheung, 2013: Governing marine fisheries in a changing climate: a game-theoretic perspective. Can. J. Agric. Econ. , 61 (2), 309–334, doi:10.1111/cjag.12011.
Millet, E.J., et al., 2016: Genome-wide analysis of yield in Europe: allelic effects vary with drought and heat scenarios. Plant Physiol. , 172 (2), 749–764, doi:10.1104/pp.16.00621.
Mills, E.N., 2018: Implicating “fisheries justice” movements in food and climate politics. Third World Q, 39 (7), 1270–1289, doi:10.1080/01436597.2017.1416288.
Mills, G., et al., 2018: Closing the global ozone yield gap: quantification and cobenefits for multistress tolerance. Glob Chang Biol, 24 (10), 4869–4893, doi:10.1111/gcb.14381.
Milner, S., et al., 2016: Potential impacts on ecosystem services of land use transitions to second-generation bioenergy crops in GB. Glob. Change Biol. Bioenergy, 8 (2), 317–333, doi:10.1111/gcbb.12263.
Minasny, B. and A.B. McBratney, 2018: Limited effect of organic matter on soil available water capacity. Eur J Soil Sci, 69, 39–47, doi:10.1111/ejss.12475.
Minet, J., et al., 2017: Crowdsourcing for agricultural applications: a review of uses and opportunities for a farmsourcing approach. Comput. Electron. Agric. , 142, 126–138, doi:10.1016/j.compag.2017.08.026.
Mingorría, S., 2018: Violence and visibility in oil palm and sugarcane conflicts: the case of Polochic Valley, Guatemala. J Peasant Stud, 45 (7), 1314–1340, doi:10.1080/03066150.2017.1293046.
Mingorría, S.G., B. Martín and E. Corbera, 2014: The oil palm boom: socio-economic implications for Q’eqchi’ households in the Polochic Valley, Guatemala. Environ. Dev. Sustain. , 16, 841–871, doi:10.1007/s10668-014-9530-0.
Minoli, S., et al., 2019: Global response patterns of major rainfed crops to adaptation by maintaining current growing periods and irrigation. Earths Future, 7 (12), 1464–1480, doi:10.1029/2018ef001130.
Mirás-Avalos, J.M., D. Uriarte, A.N. Lakso and D.S. Intrigliolo, 2018: Modeling grapevine performance with ‘VitiSim’, a weather-based carbon balance model: water status and climate change scenarios. Sci. Hortic. , 240, 561–571, doi:10.1016/j.scienta.2018.06.065.
Mistry, J. and A. Berardi, 2016: Bridging indigenous and scientific knowledge. Science, 352 (6291), 1274–1275, doi:10.1126/science.aaf1160.
Mitchell, D.C., et al., 2017: Recruitment, methods, and descriptive results of a physiologic assessment of Latino farmworkers: the California heat illness prevention study. J. Occup. Environ. Med. , 59 (7), 649–658, doi:10.1097/JOM.0000000000000988.
Miyan, M. A., 2015: Droughts in Asian least developed countries: vulnerability and sustainability. Weather. Clim. Extrem. , 7, 8–23, doi:10.1016/j.wace.2014.06.003.
Mohamad, N., et al., 2019a: Vibriosis in cultured marine fishes: a review. Aquaculture, 512, 734289, doi:10.1016/j.aquaculture.2019.734289.
Mohamad, N., et al., 2019b: Environmental factors associated with the presence of Vibrionaceae in tropical cage-cultured marine fishes. J Aquat Anim Health, 31 (2), 154–167, doi:10.1002/aah.10062.
Mohareb, E., et al., 2017: Considerations for reducing food system energy demand while scaling up urban agriculture. Environ. Res. Lett. , 12 (12), 125004, doi:10.1088/1748-9326/aa889b.
Mok, H.-F., et al., 2014: Strawberry fields forever? Urban agriculture in developed countries: a review. Agron. Sustain. Dev. , 34 (1), 21–43, doi:10.1007/s13593-013-0156-7.
Molitor, D. and J. Junk, 2019: Climate change is implicating a two-fold impact on air temperature increase in the ripening period under the conditions of the Luxembourgish grape growing region. Oeno One, 53 (3), 409–422, doi:10.20870/oeno-one.2019.53.3.2329.
Momblanch, A., et al., 2019: Untangling the water-food-energy-environment nexus for global change adaptation in a complex Himalayan water resource system. Sci. Total Environ. , 655, 35–47, doi:10.1016/j.scitotenv.2018.11.045.
Monsees, H., et al., 2019: Lettuce (Lactuca sativa, variety Salanova) production in decoupled aquaponic systems: same yield and similar quality as in conventional hydroponic systems but drastically reduced greenhouse gas emissions by saving inorganic fertilizer. PLoS ONE, 14 (6), e218368, doi:10.1371/journal.pone.0218368.
Montagnini, F. and R. Metzel, 2017: The contribution of agroforestry to Sustainable Development Goal 2: end hunger, achieve food security and improved nutrition, and promote sustainable agriculture. In: Integrating Landscapes: Agroforestry for Biodiversity Conservation and Food Sovereignty[Montagnini, F.(ed.)]. Springer, Cham, Switzerland, pp. 11–45. ISBN 978-3319693705.
Montanchez, I. and V.R. Kaberdin, 2020: Vibrio harveyi: a brief survey of general characteristics and recent epidemiological traits associated with climate change. Mar. Environ. Res. , 154, 104850, doi:10.1016/j.marenvres.2019.104850.
Montanchez, I., et al., 2019: Analysis of Vibrio harveyi adaptation in sea water microcosms at elevated temperature provides insights into the putative mechanisms of its persistence and spread in the time of global warming. Sci Rep, 9 (1), 289, doi:10.1038/s41598-018-36483-0.
Montefrio, M.J.F., 2017: Land control dynamics and social-ecological transformations in upland Philippines. J Peasant Stud, 44 (4), 770–790, doi:10.1080/03066150.2016.1257988.
Monteiro, A.P.A., S. Tao, I.M.T. Thompson and G.E. Dahl, 2016: In utero heat stress decreases calf survival and performance through the first lactation. J. Dairy Sci. , 99 (10), 8443–8450, doi:10.3168/jds.2016-11072.
Montenegro de Wit, M., 2019: Gene driving the farm: who decides, who owns, and who benefits?Agroecol. Sustain. Food Syst. , 43 (9), 1054–1074, doi:10.1080/21683565.2019.1591566.
Montenegro de Wit, M., A.R. Kapuscinski and E. Fitting, 2020: Democratizing CRISPR? Stories, practices, and politics of science and governance on the agricultural gene editing frontier. Elem. Sci. Anthropocene, 8, doi:10.1525/elementa.405.
Moor, M. A., et al., 2017: Individual and community factors contributing to anemia among women in rural Baja California, Mexico. Plos One, 12 (11), e188590, doi:10.1371/journal.pone.0188590.
Moore, A.D. and A. Ghahramani, 2014: Climate change and broadacre livestock production across southern Australia. 3. Adaptation options via livestock genetic improvement. Anim. Prod. Sci. , 54 (2), 111–124, doi:10.1071/An13052.
Moore, F., 2020: The fingerprint of anthropogenic warming on global agriculture. EarthArXiv, doi:10.31223/x5q30z.
Moore, F.C. and D.B. Lobell, 2015: The fingerprint of climate trends on European crop yields. Proc. Natl. Acad. Sci. U. S. A. , 112 (9), 2670–2675, doi:10.1073/pnas.1409606112.
Moore, S.E. and H.P. Huntington, 2008: Arctic marine mammals and climate change: impacts and resilience. Ecol Appl, 18 (2 Suppl), s157–165, doi:10.1890/06-0571.1.
Morales-Castilla, I., et al., 2020: Diversity buffers winegrowing regions from climate change losses. Proc. Natl. Acad. Sci. U. S. A. , 117 (6), 2864–2869, doi:10.1073/pnas.1906731117.
Morales, F., et al., 2014: Methodological advances: using greenhouses to simulate climate change scenarios. Plant Sci. , 226, 30–40, doi:10.1016/j.plantsci.2014.03.018.
Moreda, T., 2017: Large-scale land acquisitions, state authority and indigenous local communities: insights from Ethiopia. Third World Q, 38 (3), 698–716, doi:10.1080/01436597.2016.1191941.
Moreda, T., 2018: The right to food in the context of large-scale land investment in Ethiopia. Third World Q, 39 (7), 1326–1347, doi:10.1080/01436597.2018.1460199.
Moretti, A., M. Pascale and A. F. Logrieco, 2019: Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. , 84, 38–40, doi:10.1016/j.tifs.2018.03.008.
Morgan, M., et al., 2015: Understanding the Gender Dimensions of Adopting Climate-Smart Smallholder Aquaculture Innovations. Working Paper, CGIAR Research Program on Aquatic Agricultural Systems, Penang, Malaysia, <Go to WoS>://WOS:000361906400033. Accessed 2021 . (43 pp).
Moriondo, M., et al., 2015: Modelling olive trees and grapevines in a changing climate. Environ. Model. Softw. , 72, 387–401, doi:10.1016/j.envsoft.2014.12.016.
Morita, S., H. Wada and Y. Matsue, 2016: Countermeasures for heat damage in rice grain quality under climate change. Plant. Prod. Sci. , 19 (1), 1–11, doi:10.1080/1343943x.2015.1128114.
Morrissey, M.T., R. Rasmussen and T. Okada, 2005: Mercury Content in Pacific Troll-Caught Albacore Tuna ( Thunnus alalunga ). J. Aquat. Food Prod. Technol. , 13 (4), 41–52, doi:10.1300/J030v13n04_04.
Mortimore, M., 2010: Adapting to drought in the Sahel: lessons for climate change. Wiley. Interdiscip. Rev. Clim. Change. , 1 (1), 134–143, doi:10.1002/wcc.25.
Mosberg, M. and S.H. Eriksen, 2015: Responding to climate variability and change in dryland Kenya: the role of illicit coping strategies in the politics of adaptation. Glob. Environ. Change. , 35, 545–557, doi:10.1016/j.gloenvcha.2015.09.006.
Mosedale, J.R., et al., 2016: Climate change impacts and adaptive strategies: lessons from the grapevine. Glob Chang Biol, 22 (11), 3814–3828, doi:10.1111/gcb.13406.
Mosedale, J.R., R.J. Wilson and I.M. Maclean, 2015: Climate change and crop exposure to adverse weather: changes to frost risk and grapevine flowering conditions. PLoS ONE, 10 (10), e141218, doi:10.1371/journal.pone.0141218.
Moseley, W., M. Schnurr and R.B. Kerr, 2015: Interrogating the technocratic (neoliberal) agenda for agricultural development and hunger alleviation in Africa. Afr. Geogr. Rev. , 34 (1), 1–7, doi:10.1080/19376812.2014.1003308.
Moses, J.A., D.S. Jayas and K. Alagusundaram, 2015: Climate change and its implications on stored food grains. Agric. Res. , 4 (1), 21–30, doi:10.1007/s40003-015-0152-z.
Mosnier, A., et al., 2014: Global food markets, trade and the cost of climate change adaptation. Food Secur, 6 (1), 29–44, doi:10.1007/s12571-013-0319-z.
Mottet, A., et al., 2020: Assessing Transitions to Sustainable Agricultural and Food Systems: A Tool for Agroecology Performance Evaluation (TAPE). Front. Sustain. Food Syst. , 4, doi:10.3389/fsufs.2020.579154.
Moukrim, S., et al., 2019: Climate change impacts on potential distribution of multipurpose agro-forestry species: Argania spinosa (L.) Skeels as case study. Agrofor. Syst. , 93 (4), 1209–1219.
Moulton, A.A. and M.R. Machado, 2019: Bouncing Forward After Irma and Maria: Acknowledging Colonialism, Problematizing Resilience and Thinking Climate Justice. J. Extrem. Events, 06 (01), doi:10.1142/s2345737619400037.
Mousseau, F., 2015: The untold success story of agroecology in Africa. Development , 58, 341–345, doi:10.1057/s41301-016-0026-0.
Mousseau, F. and E. Teare, 2019: Evicted for Carbon Credits: Norway, Sweden, and Finland Displace Ugandan Farmers for Carbon Trading. The Oakland Institute, Oakland, CA, https://www.oaklandinstitute.org/evicted-carbon-credits-green-resources. Accessed 2021 . (15 pp).
Mpandeli, S., et al., 2018: Climate change adaptation through the water-energy-food nexus in Southern Africa. Int J Environ Res Public Health, 15 (10), 2306, doi:10.3390/ijerph15102306.
Mrabet, R., et al., 2020: Food. In: Climate and Environmental Change in the Mediterranean Basin- Current Situation and Risks fo the Future. First Mediterranean Assessment Report [Cramer, W., J. Guiot and K. Marini(eds.)]. Union for the Mediterranean, Plan Bleu, UNEP/MAP, Marseille, France, pp. 237–264.
Msuya, F.E. and A.Q. Hurtado, 2017: The role of women in seaweed aquaculture in the Western Indian Ocean and South-East Asia. Eur. J. Phycol. , 52 (4), 482–494, doi:10.1080/09670262.2017.1357084.
Mu, J.E., J.M. Antle and J.T. Abatzoglou, 2019: Representative agricultural pathways, climate change, and agricultural land uses: an application to the Pacific Northwest of the USA. Mitig Adapt Strateg Glob Change, 24 (5), 819–837, doi:10.1007/s11027-018-9834-8.
Muchane, M.N., et al., 2020: Agroforestry boosts soil health in the humid and sub-humid tropics: a meta-analysis. Agric Ecosyst Environ, 295, 106899, doi:10.1016/j.agee.2020.106899.
Muchuru, S. and G. Nhamo, 2019: A review of climate change adaptation measures in the African crop sector. Clim. Dev. , 11 (10), 873–885, doi:10.1080/17565529.2019.1585319.
Muehe, E.M., et al., 2019: Rice production threatened by coupled stresses of climate and soil arsenic. Nat Commun, 10 (1), 4985, doi:10.1038/s41467-019-12946-4.
Mueller, B., et al., 2015: Lengthening of the growing season in wheat and maize producing regions. Weather. Clim. Extrem. , 9, 47–56, doi:10.1016/j.wace.2015.04.001.
Muema, E., J. Mburu, J. Coulibaly and J. Mutune, 2018: Determinants of access and utilisation of seasonal climate information services among smallholder farmers in Makueni County, Kenya. Heliyon, 4 (11), e889, doi:10.1016/j.heliyon.2018.e00889.
Mukhopadhyay, R. and S.B. Roy, 2017: Traditional knowledge for biodiversity conservation, maintain ecosystem services and livelihood security in the context of climate change: case studies from West Bengal, India. J. Biodivers. , 6, 22–29, doi:10.1080/09766901.2015.11884752.
Mul, M., et al., 2015: Water Resources Assessment of the Volta River Basin. IWMI Working Paper, International Water Management Institute, Colombo, Sri Lanka, ISBN 978-9290908296.
Muller, A., et al., 2017: Strategies for feeding the world more sustainably with organic agriculture. Nat Commun, 8 (1), 1290, doi:10.1038/s41467-017-01410-w.
Mulwa, C.K. and M. Visser, 2020: Farm diversification as an adaptation strategy to climatic shocks and implications for food security in northern Namibia. World Dev, 129, 104906, doi:10.1016/j.worlddev.2020.104906.
Munt, D.D., P. Munoz-Rodriguez, I. Marques and J.C.M. Saiz, 2016: Effects of climate change on threatened Spanish medicinal and aromatic species: predicting future trends and defining conservation guidelines. Isr. J. Plant Sci. , 63 (4), 309–319, doi:10.1080/07929978.2016.1258257.
Mupambwa, H.A., M.K. Hausiku, A.D. Nciizah and E. Dube, 2019: The unique Namib desert-coastal region and its opportunities for climate smart agriculture: a review. Cogent Food Agric, 5 (1), 1645258, doi:10.1080/23311932.2019.1645258.
Murakami, H., G. Vecchi and S. Underwood, 2017: Increasing frequency of extremely severe cyclonic storms over the Arabian Sea. Nat. Clim. Chang. , 7, 885–889, doi:10.1038/s41558-017-0008-6.
Muringai, R.T., P.L. Mafongoya and R. Lottering, 2021: Climate Change and Variability Impacts on Sub-Saharan African Fisheries: A Review. Rev. Fish. Sci. Aquac. , 1–21, doi:10.1080/23308249.2020.1867057.
Muscat, A., E.M. de Olde, I.J.M. de Boer and R. Ripoll-Bosch, 2020: The battle for biomass: A systematic review of food-feed-fuel competition. Glob. Food Secur. , 25, 100330–100330, doi:10.1016/j.gfs.2019.100330.
Muscat, A., et al., 2021: Food, energy or biomaterials? Policy coherence across agro-food and bioeconomy policy domains in the EU. Environ Sci Policy, 123, 21–30, doi:10.1016/J.ENVSCI.2021.05.001.
Musinguzi, L., et al., 2016: Fishers’ perceptions of climate change, impacts on their livelihoods and adaptation strategies in environmental change hotspots: A case of Lake Wamala, Uganda. Environ. Dev. Sustain. , 18 (4), 1255–1273.
Musinguzi, L., V. Natugonza, J. Efitre and R. Ogutu-Ohwayo, 2018: The role of gender in improving adaptation to climate change among small-scale fishers. Clim. Dev. , 10 (6), 566–576, doi:10.1080/17565529.2017.1372262.
Muzhinji, N. and V. Ntuli, 2021: Genetically modified organisms and food security in Southern Africa: conundrum and discourse. GM Crops Food. , 12 (1), 25–35, doi:10.1080/21645698.2020.1794489.
Mweya, C.N., L.E. G. Mboera and S.I. Kimera, 2017: Climate Influence on emerging risk areas for Rift Valley fever epidemics in Tanzania. Am. J. Trop. Med. Hyg. , 97 (1), 109–114, doi:10.4269/ajtmh.16-0444.
Myers, B.J.E., et al., 2017a: Global synthesis of the documented and projected effects of climate change on inland fishes. Rev. Fish Biol. Fish. , 27 (2), 339–361, doi:10.1007/s11160-017-9476-z.
Myers, S.S., et al., 2017b: Climate change and global food systems: potential impacts on food security and undernutrition. Annu. Rev. Public Health, 38 (1), 259–277, doi:10.1146/annurev-publhealth-031816-044356.
Myers, S.S., et al., 2014: Increasing CO2 threatens human nutrition. Nature, 510 (7503), 139–142, doi:10.1038/nature13179.
Naah, J.S.N. and B. Braun, 2019: Local agro-pastoralists’ perspectives on forage species diversity, habitat distributions, abundance trends and ecological drivers for sustainable livestock production in West Africa. Sci Rep, 9 (1), 1707, doi:10.1038/s41598-019-38636-1.
Nabuurs, G.-J., et al., 2018: Climate-Smart Forestry: Mitigation Impacts in Three European Regions. From Science to Policy, Vol. 6. European Forest Institute, ISBN 978-9525980530.
Nadeau, A., G. Cloutier, C. Poitras and A. Aylett, 2019: Racines citoyennes: la communauté locale au coeur de la transition écologique l’impact des initiatives climatiques locales et citoyennes à Montréal. Can. J. Urban. Res. , 28 (2), 16–31.
Nagahama, N. and M.F. Bonino, 2020: Modeling the potential distribution of Valeriana carnosa Sm. in Argentinean Patagonia: a proposal for conservation and in situ cultivation considering climate change projections. J. Appl. Res. Med. Aromat. Plants, 16, 100240, doi:10.1016/j.jarmap.2020.100240.
Naghipour Borj, A.A., Z. Ostovar and E. Asadi, 2019: The influence of climate change on distribution of an endangered medicinal plant (Fritillaria Imperialis L.) in Central Zagros. J. Rangel. Sci. , 9 (2), 159–171.
Nagoda, S., 2015: New discourses but same old development approaches? Climate change adaptation policies, chronic food insecurity and development interventions in northwestern Nepal. Glob. Environ. Chang. , 35, 570–579, doi:10.1016/j.gloenvcha.2015.08.014.
Nagoda, S. and A.J. Nightingale, 2017: Participation and Power in Climate Change Adaptation Policies: Vulnerability in Food Security Programs in Nepal. World Dev, 100, 85–93, doi:10.1016/j.worlddev.2017.07.022.
Naicker, P.R., 2011: The impact of climate change and other factors on zoonotic diseases. Arch. Clin. Microbiol. , 2 (6), doi:10:3823/226.
Najafi, E., I. Pal and R. Khanbilvardi, 2019: Climate drives variability and joint variability of global crop yields. Sci. Total Environ. , 662, 361–372, doi:10.1016/j.scitotenv.2019.01.172.
Nakashima, D.J., et al., 2012: Weathering Uncertainty: Traditional Knowledge for Climate Change Assessment and Adaptation. United Nations Educational Scientific and Cultural Organization and United Nations University- Traditional Knowledge Initiative, Paris, France and Darwin, Australia, ISBN 978-9230010683. 120 pp.
Nalau, J., S. Becken and B. Mackey, 2018: Ecosystem-based adaptation: a review of the constraints. Environ Sci Policy, 89, 357–364, doi:10.1016/j.envsci.2018.08.014.
Namgay, K., J.E. Millar, R.S. Black and T. Samdup, 2014: Changes in transhumant agro-pastoralism in Bhutan: a disappearing livelihood?Hum Ecol, 42 (5), 779–792, doi:10.1007/s10745-014-9684-2.
Nandal, U. and R.L. Bhardwaj, 2014: The role of underutilized fruits in nutritional and economic security of tribals: a review. Crit. Rev. Food Sci. Nutr. , 54 (7), 880–890, doi:10.1080/10408398.2011.616638.
Nandintsetseg, B., et al., 2021: Risk and vulnerability of Mongolian grasslands under climate change. Environ. Res. Lett. , 16 (3), 34035–34035, doi:10.1088/1748-9326/abdb5b.
Napier, J.A., R.P. Haslam, M. Tsalavouta and O. Sayanova, 2019: The challenges of delivering genetically modified crops with nutritional enhancement traits. Nat. Plants, 5 (6), 563–567, doi:10.1038/s41477-019-0430-z.
Naqvi, S.M.K., K. De, D. Kumar and V. Sejian, 2017: Climate changes, water use and survival during severe water deprivation. In: Sheep Production Adapting to Climate Change[Sejian, V., et al.(ed.)]. Springer Singapore, Singapore, pp. 173–187. ISBN 978-9811047138.
Naqvi, S.M.K., D. Kumar, K. De and V. Sejian, 2015: Climate change and water availability for livestock: impact on both quality and quantity. In: Climate Change Impact on Livestock: Adaptation and Mitigation[Sejian, V., J. Gaughan, L. Baumgard and C. Prasad(eds.)]. Springer India, New Delhi, pp. 81–95.
Narayan, S., et al., 2020: Local adaptation responses to coastal hazards in small island communities: insights from 4 Pacific nations. Environ Sci Policy, 104, 199–207, doi:10.1016/j.envsci.2019.11.006.
Nardone, A., B. Ronchi, N. Lacetera and U. Bernabucci, 2006: Climatic Effects on Productive Traits in Livestock. Vet. Res. Commun. , 30 (S1), 75–81, doi:10.1007/s11259-006-0016-x.
Nasi, R., A. Taber and N. Van Vliet, 2011: Empty forests, empty stomachs? Bushmeat and livelihoods in the Congo and Amazon Basins. Int. For. Rev. , 13 (3), 355–368, doi:10.1505/146554811798293872.
Nasielski, J., et al., 2015: Agroforestry promotes soybean yield stability and N2-fixation under water stress. Agron. Sustain. Dev. , 35 (4), 1541–1549, doi:10.1007/s13593-015-0330-1.
Naskar, M., et al., 2018: Quantifying climate change induced threats to wetland fisheries: a stakeholder-driven approach. Environ. Dev. Sustain. , 20 (6), 2811–2830, doi:10.1007/s10668-017-0018-6.
Nasonova, O.N., Y.M. Gusev, E.M. Volodin and E.E. Kovalev, 2018: Application of the land surface model SWAP and global climate model INMCM4.0 for projecting runoff of northern Russian rivers. 1. Historical simulations. Water Resour. , 45, 73–S84, doi:10.1134/S009780781806026x.
Nath, A.J., R. Lal, G.W. Sileshi and A.K. Das, 2018: Managing India’s small landholder farms for food security and achieving the “4 per Thousand” target. Sci. Total Environ. , 634, 1024–1033, doi:10.1016/j.scitotenv.2018.03.382.
Nath, A.J., et al., 2021: Quantifying carbon stocks and sequestration potential in agroforestry systems under divergent management scenarios relevant to India’s Nationally Determined Contribution. J Clean Prod, 281, doi:10.1016/j.jclepro.2020.124831.
Nath, T.K., et al., 2016: The sustainable intensification of agroforestry in shifting cultivation areas of Bangladesh. Agroforest Syst , 90, 405–416, doi:10.1007/s10457-015-9863-1.
National Academies of Sciences Engineering and Medicine, 2016: Genetically Engineered Crops: Experiences and Prospects. National Academies Press, Washington, D.C., ISBN 978-0309437387. 420 pp.
Nature climate change Editorials, 2019: Gender in conservation and climate policy. Nat. Clim. Chang. , 9 (4), 255–255, doi:10.1038/s41558-019-0448-2.
Naughton-Treves, L. and K. Wendland, 2014: Land tenure and tropical forest carbon management. World Dev, 55, 1–6, doi:10.1016/j.worlddev.2013.01.010.
Naylor, R.L., et al., 2021: A 20-year retrospective review of global aquaculture. Nature, 591 (7851), 551–563, doi:10.1038/s41586-021-03308-6.
Ndhlovu, N., O. Saito, R. Djalante and N. Yagi, 2017: Assessing the sensitivity of small-scale fishery groups to climate change in Lake Kariba, Zimbabwe. Sustainability, 9 (12), 2209, doi:10.3390/su9122209.
Ndi, F.A., 2019: Land grabbing: A gendered understanding of perceptions and reactions from affected communities in Nguti Subdivision of South West Cameroon. Dev. Policy. Rev. , 37 (3), 348–366, doi:10.1111/dpr.12351.
Neale, T., R. Carter, T. Nelson and M. Bourke, 2019: Walking together: a decolonising experiment in bushfire management on Dja Dja Wurrung country. Cult. Geogr. , 26 (3), 341–359, doi:10.1177/1474474018821419.
Neethling, E., G. Barbeau, C. Coulon-leroy and H. Quénol, 2019: Agricultural and Forest Meteorology Spatial complexity and temporal dynamics in viticulture : A review of climate-driven scales. Agric For Meteorol, 276-277 (June), 107618–107618, doi:10.1016/j.agrformet.2019.107618.
Negin, B. and M. Moshelion, 2017: The advantages of functional phenotyping in pre-field screening for drought-tolerant crops. Funct. Plant. Biol. , 44, 107–118, doi:10.1071/FP16156.
Negra, C., et al., 2020: Sustainable agri-food investments require multi-sector co-development of decision tools. Ecol Indic, 110, 105851, doi:10.1016/j.ecolind.2019.105851.
Nehring, R., A. Miranda and A. Howe, 2017: Making the case for institutional demand: supporting smallholders through procurement and food assistance programmes. Glob. Food Secur. Policy Econ. Environ. , 12, 96–102, doi:10.1016/j.gfs.2016.09.003.
Nelson, G., et al., 2018: Income growth and climate change effects on global nutrition security to mid-century. Nat. Sustain. , 1 (12), 773–781, doi:10.1038/s41893-018-0192-z.
Nelson, G.C., et al., 2014: Climate change effects on agriculture: economic responses to biophysical shocks. Proc. Natl. Acad. Sci. U. S. A. , 111 (9), 3274–3279, doi:10.1073/pnas.1222465110.
Neset, T.S., T. Asplund, J. Kayhko and S. Juhola, 2019a: Making sense of maladaptation: Nordic agriculture stakeholders’ perspectives. Clim Change, 153 (1-2), 107–121, doi:10.1007/s10584-019-02391-z.
Neset, T.S., et al., 2019b: Maladaptation in Nordic agriculture. Clim. Risk Manag. , 23, 78–87, doi:10.1016/j.crm.2018.12.003.
Neumann, R.B., A.L. Seyfferth, J. Teshera-Levye and J. Ellingson, 2017: Soil Warming Increases Arsenic Availability in the Rice Rhizosphere. Agric. Environ. Lett. , 2 (1), doi:10.2134/ael2017.02.0006.
Neuner, S., et al., 2015: Survival of Norway spruce remains higher in mixed stands under a dryer and warmer climate. Glob Chang Biol, 21 (2), 935–946, doi:10.1111/gcb.12751.
Neven, L.G., 2000: Physiological responses of insects to heat. Postharvest Biol. Technol. , 21 (1), 103–111, doi:10.1016/S0925-5214(00)00169-1.
Newbold, T., et al., 2015: Global effects of land use on local terrestrial biodiversity. Nature, 520 (7545), 45–50, doi:10.1038/nature14324.
Newton, P., et al., 2020: The number and spatial distribution of forest-proximate people globally. One Earth, 3 (3), 363–370.
NGFS, 2020: Overview of Environmental Risk Analysis by Financial Institutions. Network for Greening the Financial System / Banque de France, https://www.ngfs.net/en/communique-de-presse/ngfs-promotes-environmental-risk-analysis-financial-industry.
Ngigi, M.W., U. Mueller and R. Birner, 2017: Gender differences in climate change adaptation strategies and participation in group-based approaches: An Intra-household analysis from rural Kenya. Ecol. Econ. , 138, 99–108, doi:10.1016/j.ecolecon.2017.03.019.
Nguyen, H.Q., et al., 2019a: Farmer adoptability for livelihood transformations in the Mekong Delta: a case in Ben Tre province. J. Environ. Plan. Manag. , 62 (9), 1603–1618, doi:10.1080/09640568.2019.1568768.
Nguyen, L.A., et al., 2018: Impact of Climate Change on the Technical Efficiency of Striped Catfish, Pangasianodon hypophthalmus, Farming in the Mekong Delta, Vietnam. J. World Aquacult. Soc. , 49 (3), 570–581, doi:10.1111/jwas.12488.
Nguyen, L.A., et al., 2016: A decision tree analysis to support potential climate change adaptations of striped catfish farming in the Mekong Delta, Vietnam. Tropicultura, 34, 105–115.
Nguyen, Q., M.H. Hoang, I. Oborn and M. van Noordwijk, 2013: Multipurpose agroforestry as a climate change resiliency option for farmers: an example of local adaptation in Vietnam. Clim Change, 117 (1-2), 241–257, doi:10.1007/s10584-012-0550-1.
Nguyen, T.A.T., K.A.T. Nguyen and C. Jolly, 2019b: Is Super-Intensification the Solution to Shrimp Production and Export Sustainability?Sustainability, 11 (19), doi:10.3390/su11195277.
Nguyen, T.P. and K.E. Parnell, 2019: Coastal land use planning in Ben Tre, Vietnam: constraints and recommendations. Heliyon, 5 (4), e1487, doi:10.1016/j.heliyon.2019.e01487.
Nhamo, L., et al., 2018: The Water-Energy-Food Nexus: Climate Risks and Opportunities in Southern Africa. Water, 10 (5), 567, doi:10.3390/w10050567.
Nhat Lam Duyen, T., 2021: A comparative analysis of gender and youth issues in rice production in North, Central, and South Vietnam. Clim. Dev. , 13 (2), 115–127, doi:10.1080/17565529.2020.1734771.
Nhung, T.T., P.V. Vo, V.V. Nghi and H.Q. Bang, 2019: Salt intrusion adaptation measures for sustainable agricultural development under climate change effects: A case of Ca Mau Peninsula, Vietnam. Clim. Risk Manag. , 23, 88–100, doi:10.1016/j.crm.2018.12.002.
Nicholson, C.C. and P.A. Egan, 2020: Natural hazard threats to pollinators and pollination. Glob Chang Biol, 26 (2), 380–391, doi:10.1111/gcb.14840.
Nidumolu, U., et al., 2020: Linking climate forecasts to rural livelihoods: Mapping decisions, information networks and value chains. Weather. Clim. Extrem. , 27, doi:10.1016/j.wace.2018.06.001.
Nie, Z.N., et al., 2016: Benefits, challenges and opportunities of integrated crop-livestock systems and their potential application in the high rainfall zone of southern Australia: A review. Agric Ecosyst Environ, 235, 17–31, doi:10.1016/j.agee.2016.10.002.
Niederle, P., A. Loconto, S. Lemeilleur and C. Dorville, 2020: Social movements and institutional change in organic food markets: Evidence from participatory guarantee systems in Brazil and France. J Rural Stud, 78, 282–291, doi:10.1016/j.jrurstud.2020.06.011.
Nielsen, J.O. and A. Reenberg, 2010: Cultural barriers to climate change adaptation: A case study from Northern Burkina Faso. Glob. Environ. Change. , 20 (1), 142–152, doi:10.1016/j.gloenvcha.2009.10.002.
Niether, W., et al., 2020: Cocoa agroforestry systems versus monocultures: a multi-dimensional meta-analysis. Environ. Res. Lett. , 15 (10), doi:10.1088/1748-9326/abb053.
Niether, W., et al., 2017: Spatial-temporal soil moisture dynamics under different cocoa production systems. CATENA, 158, 340–349, doi:10.1016/j.catena.2017.07.011.
Nightingale, A.J., 2017: Power and politics in climate change adaptation efforts: Struggles over authority and recognition in the context of political instability. Geoforum, 84, 11–20, doi:10.1016/j.geoforum.2017.05.011.
Nightingale, A.J., et al., 2020: Beyond Technical Fixes: climate solutions and the great derangement. Clim. Dev. , 12 (4), 343–352, doi:10.1080/17565529.2019.1624495.
Niles, M.T. and M. E. Brown, 2017: A multi-country assessment of factors related to smallholder food security in varying rainfall conditions. Sci Rep, 7 (1), 16277, doi:10.1038/s41598-017-16282-9.
Niles, M.T., et al., 2021: Climate impacts associated with reduced diet diversity in children across nineteen countries. Environ. Res. Lett. , 16 (1), 15010, doi:10.1088/1748-9326/abd0ab.
Nkiaka, E., N.R. Nawaz and J.C. Lovett, 2017: Evaluating Global Reanalysis Datasets as Input for Hydrological Modelling in the Sudano-Sahel Region. Hydrology, 4 (1), doi:10.3390/hydrology4010013.
Nkiaka, E., et al., 2019: Identifying user needs for weather and climate services to enhance resilience to climate shocks in sub-Saharan Africa. Environ. Res. Lett. , 14 (12), doi:10.1088/1748-9326/ab4dfe.
Nkuba, M.R., et al., 2020: Indigenous Knowledge Systems and Indicators of Rain: Evidence from Rwenzori Region, Western Uganda. Weather Clim. Soc. , 12 (2), 213–234, doi:10.1175/Wcas-D-19-0027.1.
Nolte, K. and M. Ostermeier, 2017: Labour Market Effects of Large-Scale Agricultural Investment: Conceptual Considerations and Estimated Employment Effects. World Dev, 98 (2016), 430–446, doi:10.1016/j.worlddev.2017.05.012.
Noongwook, G., H.P. Huntington, J.C. George and N.V. Savoonga, 2007: Traditional knowledge of the bowhead whale (Balaena mysticetus) around St. Lawrence Island, Alaska. Arctic, 60 (1), 47–54.
Norton-Smith, K., et al., 2016: Climate Change and Indigenous Peoples : A Synthesis of Current Impacts and Experiences, 1–142.
Nost, E., 2019: Climate services for whom? The political economics of contextualizing climate data in Louisiana’s coastal Master Plan. Clim Change, 157 (1), 27–42, doi:10.1007/s10584-019-02383-z.
Novikova, A., L. Rocchi and V. Vitunskienė, 2017: Assessing the benefit of the agroecosystem services: Lithuanian preferences using a latent class approach. Land Use Policy, 68, 277–286, doi:10.1016/j.landusepol.2017.07.051.
Noya, I., et al., 2019: Water Footprint of Livestock Farming. In: Environmental Water Footprints, [Muthu, S. S. (ed.)]. Springer, Singapore, pp. 21-54. ISBN 978-981-13-2507-6..
Nuccio, M.L., et al., 2018: Where are the drought tolerant crops? An assessment of more than two decades of plant biotechnology effort in crop improvement. Plant Sci. , 273, 110–119, doi:10.1016/j.plantsci.2018.01.020.
Nunes, P.A.L.D. and M.L. Loureiro, 2016: Economic valuation of climate-change-induced vinery landscape impacts on tourism flows in Tuscany. Agric. Econ. , 47 (4), 365–374, doi:10.1111/agec.12236.
Nunn, P.D., et al., 2020: Adaptation to Climate Change: Contemporary Challenges and Perspectives. In: Climate Change and Impacts in the Pacific[Kumar, L.(ed.)]. Springer International Publishing, Cham, pp. 499–524. ISBN 978-3030328771.
Nunoo, F.K.E., et al., 2014: Marine Fisheries Catches in Ghana: Historic Reconstruction for 1950 to 2010 and Current Economic Impacts. Rev. Fish. Sci. Aquac. , 22 (4), 274–283, doi:10.1080/23308249.2014.962687.
Nuttall, J.G., et al., 2017: Models of grain quality in wheat-A review. Field Crop. Res. , 202, 136–145, doi:10.1016/j.fcr.2015.12.011.
Nuttall, M., 2009: Living in a world of movement: human resilience to environmental instability in Greenland. In: Anthropology and climate change: from encounters to actions[Crate, S.A. and M. Nuttall(eds.)]. Routledge, Oxon, England, pp. 292–310.
Nuttall, M., 2017: Climate, Society and Subsurface Politics in Greenland: Under the Great Ice. Taylor & Francis.
Nyamushamba, G.B., et al., 2017: Conservation of indigenous cattle genetic resources in Southern Africa’s smallholder areas: turning threats into opportunities - A review. Asian-australas J Anim Sci, 30 (5), 603–621, doi:10.5713/ajas.16.0024.
Nyangiwe, N., M. Yawa and V. Muchenje, 2018: Driving forces for changes in geographic range of cattle ticks (Acari: Ixodidae) in Africa: A review. S Afr J Anim Sci, 48 (5), 829–841, doi:10.4314/sajas.v48i5.4.
Nyantakyi-Frimpong, H., 2019: Combining feminist political ecology and participatory diagramming to study climate information service delivery and knowledge flows among smallholder farmers in northern Ghana. Appl. Geogr. , 112, doi:10.1016/j.apgeog.2019.102079.
Nyantakyi-Frimpong, H., 2020a: Unmasking difference: intersectionality and smallholder farmers’ vulnerability to climate extremes in Northern Ghana. Gend. Place Cult. , 27 (11), 1536–1554, doi:10.1080/0966369X.2019.1693344.
Nyantakyi-Frimpong, H., 2020b: What lies beneath: Climate change, land expropriation, and zaï agroecological innovations by smallholder farmers in Northern Ghana. Land Use Policy, 92 (December 2019), 104469–104469, doi:10.1016/j.landusepol.2020.104469.
Nyantakyi-Frimpong, H. and R. Bezner-Kerr, 2015: The relative importance of climate change in the context of multiple stressors in semi-arid Ghana. Glob. Environ. Change. , 32, 40–56, doi:10.1016/j.gloenvcha.2015.03.003.
Nyantakyi-Frimpong, H. and R. Bezner Kerr, 2017: Land grabbing, social differentiation, intensified migration and food security in northern Ghana. J Peasant Stud, 44 (2), 421–444, doi:10.1080/03066150.2016.1228629.
Nyberg, Y., J. Wetterlind, M. Jonsson and I. Oborn, 2020: The role of trees and livestock in ecosystem service provision and farm priorities on smallholder farms in the Rift Valley, Kenya. Agric Syst , 181, 102815–102815, doi:10.1016/j.agsy.2020.102815.
Nyguyen, K.A.T. and T. Pongthanapanic, 2016: Aquaculture Insurance in Viet Nam: Experiences from the pilot program. FAO Fisheries and Aquaculture Circular, Vol. 1133. FAO, Rome. 20 pp.
Nyland, K.E., et al., 2017: Traditional Inupiat Ice Cellars (Sigluaq) in Barrow, Alaska: Characteristics, Temperature Monitoring, and Distribution. Geogr Rev, 107 (1), 143–158, doi:10.1111/j.1931-0846.2016.12204.x.
Nyong, A., F. Adesina and B. Osman Elasha, 2007: The value of Indigenous knowledge in climate change mitigation and adaptation strategies in the African Sahel. Mitig Adapt Strateg Glob Change, 12, 787–797.
Nyoni, N.M.B., S. Grab and E.R.M. Archer, 2019: Heat stress and chickens: climate risk effects on rural poultry farming in low-income countries. Clim. Dev. , 11 (1), 83–90, doi:10.1080/17565529.2018.1442792.
O’Corry-Crowe, G. M., 2009: Beluga whale: Delphinapterus leucas. In: Encyclopedia of Marine Mammals (Second Edition) [Perrin, W. F., B. Würsig and J. G. M. Thewissen (eds.)]. Academic Press, UK, pp. 108-112. ISBN 9780123735539.
O’Sullivan, C.A., et al., 2019: Strategies to improve the productivity, product diversity and profitability of urban agriculture. Agric Syst , 174 (December 2018), 133–144, doi:10.1016/j.agsy.2019.05.007.
O’Brien, K., 2018: Is the 1.5 C target possible? Exploring the three spheres of transformation. Curr. Opin. Environ. Sustain. , 31, 153–160.
O’Brien, K., S.E.H. Eriksen, T.H. Inderberg and L. Sygna, 2015: Climate change and development: adaptation through transformation. In: Climate Change Adaptation and Development: Transforming Paradigms and Practices[Inderberg, T.H., S.E.H. Eriksen, K. O’Brien and L. Sygna(eds.)]. Routledge, New York, NY, pp. 273–289.
O’Connell-Milne, S.A. and C. Hepburn, 2015: A harvest method informed by traditional knowledge maximises yield and regeneration post harvest for karengo (Bangiaceae). J Appl Phycol, 27, 447–454.
Oates, L., R. Gillard, A. Sudmant and A. Gouldson, 2020: Secure and equal access to land for all: Lessons on land governance and climate resilience from Dar es Salaam, Tanzania. University of Leeds, Leeds.
Oberlack, C., et al., 2016: Sustainable livelihoods in the global land rush? Archetypes of livelihood vulnerability and sustainability potentials. Glob. Environ. Chang. , 41, 153–171, doi:10.1016/j.gloenvcha.2016.10.001.
Obermeier, W.A., et al., 2016: Reduced CO2 fertilization effect in temperate C3 grasslands under more extreme weather conditions. Nat. Clim. Chang. , 7 (2), 137–141, doi:10.1038/nclimate3191.
Obersteiner, M., et al., 2016: Assessing the land resource-food price nexus of the Sustainable Development Goals. Sci. Adv. , 2 (9), e1501499, doi:10.1126/sciadv.1501499.
OECD, 2020: Climate Change: OECD DAC External Development Finance Statistics. https://www.oecd.org/dac/financing-sustainable-development/development-finance-topics/climate-change.htm. (last accessed January 6, 2022).
Oerke, E.C., 2006: Crop losses to pests. J. Agric. Sci. , 144, 31–43, doi:10.1017/S0021859605005708.
Ogalleh, S.A., C.R. Vogl, J. Eitzinger and M. Hauser, 2012: Local perceptions and responses to climate change and variability: the case of Laikipia district, Kenya. Sustainability, 4 (12), 3302–3325, doi:10.3390/su4123302.
Ogier, E., et al., 2020: Responding to climate change: participatory evaluation of adaptation options for key marine fisheries in Australia’s south east. Front. Mar. Sci. , 7, doi:10.3389/fmars.2020.00097.
Ogren, W.L. and R. Chollet, 1982: Photorespiration. In: Photosynthesis[Govindjee(ed.)]. Academic Press, Cambridge, MA, pp. 191–232.
OIV (ed.), OIV Statistical Report on World Vitiviniculture. 2018 World Vitiviniculture Situation, 1–26. ISBN 00085472.
Ojea, E., S.E. Lester and D. Salgueiro-Otero, 2020: Adaptation of fishing communities to climate-driven shifts in target species. One Earth, 2 (6), 544–556, doi:10.1016/j.oneear.2020.05.012.
Oliver, E.C.J., et al., 2019: Projected Marine Heatwaves in the 21st Century and the Potential for Ecological Impact. Front. Mar. Sci. , 6, doi:10.3389/fmars.2019.00734.
Oliver, E.C.J., et al., 2018: Longer and more frequent marine heatwaves over the past century. Nat Commun, 9 (1), 1324, doi:10.1038/s41467-018-03732-9.
Olivier, T., et al., 2020: Urbanization and agricultural intensification destabilize animal communities differently than diversity loss. Nat Commun, 11 (1), 2686, doi:10.1038/s41467-020-16240-6.
Ollat, N. and C.V. Leeuwen, 2016: Climate change impacts and adaptations : new challenges for the wine industry*. Journal of Wine Economics, 11 (1), 139–145, doi:10.1017/jwe.2016.3.
Olorunfemi, F., Mayowa Fasona, Grace Oloukoi, Peter Elias, and vide Adedayo, 2017: traditional knowledge in the use and management of forest ecosystem for livelihoods and food security in Nigerian Savanna. J. Hum. Ecol. , 53, 167–175.
Olsen, E., et al., 2019: Ecological Effects and Ecosystem Shifts Caused by Mass Mortality Events on Early Life Stages of Fish. Front. Mar. Sci. , 6, doi:10.3389/fmars.2019.00669.
Olsson, L., H. Barbosa, S. Bhadwal, A. Cowie, K. Delusca, D. Flores-Renteria, K. Hermans, E. Jobbagy, W. Kurz, D. Li, D.J. Sonwa, L. Stringer, 2019: Land Degradation. In: Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems[Shukla, P. R., J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H. O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi and J. Malley (eds.)]. In press.
Olwig, M.F., C. Noe, R. Kangalawe and E. Luoga, 2016: Inverting the moral economy : the case of land acquisitions for forest plantations in Tanzania. Third World Q, 6597 (January), doi:10.1080/01436597.2015.1078231.
Omari-Motsumi, K., M. Barnett and L. Schalatek, 2019: Broken Connections and Systemic Barriers; Overcoming the Challenge of the “Missing Middle” in Adaptation Finance. Global Commission on Adaptation Background Paper Global Center on Adaptation.
Omori, K., et al., 2020: Assessment of paddy fields’ damage caused by Cyclone Nargis using MODIS time-series images (2004–2013). Paddy Water Environ, doi:10.1007/s10333-020-00829-0.
Oppenheimer, M. and J.K. Anttila-Hughes, 2016: The Science of Climate Change. Future Child. , 26 (1), 11–30.
Oppenheimer, M., B.C. Glavovic , J. Hinkel, R. van de Wal, A.K. Magnan, A. Abd-Elgawad, R. Cai, M. Cifuentes-Jara, R.M. DeConto, T. Ghosh, J. Hay, F. Isla, B. Marzeion, B. Meyssignac, and Z. Sebesvari, 2019: Sea Level Rise and Implications for Low-Lying Islands, Coasts and Communities. In: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate[Pörtner, H.-O., D. C. Roberts, V. VMasson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegría, M. Nicolai, A. Okem, J. Petzold, B. Rama and N. M. Weyer (eds.)], pp. 321–445.
Orchard, B., 2019: Rethinking Climate Finance - Taking Action against Climate Change. Opportunities and Challenges for Financial Players. BlueOrchard, Zurich, Switzerland, https://www.blueorchard.com/rethinking-climate-finance-taking-action-climate-change-opportunities-challenges-financial-players/. (last accessed 6 January, 2022).
Oremus, K.L., et al., 2020: Governance challenges for tropical nations losing fish species due to climate change. Nat. Sustain. , 3 (4), 277–280, doi:10.1038/s41893-020-0476-y.
Ortiz-Bobea, A., et al., 2021: Anthropogenic climate change has slowed global agricultural productivity growth. Nat. Clim. Change, 11 (4), 306–312, doi:10.1038/s41558-021-01000-1.
Ortiz-Bobea, A., E. Knippenberg and R.G. Chambers, 2018: Growing climatic sensitivity of U.S. agriculture linked to technological change and regional specialization. Sci. Adv. , 4 (12), eaat4343, doi:10.1126/sciadv.aat4343.
Ortiz-Colón, G., et al., 2018: Assessing climate vulnerabilities and adaptive strategies for resilient beef and dairy operations in the tropics. Clim Change, 146 (1-2), 47–58, doi:10.1007/s10584-017-2110-1.
Osabuohien, E.S., U.R. Efobi, R.T. Herrmann and C.M.W. Gitau, 2019: Female labor outcomes and large-scale agricultural land investments: Macro-micro evidencefrom Tanzania. Land Use Policy, 82, 716–728, doi:10.1016/j.landusepol.2019.01.005.
Osborne, C.P., et al., 2014: A global database of C4 photosynthesis in grasses. New Phytol. , 204 (3), 441–446, doi:10.1111/nph.12942.
Osgood, D., et al., 2018: Farmer Perception, Recollection, and Remote Sensing in Weather Index Insurance: An Ethiopia Case Study. Remote Sens, 10 (12), doi:10.3390/rs10121887.
Osterhoudt, S., 2018: Remembered resilience: oral history narratives and community resilience in agroforestry systems. Renew. Agric. Food Syst. , 33 (3), 252–255, doi:10.1017/S1742170517000679.
Østhagen, A., J. Spijkers and O.A. Totland, 2020: Collapse of cooperation? The North-Atlantic mackerel dispute and lessons for international cooperation on transboundary fish stocks. Marit. Stud. , 19 (2), 155–165, doi:10.1007/s40152-020-00172-4.
Otekunrin, O.A., O.A. Otekunrin, S. Momoh and I. A. Ayinde, 2019: How far has Africa gone in achieving the zero hunger target? Evidence from Nigeria. Glob. Food Secur. , 22, 1–12, doi:10.1016/j.gfs.2019.08.001.
Otieno, G., et al., 2018: Social Seed Networks for Climate Change Adaptation in Uganda: Strategies to Improve Access to Genetic Diversity and Information. CCAFS Info note, CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS), Wageningen, the Netherlands.
Otto, I.M., et al., 2017: Social vulnerability to climate change: a review of concepts and evidence. Reg Environ Change, 17 (6), 1651–1662, doi:10.1007/s10113-017-1105-9.
Ouedraogo, I., et al., 2018: Closing the Gap between Climate Information Producers and Users: Assessment of Needs and Uptake in Senegal. Climate, 6 (1), doi:10.3390/cli6010013.
Ouédraogo, M., Y. Dembélé and L. Somé, 2010: Perceptions et stratégies d’adaptation aux changements des précipitations: cas des paysans du Burkina Faso. Sci. Et Chang. Planétaires/sécheresse, 21, 87–96.
Ouédraogo, M., et al., 2017: Markets and climate are driving rapid change in farming practices in Savannah West Africa. Reg Environ Change, 17 (2), 437–449, doi:10.1007/s10113-016-1029-9.
Ouma, S., 2014: Situating global finance in the Land Rush Debate: A critical review. Geoforum, 57, 162–166, doi:10.1016/j.geoforum.2014.09.006.
Ouyang, F., et al., 2014: Weakening density dependence from climate change and agricultural intensification triggers pest outbreaks: a 37-year observation of cotton bollworms. Ecol Evol, 4 (17), 3362–3374, doi:10.1002/ece3.1190.
Ovalle-Rivera, O., et al., 2015: Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS ONE, 10 (4), e124155, doi:10.1371/journal.pone.0124155.
Oviedo, A. F., S. Mitraud, D.G. McGrath and M. Bursztyn, 2016: Implementing climate variability adaptation at the community level in the Amazon floodplain. Environ Sci Policy, 63, 151–160.
Owusu-Daaku, K.N., 2018: (Mal)Adaptation opportunism: when other interests take over stated or intended climate change adaptation objectives (and their unintended effects). Local Environ, 23 (9), 934–951, doi:10.1080/13549839.2018.1505836.
Oxenford, H.A. and I. Monnereau, 2018: Climate change impacts, vulnerabilities and adaptations: Western Central Atlantic marine fisheries. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options. Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 185–206. ISBN 978-9251306079.
Oyinlola, M. A., G. Reygondeau, C.C.C. Wabnitz and W.W.L. Cheung, 2020: Projecting global mariculture diversity under climate change. Glob Chang Biol, doi:10.1111/gcb.14974.
Padel, S., L. Levidow and B. Pearce, 2020: UK farmers’ transition pathways towards agroecological farm redesign: evaluating explanatory models. Agroecol. Sustain. Food Syst. , 44 (2), 139–163, doi:10.1080/21683565.2019.1631936.
Paik, S., D.T.P. Le, L.T. Nhu and B.F. Mills, 2020: Salt-tolerant rice variety adoption in the Mekong River Delta: Farmer adaptation to sea-level rise. PLoS ONE, 15 (3), e229464, doi:10.1371/journal.pone.0229464.
Palacios-Abrantes, J., G. Reygondeau, C.C.C. Wabnitz and W.W.L. Cheung, 2020: The transboundary nature of the world’s exploited marine species. Sci Rep, 10 (1), 17668, doi:10.1038/s41598-020-74644-2.
Palahi, M., et al., 2020: Investing in Nature to Transform the Post COVID-19 Economy : A 10-point Action Plan to create a circular bioeconomy devoted to sustainable wellbeing. Solutions J. , 11 (2).
Palhares, J.C.P. and J.R.M. Pezzopane, 2015: Water footprint accounting and scarcity indicators of conventional and organic dairy production systems. J Clean Prod, 93, 299–307, doi:10.1016/j.jclepro.2015.01.035.
Palmer, A., et al., 2017: Between Global and Local: Exploring Regional Food Systems from the Perspectives of Four Communities in the U.S. Northeast. J. Agric. Food Syst. Community Dev. , 187–205, doi:10.5304/jafscd.2017.074.017.
Pandey, R., et al., 2017: Agroecology as a Climate Change Adaptation Strategy for Smallholders of Tehri-Garhwal in the Indian Himalayan Region. Small-scale Forestry, 16 (1), 53–63, doi:10.1007/s11842-016-9342-1.
Paneque-Galvez, J., et al., 2018: High overlap between traditional ecological knowledge and forest conservation found in the Bolivian Amazon. Ambio, 47 (8), 908–923, doi:10.1007/s13280-018-1040-0.
Pang, J., M.H. Ryan, H. Lambers and K.H. Siddique, 2018: Phosphorus acquisition and utilisation in crop legumes under global change. Curr. Opin. Plant Biol. , 45 (Pt B), 248–254, doi:10.1016/j.pbi.2018.05.012.
Panikkar, B. and B. Lemmond, 2020: Being on Land and Sea in Troubled Times: Climate Change and Food Sovereignty in Nunavut. Land, 9 (12), 508, doi:10.3390/land9120508.
Pant, J., et al., 2014: Can aquaculture benefit the extreme poor? A case study of landless and socially marginalized Adivasi (ethnic) communities in Bangladesh. Aquaculture, 418-419, 1–10, doi:10.1016/j.aquaculture.2013.09.027.
Pantanella, E., 2018: Aquaponics production, practices and opportunities. In: Sustainable Aquaculture[Hai, F.I., C. Visvanathan and R. Boopathy (eds.)], pp. 191–248. ISBN 97833197325729783319732565.
Panthou, G., et al., 2018: Rainfall intensification in tropical semi-arid regions: the Sahelian case. Environ. Res. Lett. , 13 (6), doi:10.1088/1748-9326/aac334.
Paolotti, L., et al., 2016: Combining livestock and tree crops to improve sustainability in agriculture: a case study using the Life Cycle Assessment (LCA) approach. J Clean Prod, 131, 351–363, doi:10.1016/j.jclepro.2016.05.024.
Paprocki, K., 2018: Threatening dystopias: development and adaptation regimes in Bangladesh. Ann. Am. Assoc. Geogr. , 108 (4), 955–973, doi:10.1080/24694452.2017.1406330.
Paprocki, K., 2019: All that is solid melts into the bay: anticipatory ruination and climate change adaptation. Antipode, 51 (1), 295–315, doi:10.1111/anti.12421.
Paprocki, K. and S. Huq, 2018: Shrimp and coastal adaptation: on the politics of climate justice. Clim. Dev. , 10 (1), 1–3, doi:10.1080/17565529.2017.1301871.
Parfitt, J., M. Barthel and S. Macnaughton, 2010: Food waste within food supply chains: quantification and potential for change to 2050. Philos. Trans. Royal Soc. B: Biol. Sci. , 365 (1554), 3065–3081, doi:10.1098/rstb.2010.0126.
Park, C.S., et al., 2019: The global effect of extreme weather events on nutrient supply: a superposed epoch analysis. Lancet Planet Health, 3 (10), e429–e438, doi:10.1016/S2542-5196(19)30193-7.
Parker, L., C. Bourgoin, A. Martinez-Valle and P. Laderach, 2019: Vulnerability of the agricultural sector to climate change: The development of a pan-tropical Climate Risk Vulnerability Assessment to inform sub-national decision making. PLoS ONE, 14 (3), e213641, doi:10.1371/journal.pone.0213641.
Parker, L.M., et al., 2015: Persistence of positive carryover effects in the oyster, saccostrea glomerata, following transgenerational exposure to ocean acidification. PLoS ONE, 10 (7), e132276, doi:10.1371/journal.pone.0132276.
Parker, R., 2018: Implications of high animal by-product feed inputs in life cycle assessments of farmed Atlantic salmon. Int J Life Cycle Assess, 23 (5), 982–994, doi:10.1007/s11367-017-1340-9.
Parlee, B. and F. Berkes, 2005: Health of the land, health of the people: a case study on Gwich’in berry harvesting in northern Canada. EcoHealth, 2 (2), 127–137.
Parodi, A., et al., 2018: The potential of future foods for sustainable and healthy diets. Nat. Sustain. , 1 (12), 782–789, doi:10.1038/s41893-018-0189-7.
Parraguez-Vergara, E., J.R. Barton and G. Raposo-Quintana, 2016: Impacts of climate change in the andean foothills of Chile: economic and cultural vulnerability of indigenous Mapuche livelihoods. J. Dev. Soc. , 32 (4), 454–483, doi:10.1177/0169796x16667874.
Partey, S.T., et al., 2020: Gender and climate risk management: evidence of climate information use in Ghana. Clim. Change, 158 (1), 61–75, doi:10.1007/s10584-018-2239-6.
Parvin, S., et al., 2018: Water availability moderates N2 fixation benefit from elevated [CO2 ]: A 2-year free-air CO2 enrichment study on lentil (Lens culinaris MEDIK.) in a water limited agroecosystem. Plant Cell Environ, 41 (10), 2418–2434, doi:10.1111/pce.13360.
Parvin, S., et al., 2019: Grain mineral quality of dryland legumes as affected by elevated CO2 and drought: a FACE study on lentil (Lens culinaris) and faba bean (Vicia faba). Crop. Pasture Sci. , 70 (3), 244–253, doi:10.1071/CP18421.
Pastor, A.V., et al., 2019: The global nexus of food-trade-water sustaining environmental flows by 2050. Nat. Sustain. , 2 (6), 499–507, doi:10.1038/s41893-019-0287-1.
Paterson, R.R.M., et al., 2018: Predominant mycotoxin, mycotoxigenic fungi and climate change related to wine. Food. Res. Int. , 103, 478–491.
Paudel, D., K.R. Tiwari, R.M. Bajracharya and N. Raut, 2017: Agroforestry system: an opportunity for carbon sequestration and climate change adaptation in the mid-hills of Nepal. Octa J. Environ. Res. , 5 (1).
Paudyal, B.R., et al., 2019: Gender Integration in Climate Change and Agricultural Policies: The Case of Nepal. Front. Sustain. Food Syst. , 3, doi:10.3389/fsufs.2019.00066.
Paukert, C.P., et al., 2016: Adapting Inland Fisheries Management to a Changing Climate. Fisheries, 41 (7), 374–384, doi:10.1080/03632415.2016.1185009.
Paul, K.I., et al., 2016: Managing reforestation to sequester carbon, increase biodiversity potential and minimize loss of agricultural land. Land Use Policy, 51, 135–149, doi:10.1016/j.landusepol.2015.10.027.
Paustian, K., et al., 2016: Climate-smart soils. Nature, 532 (7597), 49–57, doi:10.1038/nature17174.
Pearce, W., M. Mahony and S. Raman, 2018: Science advice for global challenges: Learning from trade-offs in the IPCC. Environ Sci Policy, 80, 125–131, doi:10.1016/j.envsci.2017.11.017.
Pearse, R., 2017: Gender and climate change. Wires Clim. Chang. , 8 (2), doi:10.1002/wcc.451.
Peck, M. and J.K. Pinnegar, 2018: Climate change impacts, vulnerabilities and adaptations: North Atlantic and Atlantic Arctic marine fisheries. In: Impacts of climate change on fisheries and aquaculture: synthesis of current knowledge, adaptation and mitigation options[Barange, M., et al.(ed.)]. FAO, Rome, pp. 87–112.
Peck, M. A., et al., 2020: Climate Change and European Fisheries and Aquaculture: CERES Project Synthesis Report . Available at, University of Hamburg, doi:10.25592/uhhfdm.804.
Pecl, G.T., et al., 2017: Biodiversity redistribution under climate change: impacts on ecosystems and human well-being. Science, 355 (6332), eaai9214, doi:10.1126/science.aai9214.
Pecl, G.T., et al., 2019: Autonomous adaptation to climate-driven change in marine biodiversity in a global marine hotspot. Ambio, 48 (12), 1498–1515, doi:10.1007/s13280-019-01186-x.
Pelling, M., et al., 2021: Synergies Between COVID-19 and Climate Change Impacts and Responses. J. Extrem. Events., 08 (03), doi:10.1142/s2345737621310023.
Pendrill, F., et al., 2019: Agricultural and forestry trade drives large share of tropical deforestation emissions. Glob. Environ. Change. , 56, 1–10, doi:10.1016/j.gloenvcha.2019.03.002.
Peng, D.L., et al., 2019: Species distribution modelling and seed germination of four threatened snow lotus (Saussurea), and their implication for conservation. Glob. Ecol. Conserv. , 17, e565, doi:10.1016/j.gecco.2019.e00565.
Peng, W., et al., 2020: A review of historical and recent locust outbreaks: Links to global warming, food security and mitigation strategies. Environ. Res. , 191, 110046–110046, doi:10.1016/j.envres.2020.110046.
Penney, R., G. Wilson and L. Rodwell, 2017: Managing sino-ghanaian fishery relations: A political ecology approach. Mar. Policy. , 79, 46–53, doi:10.1016/j.marpol.2017.02.008.
Pentz, B. and N. Klenk, 2020: Understanding the limitations of current RFMO climate change adaptation strategies: the case of the IATTC and the Eastern Pacific Ocean. Int. Environ. Agreem.: Polit. Law Econ. , 20 (1), 21–39, doi:10.1007/s10784-019-09452-9.
Perera, E., et al., 2021: Effects of genetics and early-life mild hypoxia on size variation in farmed gilthead sea bream (Sparus aurata). Fish Physiol Biochem, 47 (1), 121–133, doi:10.1007/s10695-020-00899-1.
Perez-Garcia, J., L.A. Joyce, A.D. McGuire and X.M. Xiao, 2002: Impacts of climate change on the global forest sector. Clim Change, 54 (4), 439–461, doi:10.1023/A:1016124517309.
Pérez-Marin, A.M., et al., 2017: Agroecological and Social Transformations for Coexistence with Semi-Aridity in Brazil. Sustainability, 9 (6), doi:10.3390/su9060990.
Pérez-Urrestarazu, L., J. Lobillo-Eguíbar, R. Fernández-Cañero and V.M. Fernández-Cabanás, 2019: Suitability and optimization of FAO’s small-scale aquaponics systems for joint production of lettuce (Lactuca sativa) and fish (Carassius auratus). Aquac Eng, 85, 129–137, doi:10.1016/j.aquaeng.2019.04.001.
Perez-Garcia, J., L.A. Joyce, C.S. Binkley and A.D. McGuire, 1997: Economic impacts of climatic change on the global forest sector: An integrated ecological/economic assessment. Crit Rev Environ Sci Technol, 27, 123–138.
Perfecto, I., et al., 2019: Response of Coffee Farms to Hurricane Maria: Resistance and Resilience from an Extreme Climatic Event. Sci Rep, 9 (1), 15668–15668, doi:10.1038/s41598-019-51416-1.
Peri, M., 2017: Climate variability and the volatility of global maize and soybean prices. Food Secur, 9 (4), 673–683, doi:10.1007/s12571-017-0702-2.
Perry, S.J., S. McGregor, S. Gupta and M.H. England, 2017: Future Changes to El Nino-Southern Oscillation Temperature and Precipitation Teleconnections. Geophys. Res. Lett. , 44 (20), 10608–10616, doi:10.1002/2017gl074509.
Peters, K., L. Breitsameter and B. Gerowitt, 2014: Impact of climate change on weeds in agriculture: a review. Agron. Sustain. Dev. , 34 (4), 707–721, doi:10.1007/s13593-014-0245-2.
Peterson, C.A., L.W. Bell, P.C. de F. Carvalho and P.C.F.A.C.M. Gaudin, 2020: Resilience of an Integrated Crop–Livestock System to Climate Change: A Simulation Analysis of Cover Crop Grazing in Southern Brazil. Front. Sustain. Food Syst. , 4, 222–222.
Petheram, L., A.E. Fleming, N. Stacey and A. Perry, 2013: Indigenous women’s preferences for climate change adaptation and aquaculture development to build capacity in the Northern Territory. National Climate Change Adaptation Research Facility Gold Coast, ISBN 978-1925039559.
Pezzopane, J.R.M., et al., 2019: Animal thermal comfort indexes in silvopastoral systems with different tree arrangements. J. Therm. Biol. , 79, 103–111, doi:10.1016/j.jtherbio.2018.12.015.
Phalkey, R.K., et al., 2015: Systematic review of current efforts to quantify the impacts of climate change on undernutrition. Proc. Natl. Acad. Sci. U. S. A. , 112 (33), E4522–4529, doi:10.1073/pnas.1409769112.
Pham, T.T.T., et al., 2021: Guidelines for co-creating climate adaptation plans for fisheries and aquaculture. Clim Change, 164 (3), 62, doi:10.1007/s10584-021-03041-z.
Pham, Y., K. Reardon-Smith, S. Mushtaq and G. Cockfield, 2019: The impact of climate change and variability on coffee production: a systematic review. Clim Change, 156 (4), 609–630, doi:10.1007/s10584-019-02538-y.
Phanxay, I., H. Isao and Y. Satoshi, 2015: Access to Livelihood Assets under the Influence of a Climate Event in Upland Northern Laos. Geogr. Rev. Japan Ser. B, 87 (2), 131–145, doi:10.4157/geogrevjapanb.87.131.
Phogat, V., J.W. Cox and J. Simunek, 2018: Identifying the future water and salinity risks to irrigated viticulture in the Murray-Darling Basin, South Australia. Agric. Water Manag. , 201, 107–117, doi:10.1016/j.agwat.2018.01.025.
Pickering, K., R. Plummer, T. Shaw and G. Pickering, 2015: Assessing the adaptive capacity of the Ontario wine industry for climate change adaptation, Int. J. Wine Res., 7, 13-27, doi:10.2147/ijwr.S73861.
Pietilainen, E.P. and G. Otero, 2019: Power and dispossession in the neoliberal food regime: oil palm expansion in Guatemala. J Peasant Stud, 46 (6), 1142–1166, doi:10.1080/03066150.2018.1499093.
Piggott-McKellar, A.E., K.E. McNamara and P.D. Nunn, 2020: Who defines “good” climate change adaptation and why it matters: a case study from Abaiang Island, Kiribati. Reg Environ Change, 20 (2), doi:10.1007/s10113-020-01614-9.
Pingle, U., 1975: Some Studies in Two Tribal Groups of Central India: Part 2: Nutritive importance of foods consumed in two different seasons. Plant Foods Man, 1 (3-4), 195–208.
Pinsky, M.L., et al., 2020: Fish and fisheries in hot water: What is happening and how do we adapt?Popul. Ecol. , 63 (1), 17–26, doi:10.1002/1438-390x.12050.
Pinsky, M.L., et al., 2018: Preparing ocean governance for species on the move. Science, 360 (6394), 1189–1191, doi:10.1126/science.aat2360.
Pitcher, G.C., et al., 2019: Devastating farmed abalone mortalities attributed to yessotoxin-producing dinoflagellates. Harmful Algae, 81, 30–41, doi:10.1016/j.hal.2018.11.006.
Pittelkow, C.M., et al., 2014: Productivity limits and potentials of the principles of conservation agriculture. Nature, doi:10.1038/nature13809.
Planque, B., et al., 2019: A participatory scenario method to explore the future of marine social-ecological systems. Fish. Fish. , 20 (3), 434–451, doi:10.1111/faf.12356.
Platis, D.P., et al., 2019: Energy analysis, and carbon and water footprint for environmentally friendly farming practices in agroecosystems and agroforestry. Sustainability, 11 (6), 1664–1664, doi:10.3390/su11061664.
Plieninger, T. and M. Schaar, 2008: Modification of land cover in a traditional agroforestry system in spain: processes of tree expansion and regression. Ecol. Soc. , 13 (2), art25, doi:10.5751/ES-02521-130225.
Plieninger, T., D. van der Horst, C. Schleyer and C. Bieling, 2014: Sustaining ecosystem services in cultural landscapes. Ecol. Soc. , 19 (2), art59, doi:10.5751/Es-06159-190259.
Poesch, M.S., et al., 2016: Climate Change Impacts on Freshwater Fishes: A Canadian Perspective. Fisheries, 41 (7), 385–391, doi:10.1080/03632415.2016.1180285.
Polce, C., et al., 2014: Climate-driven spatial mismatches between British orchards and their pollinators: increased risks of pollination deficits. Glob. Change Biol. , 20 (9), 2815–2828, doi:10.1111/gcb.12577.
Pomeroy, R., J. Parks, K.L. Mrakovcich and C. LaMonica, 2016: Drivers and impacts of fisheries scarcity, competition, and conflict on maritime security. Mar. Policy. , 67, 94–104, doi:10.1016/j.marpol.2016.01.005.
Pongthanapanic, T., K.A.T. Nguyen and C.M. Jolli, 2019: Risk management practices of small intensive shrimp farmers in the Mekong Delta of Viet Nam. FAO Fisheries and Aquaculture Circular, Vol. 1194. FAO, Rome, 1–20.
Ponisio, L.C., et al., 2015: Diversification practices reduce organic to conventional yield gap. Proc. R. Soc. B. , 282 (1799), 20141396–20141396, doi:10.1098/rspb.2014.1396.
Ponti, L., A.P. Gutierrez, P.M. Ruti and A. Dell’Aquila, 2014: Fine-scale ecological and economic assessment of climate change on olive in the Mediterranean Basin reveals winners and losers. Proc. Natl. Acad. Sci. USA, 111 (15), 5598–5603, doi:10.1073/pnas.1314437111.
Poore, J. and T. Nemecek, 2019: Reducing food’s environmental impacts through producers and consumers (vol 363, eaaw9908, 2019). Science, 363 (6430), 939–939.
Popke, J. and K. Rhiney, 2019: Introduction to Special Issue on The Caribbean after Irma and Maria: Climate, Development & the Post-Hurricane Context. J. Extrem. Events, 06 (01), doi:10.1142/s2345737619020019.
Porsani, J., M. A. Caretta and K. Lehtilä, 2019: Large-scale land acquisitions aggravate the feminization of poverty: findings from a case study in Mozambique. GeoJournal, 84 (1), 215–236, doi:10.1007/s10708-017-9836-1.
Porter, J.R., et al., 2019: Invited review: Intergovernmental Panel on Climate Change, agriculture, and food-A case of shifting cultivation and history. Glob Chang Biol, 25 (8), 2518–2529, doi:10.1111/gcb.14700.
Porter, J.R., L. Xie, A.J. Challinor, K. Cochrane, S.M. Howden, M.M. Iqbal, D.B. Lobell, and M.I. Travasso, 2014: Food Security and Food Production Systems. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. [Field, C. B., V. R. Barros, D. J. Dokken, K. J. Mach, M. D. Mastrandrea, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea and L. L. White (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 485–533. ISBN 978-1107058071.
Postigo, J.C. and K.R. Young, 2021: Preparing for a diminished cryosphere. Sustain Sci, 2021, 1–4, doi:10.1007/S11625-021-01023-9.
Potopová, V., et al., 2017: The impacts of key adverse weather events on the field-grown vegetable yield variability in the Czech Republic from 1961 to 2014. Int. J. Climatol. , 37 (3), 1648–1664, doi:10.1002/joc.4807.
Potts, S.G., et al., 2010: Global pollinator declines: trends, impacts and drivers. Trends Ecol. Evol. , 25 (6), 345–353, doi:10.1016/j.tree.2010.01.007.
Potts, S.G., et al., 2016: Safeguarding pollinators and their values to human well-being. Nature, 540 (7632), 220–229, doi:10.1038/nature20588.
Poulain, F., A. Himes-Cornell and C. Shelton, 2018: Chapter 35: Methods and tools for climate change adaptation in fisheries and aquaculture. In: Impacts of climate change on fisheries and aquaculture: synthesis of current knowledge, adaptation and mitigation options[Barange, M., et al.(ed.)]. FAO, Rome, pp. 535–566.
Powell, B., et al., 2014: Wild leafy vegetable use and knowledge across multiple sites in Morocco: a case study for transmission of local knowledge?J Ethnobiology Ethnomedicine, 10 (1), 34.
Powell, B., et al., 2015: Improving diets with wild and cultivated biodiversity from across the landscape. Food Secur, 7 (3), 535–554, doi:10.1007/s12571-015-0466-5.
Pradhan, P., et al., 2014: Food self-sufficiency across scales: how local can we go?Environ. Sci. Technol. , 48, 9463–9470.
Pramanik, M., et al., 2018: Predicting climate change impacts on the distribution of the threatened Garcinia indica in the Western Ghats, India. Clim. Risk Manag. , 19, 94–105, doi:10.1016/j.crm.2017.11.002.
Prasad, P.V.V., R. Bheemanahalli and S.V.K. Jagadish, 2017: Field crops and the fear of heat stress-Opportunities, challenges and future directions. Field Crop. Res. , 200, 114–121, doi:10.1016/j.fcr.2016.09.024.
Prestele, R., et al., 2016: Hotspots of uncertainty in land-use and land-cover change projections: a global-scale model comparison. Glob Chang Biol, 22 (12), 3967–3983, doi:10.1111/gcb.13337.
Pretty, J., 2018: Intensification for redesigned and sustainable agricultural systems. Science, 362 (6417), eaav294, doi:10.1126/science.aav0294.
Pretty, J., et al., 2018: for sustainable intensification. Nat. Sustain. , 1 (August), 441–446, doi:10.1038/s41893-018-0114-0.
Pretzsch, H., et al., 2018: Wood density reduced while wood volume growth accelerated in Central European forests since 1870. For. Ecol. Manag. , 429, 589–616, doi:10.1016/j.foreco.2018.07.045.
Prevéy, J.S., et al., 2020: Climate change shifts in habitat suitability and phenology of huckleberry (Vaccinium membranaceum). Agric For Meteorol, 280, 107803.
Prieto, I., et al., 2015: Complementary effects of species and genetic diversity on productivity and stability of sown grasslands. Nat. Plants, 1 (4), 15033, doi:10.1038/nplants.2015.33.
Probstl-Haider, U., et al., 2016: Farmers’ Preferences for Future Agricultural Land Use Under the Consideration of Climate Change. Environ Manage, 58 (3), 446–464, doi:10.1007/s00267-016-0720-4.
Prudent, M., et al., 2020: The diversity of soil microbial communities matters when legumes face drought. Plant Cell Environ, 43 (4), 1023–1035, doi:10.1111/pce.13712.
Puchta, H., 2017: Applying CRISPR/Cas for genome engineering in plants: the best is yet to come. Curr. Opin. Plant Biol. , 36, 1–8, doi:10.1016/j.pbi.2016.11.011.
Pulatov, B., M.L. Linderson, K. Hall and A.M. Jonsson, 2015: Modeling climate change impact on potato crop phenology, and risk of frost damage and heat stress in northern Europe. Agric For Meteorol, 214, 281–292, doi:10.1016/j.agrformet.2015.08.266.
Purcell, C., et al., 2018: Increasing stomatal conductance in response to rising atmospheric CO2. Ann Bot , 121 (6), 1137–1149, doi:10.1093/aob/mcx208.
Purcell, S.W. and R.S. Pomeroy, 2015: Driving small-scale fisheries in developing countries. Front. Mar. Sci. , 2 (44), doi:10.3389/fmars.2015.00044.
Putkonen, J. and G. Roe, 2003: Rain-on-snow events impact soil temperatures and affect ungulate survival. Geophys. Res. Lett. , 30 (4), doi:10.1029/2002gl016326.
Pye, O., I. Radjawali and Julia, 2017: Land grabs and the river: eco-social transformations along the Kapuas, Indonesia. Can. J. Dev. Stud. / Revue Can. D’études Du Développement , 38 (3), 378–394, doi:10.1080/02255189.2017.1298518.
Qaim, M., 2016: Genetically Modified Crops and Agricultural Development . Palgrave Macmillan, New York City, New York.
Qi, R.M., et al., 2016: Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Appl. Soil Ecol. , 102, 36–45, doi:10.1016/j.apsoil.2016.02.004.
Quan, S.P., et al., 2019: Adaptation to climate change and its impacts on wheat yield: perspective of farmers in Henan of China. Sustainability, 11 (7), doi:10.3390/su11071928.
Quandt, A., H. Neufeldt and J.T. McCabe, 2017: The role of agroforestry in building livelihood resilience to floods and drought in semiarid Kenya. Ecol. Soc. , 22 (3), art10, doi:10.5751/Es-09461-220310.
Quandt, A., H. Neufeldt and J.T. McCabe, 2019: Building livelihood resilience: what role does agroforestry play?Clim. Dev. , 11 (6), 485–500, doi:10.1080/17565529.2018.1447903.
Quiller, G., et al., 2017: Heat exposure and productivity in orchards: implications for climate change research. Arch Environ Occup Health, 72 (6), 313–316, doi:10.1080/19338244.2017.1288077.
Quiñones-Ruiz, X.F., M. Penker, C.R. Vogl and L.F. Samper-Gartner, 2015: Can origin labels re-shape relationships along international supply chains? – The case of Café de Colombia. Int. J. Commons, 9 (1), 416–416, doi:10.18352/ijc.529.
Quintas-Soriano, C., A.J. Castro, H. Castro and M. Garcia-Llorente, 2016: Impacts of land use change on ecosystem services and implications for human well-being in Spanish drylands. Land Use Policy, 54, 534–548, doi:10.1016/j.landusepol.2016.03.011.
Quiroga, S., C. Suárez, D. Solís and P. Martinez-Juarez, 2020: Framing vulnerability and coffee farmers’ behaviour in the context of climate change adaptation in Nicaragua. World Dev, 126, 104733–104733, doi:10.1016/j.worlddev.2019.104733.
Raftopoulos, M. and D. Short, 2019: Implementing free prior and informed consent: the United Nations Declaration on the Rights of Indigenous Peoples (2007), the challenges of REDD+ and the case for the precautionary principle. Int. J. Hum. Rights, 23 (1-2), 87–103, doi:10.1080/13642987.2019.1579990.
Rahimi, J., et al., 2020: Will dairy cattle production in West Africa be challenged by heat stress in the future?Clim Change, 161 (4), 665–685, doi:10.1007/s10584-020-02733-2.
Rahman, A., M. A. Mojid and S. Banu, 2018: Climate change impact assessment on three major crops in the north-central region of Bangladesh using DSSAT. Int. J. Agric. Biol. Eng. , 11 (4), 135–143, doi:10.25165/j.ijabe.20181104.3331.
Rahman, H.M.T. and G.M. Hickey, 2019: What does autonomous adaptation to climate change have to teach public policy and planning about avoiding the risks of maladaptation in Bangladesh?Front. Environ. Sci. , 7, doi:10.3389/fenvs.2019.00002.
Rajab, Y.A., et al., 2016: Cacao cultivation under diverse shade tree cover allows high carbon storage and sequestration without yield losses. PLoS ONE, 11 (2), e149949.
Ramachandran, A., et al., 2017: Spatiotemporal analysis of projected impacts of climate change on the major C3 and C4 crop yield under representative concentration pathway 4.5: Insight from the coasts of Tamil Nadu, South India. PLoS ONE, 12 (7), e180706, doi:10.1371/journal.pone.0180706.
Ramachandran Nair, P.K., M. Kumar and V.D. Nair, 2009: Agroforestry as a strategy for carbon sequestration. J Plant Nutr Soil Sci, 172 (1), 10–23, doi:10.1002/jpln.200800030.
Ramírez, F. and J. Kallarackal, 2015: Responses of Fruit Trees to Global Climate Change. Springer, Cham, ISBN 978-3-319-14199-2.
Ramesh, K., et al., 2017: Weeds in a changing climate: vulnerabilities, consequences, and implications for future weed management. Front. Plant Sci. , 8, 95, doi:10.3389/fpls.2017.00095.
Ramirez, P.J.B., T.A. Narvaez and E.J. Santos-Ramirez, 2020: Gender-inclusive value chains: the case of seaweed farming in Zamboanga Peninsula, Philippines. gender Technol. Dev. , 24 (1), 110–130, doi:10.1080/09718524.2020.1728810.
Ramos, A.K., et al., 2020: Invisible no more: the impact of COVID-19 on essential food production workers. J. Agromed. , 25 (4), 378–382, doi:10.1080/1059924X.2020.1814925.
Ranasinghe, R., A.C. Ruane, R. Vautard, N. Arnell, E. Coppola, F.A. Cruz, S. Dessai, A.S. Islam, M. Rahimi, D. Ruiz Carrascal, J. Sillmann, M.B. Sylla, C. Tebaldi, W. Wang, and R. Zaaboul, 2021: Climate Change Information for Regional Impact and for Risk Assessment. In: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. [Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press, UK.
Randriambanona, H., J.R. Randriamalala and S.M. Carrière, 2019: Native forest regeneration and vegetation dynamics in non-native Pinus patula tree plantations in Madagascar. For. Ecol. Manag. , 446, 20–28, doi:10.1016/j.foreco.2019.05.019.
Ranjan, R., 2020: Protecting warming lakes through climate-adaptive PES mechanisms. Ecol. Econ. , 177, doi:10.1016/j.ecolecon.2020.106782.
Ranjitkar, S., et al., 2020: Will heat stress take its toll on milk production in China?Clim. Chang. , 1–16, doi:10.1007/s10584-020-02688-4.
Rao, N., et al., 2019a: Gendered vulnerabilities to climate change: insights from the semi-arid regions of Africa and Asia. Clim. Dev. , 11 (1), 14–26, doi:10.1080/17565529.2017.1372266.
Rao, N., et al., 2019b: A qualitative comparative analysis of women’s agency and adaptive capacity in climate change hotspots in Asia and Africa. Nat. Clim. Chang. , 9 (12), 964–971, doi:10.1038/s41558-019-0638-y.
Rasmussen, L.V., et al., 2018: Social-ecological outcomes of agricultural intensification. Nat. Sustain. , 1 (6), 275–282, doi:10.1038/s41893-018-0070-8.
Rasul, G., et al., 2019: Food and Nutrition Security in the Hindu Kush Himalaya: Unique Challenges and Niche Opportunities. In: The Hindu Kush Himalaya Assessment . Springer International Publishing, Cham, pp. 301–338.
Rathore, P., A. Roy and H. Karnatak, 2019: Modelling the vulnerability of Taxus wallichiana to climate change scenarios in South East Asia. Ecol Indic, 102, 199–207, doi:10.1016/j.ecolind.2019.02.020.
Raven, J.A., C.J. Gobler and P.J. Hansen, 2020: Dynamic CO2 and pH levels in coastal, estuarine, and inland waters: theoretical and observed effects on harmful algal blooms. Harmful Algae, 91, 101594, doi:10.1016/j.hal.2019.03.012.
Ravi, S., D.D. Breshears, T.E. Huxman and P. D’Odorico, 2010: Land degradation in drylands: Interactions among hydrologic-aeolian erosion and vegetation dynamics. Geomorphology, 116 (3), 236–245, doi:10.1016/j.geomorph.2009.11.023.
Ravikumar, A., et al., 2015: Multilevel governance challenges in transitioning towards a national approach for REDD plus : evidence from 23 subnational REDD plus initiatives. Int. J. Commons, 9 (2), 909–931, doi:10.18352/ijc.593.
Raworth, K., 2017: A doughnut for the Anthropocene: humanity’s compass in the 21st century. Lancet Planet Health, 1 (2), e48–e49, doi:10.1016/S2542-5196(17)30028-1.
Ray, D.K., J.S. Gerber, G.K. MacDonald and P.C. West, 2015: Climate variation explains a third of global crop yield variability. Nat. Commun. , 6, 5989, doi:10.1038/ncomms6989.
Ray, D.K., et al., 2019: Climate change has likely already affected global food production. PLoS ONE, 14 (5), e217148, doi:10.1371/journal.pone.0217148.
Raymond-Yakoubian, J. and R. Daniel, 2018: An Indigenous approach to ocean planning and policy in the Bering Strait region of Alaska. Mar. Policy. , 97, 101–108, doi:10.1016/j.marpol.2018.08.028.
Raymond-Yakoubian, J., B. Raymond-Yakoubian and C. Moncrieff, 2017: The incorporation of traditional knowledge into Alaska federal fisheries management. Mar. Policy. , 78, 132–142, doi:10.1016/j.marpol.2016.12.024.
Raymundo, R., S. Asseng, D. Cammarano and R. Quiroz, 2014: Potato, sweet potato, and yam models for climate change: a review. Field Crops Res. , 166, 173–185, doi:10.1016/j.fcr.2014.06.017.
Raymundo, R., et al., 2018: Climate change impact on global potato production. Eur J Agron, 100, 87–98, doi:10.1016/j.eja.2017.11.008.
Reed, J., J. van Vianen, J. Barlow and T. Sunderland, 2017: Have integrated landscape approaches reconciled societal and environmental issues in the tropics?Land Use Policy, 63, 481–492, doi:10.1016/j.landusepol.2017.02.021.
Reed, J., et al., 2016: Integrated landscape approaches to managing social and environmental issues in the tropics: learning from the past to guide the future. Glob Chang Biol, 22 (7), 2540–2554, doi:10.1111/gcb.13284.
Reeves, M.C., et al., 2020: An Assessment of Production Trends on the Great Plains from 1984 to 2017. Rangel. Ecol. Manag. , doi:10.1016/j.rama.2020.01.011.
Refatti, J.P., et al., 2019: High [CO2] and Temperature Increase Resistance to Cyhalofop-Butyl in Multiple-Resistant Echinochloa colona. Front. Plant Sci. , 10, 529, doi:10.3389/fpls.2019.00529.
Reich, P.B., S.E. Hobbie, T.D. Lee and M. A. Pastore, 2018: Unexpected reversal of C3 versus C4 grass response to elevated CO2 during a 20-year field experiment. Science, 360 (6386), 317–320.
Reid, G.K., et al., 2019: Climate change and aquaculture: considering biological response and resources. Aquac. Environ. Interactions, 11, 569–602, doi:10.3354/aei00332.
Reid, R.S., M. E. Fernandez-Gimenez and K. A. Galvin, 2014: Dynamics and Resilience of Rangelands and Pastoral Peoples Around the Globe. Annu. Rev. Environ. Resour. , 39 (39), 217–242, doi:10.1146/annurev-environ-020713-163329.
Reimer, J.R., H. Caswell, A.E. Derocher and M. A. Lewis, 2019: Ringed seal demography in a changing climate. Ecol Appl, 29 (3), e1855, doi:10.1002/eap.1855.
Reisman, E., 2019: The great almond debate: A subtle double movement in California water. Geoforum, 104, 137–146, doi:10.1016/j.geoforum.2019.04.021.
Renard, D. and D. Tilman, 2019: National food production stabilized by crop diversity. Nature, 571 (7764), 257–260, doi:10.1038/s41586-019-1316-y.
Renaud, F.G., et al., 2015: Resilience and shifts in agro-ecosystems facing increasing sea-level rise and salinity intrusion in Ben Tre Province, Mekong Delta. Clim Change, 133 (1), 69–84, doi:10.1007/s10584-014-1113-4.
Renner, S.S. and C.M. Zohner, 2018: Climate Change and Phenological Mismatch in Trophic Interactions Among Plants, Insects, and Vertebrates. Annu. Rev. Ecol. Evol. Syst. , 49 (49), 165–182, doi:10.1146/annurev-ecolsys-110617-062535.
Renwick, L.L.R., et al., 2020: Maize-Pigeonpea Intercropping Outperforms Monocultures Under Drought. Front. Sustain. Food Syst. , 4, 253–253.
Reo, N.J. and A.K. Parker, 2013: Re-thinking colonialism to prepare for the impacts of rapid environmental change. In: Climate Change and Indigenous Peoples in the United States. [Maldonado, J. K., B. Colombi and R. Pandya (eds.)]. Springer, Heidelberg, New York, Dordrecht, London, pp. 163-174.
Reppin, S., et al., 2019: Contribution of agroforestry to climate change mitigation and livelihoods in Western Kenya. Agrofor. Syst. , 94 (1), 203–220, doi:10.1007/s10457-019-00383-7.
Resco, P., A. Iglesias, I. Bardaji and V. Sotes, 2016: Exploring adaptation choices for grapevine regions in Spain. Reg Environ Change, 16 (4), 979–993, doi:10.1007/s10113-015-0811-4.
Resosudarmo, I.A.P., et al., 2014: Does tenure security lead to REDD+ project effectiveness? Reflections from five emerging sites in Indonesia. World Dev, 55, 68–83, doi:10.1016/j.worlddev.2013.01.015.
Rességuier, L.D., S. Mary, R.L. Roux and T. Petitjean, 2020: Temperature variability at local scale in the Bordeaux area. Relations with environmental factors and impact on vne Phenology. Front. Plant Sci. , 11 (May), 1–20, doi:10.3389/fpls.2020.00515.
Reverter, M., et al., 2020: Moving towards more sustainable aquaculture practices: a meta-analysis on the potential of plant-enriched diets to improve fish growth, immunity and disease resistance. Rev. Aquac. , 13 (1), 537–555, doi:10.1111/raq.12485.
Revi, A., D.E. Satterthwaite, F. Aragón-Durand, J. Corfee-Morlot, R.B.R. Kiunsi, M. Pelling, D.C. Roberts, and W. Solecki, 2014: Urban Areas. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[Field, C. B., V. R. Barros, D. J. Dokken, K. J. Mach, M. D. Mastrandrea, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea and L. L. White (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 535–612. ISBN 9781107058071.
Reyer, C.P.O., et al., 2017: Are forest disturbances amplifying or canceling out climate change-induced productivity changes in European forests?Environ. Res. Lett. , 12 (3), 34027, doi:10.1088/1748-9326/aa5ef1.
Reyes-García, V., et al., 2014: Resilience of traditional knowledge systems. Glob. Environ. Chang. , 24, 223–231.
Reyes-García, V., et al., 2019: Dietary transitions among three contemporary hunter-gatherers across the tropics. Food Secur, 11 (1), 109–122.
Reyes-Garcia, V., et al., 2016: Local indicators of climate change: The potential contribution of local knowledge to climate research. Wiley. Interdiscip. Rev. Clim. Change. , 7 (1), 109–124, doi:10.1002/wcc.374.
Reyes-Garcia, V., et al., 2019: A collaborative approach to bring insights from local observations of climate change impacts into global climate change research. Curr. Opin. Environ. Sustain. , 39 (1-8), 1–8, doi:10.1016/j.cosust.2019.04.007.
Rhiney, K., A. Eitzinger, A.D. Farrell and S.D. Prager, 2018: Assessing the implications of a 1.5 degrees C temperature limit for the Jamaican agriculture sector. Reg. Environ. Change, 18 (8), 2313–2327, doi:10.1007/s10113-018-1409-4.
Rhoné, B., et al., 2020: Pearl millet genomic vulnerability to climate change in West Africa highlights the need for regional collaboration. Nat Commun, 11 (1), 5274, doi:10.1038/s41467-020-19066-4.
Ribot, J., 2014: Cause and response: vulnerability and climate in the Anthropocene. J Peasant Stud, 41 (5), 667–705, doi:10.1080/03066150.2014.894911.
Ricciardi, V., et al., 2018: How much of the world’s food do smallholders produce?Glob. Food Secur. , 17 (June), 64–72, doi:10.1016/j.gfs.2018.05.002.
Richards, C. and K. Lyons, 2016: The new corporate enclosures: Plantation forestry, carbon markets and the limits of financialised solutions to the climate crisis. Land Use Policy, 56, 209–216, doi:10.1016/j.landusepol.2016.05.013.
Richards, G., J. Frehs, E. Myers and M. Van Bibber, 2019: Commentary - the climate change and health adaptation program: indigenous climate leaders’ championing adaptation efforts. Health Promot Chronic Dis Prev Can, 39 (4), 127–130, doi:10.24095/hpcdp.39.4.03.
Richardson, K.J., et al., 2018: Food security outcomes under a changing climate: impacts of mitigation and adaptation on vulnerability to food insecurity. Clim Change, 147 (1-2), 327–341, doi:10.1007/s10584-018-2137-y.
Ricker-Gilbert, J. and J. Chamberlin, 2018: Transaction costs, land rental markets, and their impact on youth access to agriculture in Tanzania. Land. Econ. , 94 (4), 541–555, doi:10.3368/le.94.4.541.
Rigolot, C., et al., 2017: Interactions between intervention packages, climatic risk, climate change and food security in mixed crop-livestock systems in Burkina Faso. Agric Syst , 151, 217–224, doi:10.1016/j.agsy.2015.12.017.
Rinaldi, L., et al., 2015: Sheep and Fasciola hepatica in Europe: the GLOWORM experience. Geospat Health, 9 (2), 309–317, doi:10.4081/gh.2015.353.
Rivera-Ferre, M.G., et al., 2016: Re-framing the climate change debate in the livestock sector: mitigation and adaptation options. Wiley. Interdiscip. Rev. Clim. Change. , 7 (6), 869–892, doi:10.1002/wcc.421.
Robalino, J., J. Jimenez and A. Chacon, 2015: The Effect of Hydro-Meteorological Emergencies on Internal Migration. World Dev, 67, 438–448, doi:10.1016/j.worlddev.2014.10.031.
Robinson, B.E., M.B. Holland and L. Naughton-Treves, 2014: Does secure land tenure save forests? A meta-analysis of the relationship between land tenure and tropical deforestation. Glob. Environ. Change. , 29, 281–293, doi:10.1016/j.gloenvcha.2013.05.012.
Robinson, C., et al., 2016a: Participatory mapping to negotiate indigenous knowledge used to assess environmental risk. Sustain Sci, 11 (1), 115–126, doi:10.1007/s11625-015-0292-x.
Robinson, C.J., et al., 2016b: Indigenous benefits and carbon offset schemes: An Australian case study. Environ Sci Policy, 56, 129–134, doi:10.1016/j.envsci.2015.11.007.
Robinson, L.M., et al., 2015: Rapid assessment of an ocean warming hotspot reveals “high” confidence in potential species’ range extensions. Glob. Environ. Change. , 31, 28–37, doi:10.1016/j.gloenvcha.2014.12.003.
Robinson, T.P., et al., 2011: Global livestock production systems. FAO and ILRI, Rome
Rochedo, P.R.R., et al., 2018: The threat of political bargaining to climate mitigation in Brazil. Nat. Clim. Chang. , 8 (8), 695, doi:10.1038/s41558-018-0213-y.
Rockström, J., et al., 2017: Sustainable intensification of agriculture for human prosperity and global sustainability. Ambio, 46 (1), 4–17, doi:10.1007/s13280-016-0793-6.
Rode, J., E. Gomez-Baggethun and T. Krause, 2015: Motivation crowding by economic incentives in conservation policy: A review of the empirical evidence. Ecol. Econ. , 117, 270–282, doi:10.1016/j.ecolecon.2014.11.019.
Rodríguez-de-Francisco, J.C., et al., 2021: Post-conflict transition and REDD+ in Colombia: Challenges to reducing deforestation in the Amazon. Policy Econ, 127 (August 2019)), 102450–102450, doi:10.1016/j.forpol.2021.102450.
Rodríguez, A., et al., 2019: Implications of crop model ensemble size and composition for estimates of adaptation effects and agreement of recommendations. Agric For Meteorol, 264 (2019), 351–362, doi:10.1016/j.agrformet.2018.09.018.
Rodriguez-Llanes, J.M., S. Ranjan-Dash, A. Mukhopadhyay and D. Guha-Sapir, 2016: Flood-Exposure is Associated with Higher Prevalence of Child Undernutrition in Rural Eastern India. Int J Environ Res Public Health, 13 (2), 210, doi:10.3390/ijerph13020210.
Roesch-McNally, G.E., J.G. Arbuckle and J.C. Tyndall, 2018: Barriers to implementing climate resilient agricultural strategies: the case of crop diversification in the U.S. Corn Belt. Glob. Environ. Change, 48, 206–215, doi:10.1016/j.gloenvcha.2017.12.002.
Rogelj, J. et al., 2018: Mitigation Pathways Compatible with 1.5°C in the Context of Sustainable Development. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. [Masson-Delmotte, V., P. Zhai, H. O. Pörtner, D. Roberts, J. Skea, P. R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J. B. R. Matthews, Y. Chen, X. Zhou, M. I. Gomis, E. Lonnoy, T. Maycock, M. Tignor and T. Waterfield (eds.)]. In press.
Rojas-Downing, M.M., A.P. Nejadhashemi, T. Harrigan and S.A. Woznicki, 2017: Climate change and livestock: Impacts, adaptation, and mitigation. Clim. Risk Manag. , 16, 145–163, doi:10.1016/j.crm.2017.02.001.
Rojas, M., F. Lambert, J. Ramirez-Villegas and A.J. Challinor, 2019: Emergence of robust precipitation changes across crop production areas in the 21st century. Proc. Natl. Acad. Sci. U. S. A. , 116 (14), 6673–6678, doi:10.1073/pnas.1811463116.
Rolo, V. and G. Moreno, 2019: Shrub encroachment and climate change increase the exposure to drought of Mediterranean wood-pastures. Sci. Total Environ. , 660, 550–558, doi:10.1016/j.scitotenv.2019.01.029.
Romeo, D., E. B. Vea and M. Thomsen, 2018: Environmental Impacts of Urban Hydroponics in Europe: A Case Study in Lyon. Procedia CIRP, 69, 540-545, doi:10.1016/j.procir.2017.11.048.
Roncoli, C., et al., 2009: From accessing to assessing forecasts: an end-to-end study of participatory climate forecast dissemination in Burkina Faso (West Africa). Clim Change, 92 (3-4), 433–460, doi:10.1007/s10584-008-9445-6.
Roncoli, C., T.A. Crane. and B. Orlove, 2009: Fielding climate change in cultural anthropology. In: Anthropology and Climate Change[Crate, S. A. and M. Nuttall (eds.)]. Left Coast Press, Inc. Walnut Creek, California, United States, pp. 87-115. ISBN 9781598743333. https://ssrn.com/abstract=2396931. (last accessed on January, 31, 2022).
Rondeau, R.J., K.L. Decker and G.A. Doyle, 2018: Potential Consequences of Repeated Severe Drought for Shortgrass Steppe Species. Rangel. Ecol. Manag. , 71 (1), 91–97, doi:10.1016/j.rama.2017.07.002.
Rosa-Schleich, J., J. Loos, O. Musshoff and T. Tscharntke, 2019: Ecological-economic trade-offs of Diversified Farming Systems - A review. Ecol. Econ. , 160, 251–263, doi:10.1016/j.ecolecon.2019.03.002.
Rosa, L., et al., 2021: Energy implications of the 21st century agrarian transition. Nat Commun, 12 (1), 2319–2319, doi:10.1038/s41467-021-22581-7.
Rose, J.B. and F. Wu, 2015: Waterborne and Foodborne Diseases. In: Climate Change and Public Health. Oxford University Press, New York, pp. 157–172.
Rosegrant, M.W., M.M. Dey, R. Valmonte-Santos and O.L. Chen, 2016: Economic impacts of climate change and climate change adaptation strategies in Vanuatu and Timor-Leste. Mar. Policy. , 67, 179–188, doi:10.1016/j.marpol.2015.12.010.
Rosenstock, T.S., et al., 2014: Agroforestry with N2-fixing trees: sustainable development’s friend or foe?Curr. Opin. Environ. Sustain. , 6, 15–21, doi:10.1016/j.cosust.2013.09.001.
Rosenzweig, C., et al., 2014: Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison. Proc. Natl. Acad. Sci. USA, 111 (9), 3268–3273, doi:10.1073/pnas.1222463110.
Rosenzweig, C., et al., 2020: Climate change responses benefit from a global food system approach. Nat. Food, 1 (2), 94–97, doi:10.1038/s43016-020-0031-z.
Rosenzweig, C., et al., 2018: Coordinating AgMIP data and models across global and regional scales for 1.5 degrees C and 2.0 degrees C assessments. Philos. Trans. A. Math. Phys. Eng. Sci. , 376 (2119), 20160455, doi:10.1098/rsta.2016.0455.
Rosimo, M., J. Dalusag, J. Gonsalves and R. Vidallo, 2018: Incorporating a Gender Dimension in a Participatory Community-Based Project: A Case Study in Guinayangan Quezon Province, Philippines. In: Gender dimension of climate change research in agriculture (Case studies in Southeast Asia) [Paris, T. and M.F. Rola-Rubzen(eds.)]. CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS), Wageningen, the Netherlands, pp. 157–179.
Rosol, R., S. Powell-Hellyer and H.M. Chan, 2016: Impacts of decline harvest of country food on nutrient intake among Inuit in Arctic Canada: impact of climate change and possible adaptation plan. Int. J. Circumpolar Health, 75, 31127, doi:10.3402/ijch.v75.31127.
Ross, N., A. Agostini and M. Frick, 2016: Agricultural sectors in nationally determined contributions (NDCs): priority areas for international support . FAO, http://www.fao.org/3/a-i6400e.pdf. (last accessed August 31, 2021)
Ross, P. and C.W. Mason, 2020: “we hardly have any moose around here anymore”: climate change and the barriers to food security in the Dehcho region, northwest territories. Arctic, 73 (3), 368–385, doi:10.14430/arctic71082.
Rossa, A., M. Flubacher, L. Cristobal, H. Ramos and F. Lechthaler, 2020: Towards More Resilient Food Systems for Smallholder Farmers in the Peruvian Altiplano: The Potential of Community-Based Climate Services. In: Handbook of Climate Services. [Filho, W. L. and D. Jacob (eds.)]. Springer, Cham, pp. 327-351.Rossi, R. and A. Miola, 2017: Adaptation measures in Intended Nationally Determined Contributions from Small Island Developing States and Least Developed Countries - An analysis of communicated costs and information exhaustiveness. Publications Office of the European Union, Luxembourg.
Rotz, S., et al., 2019: Automated pastures and the digital divide: How agricultural technologies are shaping labour and rural communities. J Rural Stud, 68, 112–122, doi:10.1016/j.jrurstud.2019.01.023.
Roudier, P., et al., 2016: Assessing the benefits of weather and seasonal forecasts to millet growers in Niger. Agric For Meteorol, 223, 168–180, doi:10.1016/j.agrformet.2016.04.010.
Roudier, P., et al., 2014: The role of climate forecasts in smallholder agriculture: Lessons from participatory research in two communities in Senegal. Clim. Risk Manag. , 2, 42–55, doi:10.1016/j.crm.2014.02.001.
Roue, M. and M. Roue, 2018: Indigenous and local knowledge and science: from validation to knowledge coproduction. In: The International Encyclopedia of Anthropology, [Callan, H. (ed.)]. John Wiley & Sons, Ltd, US, pp. 1-11.
Roupsard, O., et al., 2020: How far does the tree affect the crop in agroforestry? New spatial analysis methods in a Faidherbia parkland. Agric. Ecosyst. Environ. , 296, doi:10.1016/j.agee.2020.106928.
Roux, M.-J., R.F. Tallman and Z.A. Martin, 2019: Small-scale fisheries in Canada’s Arctic: combining science and fishers knowledge towards sustainable management. Mar. Policy, 101, 177–186, doi:10.1016/j.marpol.2018.01.016.
Rowland, D., et al., 2017: Forest foods and healthy diets: quantifying the contributions. Environ Conserv, 44 (2), 102–114, doi:10.1017/S0376892916000151.
Roxy, M., et al., 2017: A threefold rise in widespread extreme rain events over central India. Nat Commun, 3 (1), 1–1.
Roxy, M.K., K. Ritika, P. Terray and S. Masson, 2014: The Curious Case of Indian Ocean Warming. J Clim, 27 (22), 8501–8509, doi:10.1175/Jcli-D-14-00471.1.
Roy, J., P. Tschakert, H. Waisman, S. Abdul Halim, P. Antwi-Agyei, P. Dasgupta, B. Hayward, M. Kanninen, D. Liverman, C. Okereke, P.F. Pinho, K. Riahi, and A.G. Suarez Rodriguez, 2018: Sustainable Development, Poverty Eradication and Reducing Inequalities. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty[Masson-Delmotte, V., P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P.R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J.B.R. Matthews, Y. Chen, X. Zhou, M.I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, and T. Waterfield (eds.)]. In Press.
RRI, 2018a: A global baseline of carbon storage in collective lands: Indigenous and local community contributions to climate change mitigation. RRI, Washington, DC, USA. 12 pp.
RRI, 2018b: Uncertainty and opportunity: The status of forest carbon rights and governance frameworks in over half of the world’s tropical forests. RRI, Washington, DC, USA. 23 pp.
Ruane, A.C., 2014: Carbon-Temperature-Water change analysis for peanut production under climate change: A prototype for the AgMIP Coordinated Climate-Crop Modeling Project (C3 MP). Glob. Chang. Biol. , 20, 394–407.
Ruane, A.C., et al., 2018: Biophysical and economic implications for agriculture of +1.5 degrees and +2.0 degrees C global warming using AgMIP Coordinated Global and Regional Assessments. Clim. Res. , 76 (1), 17–39, doi:10.3354/cr01520.
Ruane, A.C., et al., 2021: Strong regional influence of climatic forcing datasets on global crop model ensembles. Agric For Meteorol, 300, 108313.
Ruane, A.C., et al., 2017: An AgMIP framework for improved agricultural representation in IAMs. Environ. Res. Lett. , 12 (12), 125003, doi:10.1088/1748-9326/aa8da6.
Rubio, I., U. Ganzedo, A.J. Hobday and E. Ojea, 2020a: Southward re-distribution of tropical tuna fisheries activity can be explained by technological and management change. Fish Fish (oxf) , 21 (3), 511–521, doi:10.1111/faf.12443.
Rubio, N.R., N. Xiang and D.L. Kaplan, 2020b: Plant-based and cell-based approaches to meat production. Nat Commun, 11 (1), doi:10.1038/s41467-020-20061-y.
Rufino, M.C., et al., 2013: Transitions in agro-pastoralist systems of East Africa: Impacts on food security and poverty. Agric Ecosyst Environ, 179, 215–230, doi:10.1016/j.agee.2013.08.019.
Ruiz-Vera, U.M., et al., 2013: Global warming can negate the expected CO2 stimulation in photosynthesis and productivity for soybean grown in the Midwestern United States. Plant Physiol. , 162 (1), 410–423, doi:10.1104/pp.112.211938.
Ruiz Meza, L.E., 2015: Adaptive capacity of small-scale coffee farmers to climate change impacts in the Soconusco region of Chiapas, Mexico. Clim. Dev. , 7 (2), 100–109, doi:10.1080/17565529.2014.900472.
Rumbach, A. and D. Foley, 2014: Indigenous Institutions and Their Role in Disaster Risk Reduction and Resilience: Evidence from the 2009 Tsunami in American Samoa. Ecol. Soc. , 19 (1), 1–10, doi:10.5751/Es-06189-190119.
Ruml, M., et al., 2016: Response of grapevine phenology to recent temperature change and variability in the wine-producing area of Sremski Karlovci, Serbia. J. Agric. Sci. , 154 (2), 186–206, doi:10.1017/S0021859615000453.
Ruoso, L.E., et al., 2015: Reading Ecosystem Services at the Local Scale through a Territorial Approach: the Case of Peri-Urban Agriculture in the Thau Lagoon, Southern France. Ecol. Soc. , 20 (3), art11, doi:10.5751/Es-07694-200311.
Rushton, J., et al., 2018: Interactions between intensifying livestock production for food and nutrition security, and increased vulnerability to AMR and zoonoses. CGIAR Independent Science & Partnership Council (ISPC) Secretariat, Rome.
Rybråten, S., M. Bjørkan, G.K. Hovelsrud and B.P. Kaltenborn, 2018: Sustainable coasts? Perceptions of change and livelihood vulnerability in Nordland, Norway. Local Environ, 23 (12), 1156–1171, doi:10.1080/13549839.2018.1533931.
Ryngaert, C., 2017: Climate Change Mitigation Techniques and International Law: Assessing the Externalities of Reforestation and Geoengineering. Ratio Juris, 30 (3), 273–289, doi:10.1111/raju.12154.
Sachs, J.D., et al., 2019: Six Transformations to achieve the Sustainable Development Goals. Nat. Sustain. , 2 (9), 805–814, doi:10.1038/s41893-019-0352-9.
Sae-Lim, P., A. Kause, H.A. Mulder and I. Olesen, 2017: Breeding and genetics symposium: climate change and selective breeding in aquaculture. J. Anim. Sci. , 95 (4), 1801–1812, doi:10.2527/jas.2016.1066.
Sah Akwen, N., 2017: Chapter 10 - “Not Migrating After All”: Young Farmers and Climate Change Adaptation in Cameroon. In: Beyond Agricultural Impacts[Zinyengere, N., T.F. Theodory, M. Gebreyes and C.I. Speranza(eds.)]. Academic Press, London, UK, pp. 193–220. ISBN 978-0128126240.
Sainger, M., et al., 2015: GM crops for developing world in the era of climate change: For increase of farmer’s income, poverty alleviation, nutrition and health. In: Genetic Manipulation in Plants for Mitigation of Climate Change. [Jaiwal, P., R. Singh and O. Dhankher (eds.)]. Springer, New Delhi.
Sainsbury, N.C., et al., 2019: The challenges of extending climate risk insurance to fisheries. Nat. Clim. Chang. , 9 (12), 896–897, doi:10.1038/s41558-019-0645-z.
Sainz, J.F., et al., 2019: Spatial planning of marine aquaculture under climate decadal variability: a case study for mussel farms in southern California. Front. Mar. Sci. , 6 (JUN), 253–253, doi:10.3389/fmars.2019.00253.
Saki, M., M. Tarkesh Esfahani and S. Soltani, 2018: A scenario-based modeling of climate change impacts on the aboveground net primary production in rangelands of central Iran. Environ Earth Sci, 77 (19), doi:10.1007/s12665-018-7864-x.
Sakkhamduang, J., K. Miwa and M. Mihara, 2017: Resin trees: A vital source of the Phnong people’s livelihood in transition in Cambodia. Satoyama Initiative Themat. Rev. , 3, 58.
Sala, E., et al., 2018: The economics of fishing the high seas. Sci. Adv. , 4 (6), eaat2504, doi:10.1126/sciadv.aat2504.
Salack, S., et al., 2016: Global warming induced hybrid rainy seasons in the Sahel. Environ. Res. Lett. , 11 (10), doi:10.1088/1748-9326/11/10/104008.
Salack, S., et al., 2015: Crop-climate ensemble scenarios to improve risk assessment and resilience in the semi-arid regions of West Africa. Clim. Res. , 65, 107–121, doi:10.3354/cr01282.
Salazar-Parra, C., et al., 2015: Carbon balance, partitioning and photosynthetic acclimation in fruit-bearing grapevine (Vitis vinifera L. cv. Tempranillo) grown under simulated climate change (elevated CO2, elevated temperature and moderate drought) scenarios in temperature gradient gre. J. Plant Physiol. , 174, 97–109.
Salazar, C., M. Jaime, Y. Figueroa and R. Fuentes, 2018: Innovation in small-scale aquaculture in Chile. Aquacult. Econ. Manag. , 22 (2), 151–167, doi:10.1080/13657305.2017.1409293.
Salick, J., Z.D. Fang and A. Byg, 2009: Eastern Himalayan alpine plant ecology, Tibetan ethnobotany, and climate change. Glob. Environ. Change. , 19 (2), 147–155, doi:10.1016/j.gloenvcha.2009.01.008.
Salih, A.A.M., M. Baraibar, K.K. Mwangi and G. Artan, 2020: Climate change and locust outbreak in East Africa. Nat. Clim. Chang. , 10 (7), 584–585, doi:10.1038/s41558-020-0835-8.
Sambaraju, K.R., A.L. Carroll and B.H. Aukema, 2019: Multiyear weather anomalies associated with range shifts by the mountain pine beetle preceding large epidemics. For. Ecol. Manag. , 438, 86–95, doi:10.1016/j.foreco.2019.02.011.
Samberg, L.H., et al., 2016: Subnational distribution of average farm size and smallholder contributions to global food production. Environ. Res. Lett. , 11 (12), doi:10.1088/1748-9326/11/12/124010.
Sánchez-Rodríguez, A.R., et al., 2019: Extreme flood events at higher temperatures exacerbate the loss of soil functionality and trace gas emissions in grassland. Soil Biol. Biochem. , 130, 227–236, doi:10.1016/j.soilbio.2018.12.021.
Sanchez, A.C., P.E. Osborne and N. Haq, 2011: Climate change and the African baobab (Adansonia digitata L.): the need for better conservation strategies. Afr. J. Ecol. , 49 (2), 234–245, doi:10.1111/j.1365-2028.2011.01257.x.
Sánchez, Y., A. M. Martínez-Graña, F. Santos-Francés and M. Yenes, 2019: Index for the calculation of future wine areas according to climate change application to the protected designation of origin “Sierra de Salamanca” (Spain). Ecol. Indic. , 107, doi:10.1016/j.ecolind.2019.105646. .
Sanderman, J., T. Hengl and G.J. Fiske, 2017: Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. U. S. A. , 114 (36), 9575–9580, doi:10.1073/pnas.1706103114.
Sanford, G.R., et al., 2021: Perenniality and diversity drive output stability and resilience in a 26-year cropping systems experiment. Field Crop. Res. , 263, doi:10.1016/j.fcr.2021.108071.
Sango, I. and N. Godwell, 2015: Climate change trends and environmental impacts in the Makonde Communal Lands, Zimbabwe. S. Afr. J. Sci. , 111 (7-8), 78–83.
Sanogo, K., et al., 2017: Farmers’ perceptions of climate change impacts on ecosystem services delivery of parklands in southern Mali. Agrofor. Syst. , 91 (2), 345–361, doi:10.1007/s10457-016-9933-z.
Sanogo, S., et al., 2015: Spatio-temporal characteristics of the recent rainfall recovery in West Africa. Int. J. Clim. , 35 (15), 4589–4605, doi:10.1002/joc.4309.
Santillan, D., et al., 2019: Vineyards in transition: a global assessment of the adaptation needs of grape producing regions under climate change. Sci. Total Environ. , 657, 839–852, doi:10.1016/j.scitotenv.2018.12.079.
Santo, R.E., et al., 2020: Considering plant-based meat substitutes and cell-based meats: a public health and food systems perspective. Front. Sustain. Food Syst. , 4, doi:10.3389/fsufs.2020.00134.
Santo, R.E., A. Palmer and B. Kim, 2016: Vacant Lots to Vibrant Plots: A review of the benefits and limitations of urban agriculture. Johns Hopkings University, , Baltimore, MD, United States, 35-35 pp..
Santos-Fita, D., E.J. Naranjo and J.L. Rangel-Salazar, 2012: Wildlife uses and hunting patterns in rural communities of the Yucatan Peninsula, Mexico. J Ethnobiol Ethnomed, 8 (1), 38, doi:10.1186/1746-4269-8-38.
Santos, J.A., R. Costa and H. Fraga, 2017: Climate change impacts on thermal growing conditions of main fruit species in Portugal. Clim Change, 140 (2), 273–286, doi:10.1007/s10584-016-1835-6.
Santoso, M.V., et al., 2021: A Nutrition-Sensitive Agroecology Intervention in Rural Tanzania Increases Children’s Dietary Diversity and Household Food Security But Does Not Change Child Anthropometry: Results from a Cluster-Randomized Trial. J. Nutr. , doi:10.1093/jn/nxab052.
SAPEA, 2020: A sustainable food system for the European Union. Evidence Review Report, No.7, SAPEA, Berlin, Germany, 221 pp. Available at: https://www.sapea.info/wp-content/uploads/sustainable-food-system-report.pdf (accessed Jaunary 6, 2022).
Sara, G., et al., 2018: Predicting shifting sustainability trade-offs in marine finfish aquaculture under climate change. Glob Chang Biol, 24 (8), 3654–3665, doi:10.1111/gcb.14296.
Sarmiento Barletti, J.P., A.M. Larson, C. Hewlett and D. Delgado, 2020: Designing for engagement: A Realist Synthesis Review of how context affects the outcomes of multi-stakeholder forums on land use and/or land-use change. World Dev, 127, doi:10.1016/j.worlddev.2019.104753.
Saronga, N.J., et al., 2016: “I eat two meals per day” impact of climate variability on eating habits among households in Rufiji district, Tanzania: a qualitative study. Agric. Food Secur. , 5 (1), 14, doi:10.1186/s40066-016-0064-6.
Satgé, F., et al., 2019: Unraveling the impacts of droughts and agricultural intensification on the Altiplano water resources. Agric For Meteorol, 279 (April), 107710–107710, doi:10.1016/j.agrformet.2019.107710.
Scanes, E., et al., 2020: Heatwaves alter survival of the Sydney rock oyster, Saccostrea glomerata. Mar. Pollut. Bull. , 158, 111389, doi:10.1016/j.marpolbul.2020.111389.
Scelza, B.A., D.W. Bird and R.B. Bird, 2014: Bush tucker, shop tucker: production, consumption, and diet at an Aboriginal outstation. Ecol. Food. Nutr. , 53 (1), 98–117, doi:10.1080/03670244.2013.772513.
Schaefer, A.M., M.D. Hanisak, M. McFarland and J.M. Sullivan, 2019: Integrated observing systems: An approach to studying harmful algal blooms in south Florida. J. Oper. Oceanogr. , 12 (sup2), 187–S198, doi:10.1080/1755876x.2019.1606879.
Schafleitner, R., 2016: Adaptation Of Potato (Solanum Tuberosum) and Tomato (S. Lycopersicum) to Climate Change. In: Applied Mathematics and Omics to Assess Crop Genetic Resources for Climate Change Adaptive Traits. [Bari, A., A. B. Damania, M. Mackay and S. Dayanandan (eds.)]. CRC Press, FL, USA. ISBN 9781315369006.
Schartup, A.T., et al., 2019: Climate change and overfishing increase neurotoxicant in marine predators. Nature, 572 (7771), 648–650, doi:10.1038/s41586-019-1468-9.
Schauberger, B., S. Rolinski, S. Schaphoff and C. Müller, 2019a: Global historical soybean and wheat yield loss estimates from ozone pollutino considering water and temperature as modifying effects. Agric For Meteorol, 265, 1–15.
Schauberger, G., et al., 2019b: Global warming impact on confined livestock in buildings: efficacy of adaptation measures to reduce heat stress for growing-fattening pigs. Clim Change, 156 (4), 567–587, doi:10.1007/s10584-019-02525-3.
Scheben, A., Y. Yuan and D. Edwards, 2016: Advances in genomics for adapting crops to climate change. Curr. Plant Biol. , 6, 2–10, doi:10.1016/j.cpb.2016.09.001.
Scheelbeek, P.F.D., et al., 2018: Effect of environmental changes on vegetable and legume yields and nutritional quality. Proc. Natl. Acad. Sci. U. S. A. , 115 (26), 6804–6809, doi:10.1073/pnas.1800442115.
Scheidel, A. and C. Work, 2018: Forest plantations and climate change discourses: New powers of ‘green’ grabbing in Cambodia. Land Use Policy, 77, 9–18, doi:10.1016/j.landusepol.2018.04.057.
Schelhaas, M.-J., et al., 2015: Alternative forest management strategies to account for climate change-induced productivity and species suitability changes in Europe. Reg Environ Change, 15 (8), 1581–1594, doi:10.1007/s10113-015-0788-z.
Schewe, J., et al., 2019: State-of-the-art global models underestimate impacts from climate extremes. Nat Commun, 10 (1), 1005, doi:10.1038/s41467-019-08745-6.
Schipanski, M. E., et al., 2016: Realizing Resilient Food Systems. BioScience, 66 (7), 600–610, doi:10.1093/biosci/biw052.
Schirpke, U., A. Alizinger, G. Leitinger and E. Tasser, 2019: Change from agricultural to touristic use: Effects on the aesthetic value of landscapes over the last 150 years. Landsc. Urban Plann, 187, 23–35, doi:10.1016/j.landurbplan.2019.03.004.
Schleicher, J., et al., 2019: Protecting half of the planet could directly affect over one billion people. Nat. Sustain. , 2 (12), 1094–1096, doi:10.1038/s41893-019-0423-y.
Schmidt, L.M., et al., 2019: The Biorefinery Approach. In: Energy from Organic Materials (Biomass) [Kaltschmitt, M. (ed.)]. Springer New York, pp. 1383-1412. ISBN 978-1-4939-7813-7.
Schmidt, M. and O. Pearson, 2016: Pastoral livelihoods under pressure: Ecological, political and socioeconomic transitions in Afar (Ethiopia). J. Arid Environ. , 124, 22–30, doi:10.1016/j.jaridenv.2015.07.003.
Schnurr, M., 2019: Africa’s gene revolution: Genetically modified crops and the future of african agriculture.
Schoneveld, G.C., 2017: Host country governance and the African land rush: 7 reasons why large-scale farmland investments fail to contribute to sustainable development. Geoforum, 83, 119–132, doi:10.1016/j.geoforum.2016.12.007.
Schott, S., et al., 2020: Operationalizing knowledge coevolution: towards a sustainable fishery for Nunavummiut. Arct. Sci. , 6 (3), 208–228, doi:10.1139/as-2019-0011.
Schreyer, F., C. Bunn and F. Castro-Llanos, 2018: The economic case for climate action in West-African cocoa production. Adapting cocoa production in Ghana to climate change is a smart investment. Inaction will result in income losses to farmers and the economy.
Schroeder, H. and N.C. Gonzalez, 2019: Bridging knowledge divides: the case of indigenous ontologies of territoriality and REDD. Policy Econ, 100, 198–206, doi:10.1016/j.forpol.2018.12.010.
Schroth, G., et al., 2015: Winner or loser of climate change? A modeling study of current and future climatic suitability of Arabica coffee in Indonesia. Reg Environ Change, 15 (7), 1473–1482, doi:10.1007/s10113-014-0713-x.
Schroth, G., et al., 2016: Vulnerability to climate change of cocoa in West Africa: Patterns, opportunities and limits to adaptation. Sci. Total Environ. , 556, 231–241, doi:10.1016/j.scitotenv.2016.03.024.
Schuldt, B., et al., 2020: A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. , 45, 86–103, doi:10.1016/j.baae.2020.04.003.
Schulte, L.A., et al., 2017: Prairie strips improve biodiversity and the delivery of multiple ecosystem services from corn-soybean croplands. Proc. Natl. Acad. Sci. U. S. A. , 114 (42), 11247–11252, doi:10.1073/pnas.1620229114.
Schultz, H.R. and G.V. Jones, 2010: Climate Induced Historic and Future Changes in Viticulture. J Wine Res, 21 (2-3), 137-145, doi:10.1080/09571264.2010.530098
Schulze-Sylvester, M. and A. Reineke, 2019: Elevated CO2 levels impact fitness traits of vine mealybug (Planococcus ficus Signoret), but not its parasitoid Leptomastix dactylopii Howard. Agronomy, 9, 326.
Schwendenmann, L., et al., 2010: Effects of an experimental drought on the functioning of a cacao agroforestry system, Sulawesi, Indonesia. Glob. Chang. Biol. , 16 (5), 1515–1530, doi:10.1111/j.1365-2486.2009.02034.x.
Scoville-Simonds, M., H. Jamali and M. Hufty, 2020: The Hazards of Mainstreaming: Climate change adaptation politics in three dimensions. World Dev, 125, 104683, doi:10.1016/j.worlddev.2019.104683.
Scurrah, M., R. Ccanto and M. Bonierbale, 2019: Participatory varietal selection in the Andes;Farmer involvement in selecting potatoes with traits from wild relatives. In: Farmers and Plant Breeding[Westengen, O.T. and T. Winge(eds.)]. Routledge, London. ISBN 978-0429507335.
Searchinger, T., B.M., B. Campbell, R. Carter and P. Thornton, 2019b: Food security and livelihoods of small-scale producers. Adapt Now: A Global Call for Leadership on Climate Resilience, GCA, GCA, Rotterdam. Available at: https://gca.org/wp-content/uploads/2019/09/GlobalCommission_Report_FINAL.pdf (accessed Juanuary 6, 2021).
Searchinger, T., et al., 2019: Creating a Sustainable Food Future. World Resources Institute, Washington, DC, 556–556.
Searchinger, T., et al., 2019a: Creating a sustainable food future: a menu of solutions to feed nearly 10 billion people by 2050. World Resources. Institute. Available At: Https://wrr-food.wri.org/.
Searchinger, T.D., S. Wirsenius, T. Beringer and P. Dumas, 2018: Assessing the efficiency of changes in land use for mitigating climate change. Nature, 564 (7735), 249–253, doi:10.1038/s41586-018-0757-z.
Secretan, P.A.D., et al., 2007: Guidelines to meet insurance and other risk management needs in developing aquaculture in Asia. FAO Fisheries Technical Paper. FAO, Rome, 53–78.
Seddon, N., et al., 2020: Understanding the value and limits of nature-based solutions to climate change and other global challenges. Philos. Trans. R. Soc. Lond. B Biol. Sci. , 375 (1794), 20190120, doi:10.1098/rstb.2019.0120.
Segnon, A., 2019: Exploring agrobiodiversity-based climate change adaptation in semi-arid areas of West Africa: a case study in Mali. University of Ghana, Accra, Ghana.
Segnon, A.C., E. G. Achigan-Dako, O.G. Gaoue and A. Ahanchede, 2015: Farmer’s Knowledge and Perception of Diversified Farming Systems in Sub-Humid and Semi-Arid Areas in Benin. Sustainability, 7 (6), 6573–6592, doi:10.3390/su7066573.
Seidel, P., 2014: Extreme weather and plant pests: extreme knowledge gap. Gesunde Pflanz. , 66, 93–101.
Seidl, R., et al., 2016: The sensitivity of current and future forest managers to climate-induced changes in ecological processes. Ambio, 45 (4), 430–441, doi:10.1007/s13280-015-0737-6.
Seidl, R., K. Albrich, D. Thom and W. Rammer, 2018: Harnessing landscape heterogeneity for managing future disturbance risks in forest ecosystems. J Environ Manage, 209, 46–56, doi:10.1016/j.jenvman.2017.12.014.
Seidl, R., et al., 2020: Globally consistent climate sensitivity of natural disturbances across boreal and temperate forest ecosystems. Ecography, 43 (7), 967–978, doi:10.1111/ecog.04995.
Sejian, V., et al., 2018: Review: Adaptation of animals to heat stress. Animal, 12 (s2), s431–s444, doi:10.1017/S1751731118001945.
Sekine, Y., 2021: Emerging ‘agrarian climate justice’ struggles in Myanmar. J Peasant Stud, doi:10.1080/03066150.2020.1839054.
Selby, J., 2019: Climate change and the Syrian civil war, Part II: the Jazira’s agrarian crisis. Geoforum, 101, 260–274, doi:10.1016/j.geoforum.2018.06.010.
Sellberg, M.M., A.V. Norström, G.D. Peterson and L.J. Gordon, 2020: Using local initiatives to envision sustainable and resilient food systems in the Stockholm city-region. Glob. Food Secur. , 24, 100334–100334, doi:10.1016/j.gfs.2019.100334.
Selmar, D. and M. Kleinwachter, 2013: Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind Crops Prod, 42, 558–566, doi:10.1016/j.indcrop.2012.06.020.
Semenza, J.C. and J.E. Suk, 2018: Vector-borne diseases and climate change: a European perspective. FEMS Microbiol. Lett. , 365 (2), doi:10.1093/femsle/fnx244.
Senapati, S. and V. Gupta, 2017: Vulnerability to Climate Change: Issues and Challenges towards Developing Vulnerability Indicator. In: Natural Resources Management [Information Resources Management Association (ed.)]. IGI global USA, pp. 1393-1416. ISBN 9781522508038.
Sendzimir, J., C.P. Reij and P. Magnuszewski, 2011: Rebuilding Resilience in the Sahel: Regreening in the Maradi and Zinder Regions of Niger. Ecol. Soc. , 16 (3), doi:10.5751/Es-04198-160301.
Seneviratne, S.I., X. Zhang, M. Adnan, W. Badi, C. Dereczynski, A. Di Luca, S. Ghosh, I. Iskandar, J. Kossin, S. Lewis, F. Otto, I. Pinto, M. Satoh, S.M. Vicente-Serrano, M. Wehner, and B. Zhou, 2021: Weather and Climate Extreme Events in a Changing Climate. In: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change[Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press.
Senf, C., J. Müller and R. Seidl, 2019: Post-disturbance recovery of forest cover and tree height differ with management in Central Europe. Landsc Ecol, 34 (12), 2837–2850, doi:10.1007/s10980-019-00921-9.
Serrasolses, G., et al., 2016: A Matter of taste: local explanations for the consumption of wild food pants in the Catalan pyrenees and the Balearic islands. Econ Bot , 70 (2), 176–189, doi:10.1007/s12231-016-9343-1.
Settele, J., J. Bishop and S.G. Potts, 2016: Climate change impacts on pollination. Nat. Plants, 2 (7), 16092, doi:10.1038/nplants.2016.92.
Settele, J., R. Scholes, R. Betts, S. Bunn, P. Leadley, D. Nepstad, J.T. Overpeck, and M. A. Taboada, 2014:Terrestrial and inland water systems. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability.Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[Field, C.B., V.R. Barros, D.J. Dokken, K.J. Mach, M.D. Mastrandrea, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L. White (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 271–359, 10.1017/CBO9781107415379.009.
Seufert, V. and N. Ramakutty, 2017: Many shades of gray - The context-dependent perfornance of organic agriculture. Sci. Adv. , 3 (3), e1602638, doi:10.1126/sciadv.1602638.
Seymour, F. and N.L. Harris, 2019: Reducing tropical deforestation. Science, 365 (6455), 756–757, doi:10.1126/science.aax8546.
Shackleton, R., C. Shackleton, S. Shackleton and J. Gambiza, 2013: Deagrarianisation and forest revegetation in a biodiversity hotspot on the Wild Coast, South Africa. Plos One, 8 (10), e76939, doi:10.1371/journal.pone.0076939.
Shaffer, L.J., 2014: Making sense of local climate change in rural Tanzania through knowledge co-production. J. Ethnobiol, 34 (3), 315–334, doi:10.2993/0278-0771-34.3.315.
Shaffril, H.A.M., A. Abu Samah and J.L. D’Silva, 2017: Climate change: Social adaptation strategies for fishermen. Mar. Policy. , 81, 256–261, doi:10.1016/j.marpol.2017.03.031.
Shaffril, H.A.M., S.E. Krauss and S.F. Samsuddin, 2018: A systematic review on Asian’s farmers’ adaptation practices towards climate change. Sci. Total Environ. , 644, 683–695, doi:10.1016/j.scitotenv.2018.06.349.
Shakhramanyan, N.G., U.A. Schneider and B.A. McCarl, 2013: US agricultural sector analysis on pesticide externalities - the impact of climate change and a Pigovian tax. Clim Change, 117 (4), 711–723, doi:10.1007/s10584-012-0585-3.
Shannak, S., D. Mabrey and M. Vittorio, 2018: Moving from theory to practice in the water–energy–food nexus: An evaluation of existing models and frameworks. Water-energy Nexus, 1 (1), 17–25, doi:10.1016/j.wen.2018.04.001.
Sharma, J., et al., 2017: Vulnerability of forests in India: a national scale assessment. Environ Manage, 60 (3), 544–553, doi:10.1007/s00267-017-0894-4.
Sharps, K., et al., 2020: Yield Constraint Score (YCS) for the Effect of Five Crop Stresses on Global Production of Four Staple Food Crops. NERC Environmental Information Data Centre, doi:10.5285/d347ed22-2b57-4dce-88e3-31a4d00d4358.
Shayegh, S., V. Manoussi and S. Dasgupta, 2020: Climate change and development in South Africa: the impact of rising temperatures on economic productivity and labour availability. Clim. Dev. , 1–11, doi:10.1080/17565529.2020.1857675.
Sheahan, M. and C.B. Barrett, 2017: Review: Food loss and waste in Sub-Saharan Africa. Food Policy, 70, 1–12, doi:10.1016/j.foodpol.2017.03.012.
Shen, T., H. Yu and Y.Z. Wang, 2021: Assessing the impacts of climate change and habitat suitability on the distribution and quality of medicinal plant using multiple information integration: Take Gentiana rigescens as an example. Ecol Indic, 123, 107376, doi:10.1016/j.ecolind.2021.107376.
Shen, Y.L. and X.G. Liu, 2015: Phenological changes of corn and soybeans over U.S. by bayesian change-point model. Sustainability, 7 (6), 6781–6803, doi:10.3390/su7066781.
Shepherd, C.J., O. Monroig and D.R. Tocher, 2017: Future availability of raw materials for salmon feeds and supply chain implications: The case of Scottish farmed salmon. Aquaculture, 467, 49–62, doi:10.1016/j.aquaculture.2016.08.021.
Shete, M. and M. Rutten, 2015: Impacts of large-scale farming on local communities’ food security and income levels - Empirical evidence from Oromia Region, Ethiopia. Land Use Policy, 47, 282–292, doi:10.1016/j.landusepol.2015.01.034.
Shi, W., et al., 2016: Ocean acidification increases cadmium accumulation in marine bivalves: a potential threat to seafood safety. Sci Rep, 6, 20197, doi:10.1038/srep20197.
Shiferaw, B., et al., 2014: Managing vulnerability to drought and enhancing livelihood resilience in sub-Saharan Africa: Technological, institutional and policy options. Weather. Clim. Extrem. , 3, 67–79, doi:10.1016/j.wace.2014.04.004.
Shimoda, S., H. Kanno and T. Hirota, 2018: Time series analysis of temperature and rainfall-based weather aggregation reveals significant correlations between climate turning points and potato (Solanum tuberosum L) yield trends in Japan. Agric For Meteorol, 263, 147–155, doi:10.1016/j.agrformet.2018.08.005.
Shindell, D., G. Faluvegi, P. Kasibhatla and R. Van Dingenen, 2019: Spatial patterns of crop yield change by emitted pollutant. Earths Future, 7 (2), 101–112, doi:10.1029/2018EF001030.
Shindell, D.T., 2016: Crop yield changes induced by emissions of individual climate-altering pollutants. Earths Future, 4 (8), 373–380, doi:10.1002/2016ef000377.
Shukla, R., et al., 2019: Climate change perception: an analysis of climate change and risk perceptions among farmer types of Indian Western Himalayas. Clim Change, 152 (1), 103–119, doi:10.1007/s10584-018-2314-z.
Sibhatu, K.T. and M. Qaim, 2018: Review: Meta-analysis of the association between production diversity, diets, and nutrition in smallholder farm households. Food Policy, 77, 1–18, doi:10.1016/j.foodpol.2018.04.013.
Sida, T.S., F. Baudron, H. Kim and K.E. Giller, 2018: Climate-smart agroforestry: Faidherbia albida trees buffer wheat against climatic extremes in the Central Rift Valley of Ethiopia. Agric. For. Meteorol. , 248, 339–347, doi:10.1016/j.agrformet.2017.10.013.
Siegner, A., J. Sowerwine and C. Acey, 2018: Does Urban Agriculture Improve Food Security? Examining the Nexus of Food Access and Distribution of Urban Produced Foods in the United States: A Systematic Review. Sustainability, 10 (9), 8–12, doi:10.3390/su10092988.
Silanikove, N. and N. Koluman, 2015: Impact of climate change on the dairy industry in temperate zones: Predications on the overall negative impact and on the positive role of dairy goats in adaptation to earth warming. Small. Rumin. Res. , 123 (1), 27–34, doi:10.1016/j.smallrumres.2014.11.005.
Silas, M.O., et al., 2020: Adaptive capacity and coping strategies of small-scale coastal fisheries to declining fish catches: insights from Tanzanian communities. Environ. Sci. Policy, 108, 67–76, doi:10.1016/j.envsci.2020.03.012.
Silva, J.G., et al., 2020: Are medicinal plants an alternative to the use of synthetic pharmaceuticals in animal healthcare in the Brazilian semi-arid?Ethnobot. Res. Appl. , 19, 1–20.
Silwal, P., L. Roberts, H.G. Rennie and M.J. Lexer, 2019: Adapting to climate change: an assessment of local adaptation planning processes in forest-based communities in Nepal. Clim. Dev. , 11 (10), 886–898, doi:10.1080/17565529.2019.1586634.
Simelton, E., B.V. Dam and D. Catacutan, 2015: Trees and agroforestry for coping with extreme weather events: experiences from northern and central Viet Nam. Agrofor. Syst. , 89 (6), 1065–1082, doi:10.1007/s10457-015-9835-5.
Sinclair, F., et al., 2019: The Contribution of Agroecological Approaches to Realizing Climate-Resilient Agriculture. Global Center on Adaptaion, . Available at: https://gca.org/reports/the-contributions-of-agroecological-approaches-to-realizing-climate-resilient-agriculture/ (accessed January 6, 2021 ).
Singano, C.D., et al., 2020: What does global warming mean for stored-grain protection? Options for Prostephanus truncatus (Horn) control at increased temperatures. J Stored Prod Res, 85, 101532–101532, doi:10.1016/j.jspr.2019.101532.
Singh, C., 2018: Is participatory watershed development building local adaptive capacity? Findings from a case study in Rajasthan, India. Environ. Dev. , 25, 43–58, doi:10.1016/j.envdev.2017.11.004.
Singh, C. and R. Basu, 2020: Moving in and out of vulnerability: Interrogating migration as an adaptation strategy along a rural-urban continuum in India. Geogr J, 186 (1), 87–102, doi:10.1111/geoj.12328.
Singh, G.G., N. Hilmi and J.R. Bernhardt, 2019: Climate impacts on the ocean are making the sustainable development goals a moving target travelling away from us. People Nat. , 1 (3), 317–330, doi:10.1002/pan3.26.
Singh, K., et al., 2017: Mapping regional risks from climate change for rainfed rice cultivation in India. Agric. Syst. , 156, 76–84, doi:10.1016/j.agsy.2017.05.009.
Singh, P.K. and H. Chudasama, 2021: Pathways for climate change adaptations in arid and semi-arid regions. J Clean Prod, 284, 124744–124744, doi:10.1016/J.JCLEPRO.2020.124744.
Singh, R.B., 2000: Environmental consequences of agricultural development: a case study from the Green Revolution state of Haryana, India. Agric Ecosyst Environ, 82 (1-3), 97–103, doi:10.1016/S0167-8809(00)00219-X.
Singh, R.P., et al., 2020: Varietal replacement rate: Prospects and challenges for global food security. Glob. Food Secur. Policy Econ. Environ. , 25, doi:10.1016/j.gfs.2019.100324.
Sissoko, K., et al., 2011: Agriculture, livelihoods and climate change in teh West African Sahel. Reg Environ Change, 11, 119–125.
Sita, K., et al., 2017: Food Legumes and Rising Temperatures: Effects, Adaptive Functional Mechanisms Specific to Reproductive Growth Stage and Strategies to Improve Heat Tolerance. Front. Plant Sci. , 8, 1658, doi:10.3389/fpls.2017.01658.
Skibiel, A.L., et al., 2018: In Utero Heat Stress Alters the Offspring Epigenome. Sci Rep, 8 (1), 14609, doi:10.1038/s41598-018-32975-1.
Slenning, B.D., 2010: Global climate change and implications for disease emergence. Vet. Pathol. , 47 (1), 28–33, doi:10.1177/0300985809354465.
Sloat, L.L., et al., 2018: Increasing importance of precipitation variability on global livestock grazing lands. Nat. Clim. Chang. , 8 (3), 214, doi:10.1038/s41558-018-0081-5.
Smalås, A., et al., 2019: Climate warming is predicted to enhance the negative effects of harvesting on high-latitude lake fish. J Appl Ecol, 57 (2), 270–282, doi:10.1111/1365-2664.13535.
Smale, D.A., et al., 2019: Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Chang. , 9 (4), 306, doi:10.1038/s41558-019-0412-1.
Small, G.E., R. McDougall and G.S. Metson, 2019: Would a sustainable city be self-sufficient in food production?Int. J. Des. Nat. Ecodynamics, 14 (3), 178–194, doi:10.2495/dne-v14-n3-178-194.
Smith, E., et al., 2019a: Contribution of wild foods to diet, food security, and cultural values amidst climate change. J. Agric. Food Syst. Community Dev. , 9 (B), 191–214, doi:10.5304/jafscd.2019.09B.011.
Smith, L.C. and L. Haddad, 2015: Reducing Child Undernutrition: Past Drivers and Priorities for the Post-MDG Era. World Dev, 68, 180–204, doi:10.1016/j.worlddev.2014.11.014.
Smith, L.G., G.J.D. Kirk, P.J. Jones and A.G. Williams, 2019b: The greenhouse gas impacts of converting food production in England and Wales to organic methods. Nat Commun, 10 (1), 4641, doi:10.1038/s41467-019-12622-7.
Smith, M. E. and S. Bernard, 2020: Satellite ocean color based harmful algal bloom indicators for aquaculture decision support in the southern Benguela. Front. Mar. Sci. , 7, doi:10.3389/fmars.2020.00061.
Smith, M.R. and S.S. Myers, 2018: Impact of anthropogenic CO2 emissions on global human nutrition. Nat. Clim. Chang. , 8 (9), 834, doi:10.1038/s41558-018-0253-3.
Smith, P., et al., 2020a: Which practices co-deliver food security, climate change mitigation and adaptation, and combat land degradation and desertification?Glob Chang Biol, 26 (3), 1532–1575, doi:10.1111/gcb.14878.
Smith, P., J. Nkem, K. Calvin, D. Campbell, F. Cherubini, G. Grassi, V. Korotkov, A.L. Hoang, S. Lwasa, P. McElwee, E. Nkonya, N. Saigusa, J.-F. Soussana, M. A. Taboada, 2019c: Interlinkages Between Desertification, Land Degradation, Food Security and Greenhouse Gas Fluxes: Synergies, Trade-offs and Integrated Response Options. In: Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems[P.R. Shukla, J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H.- O. Portner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi, J. Malley, (eds.). In press.
Smith, P., et al., 2018: Impacts on terrestrial biodiversity of moving from a 2 C to a 1.5 C target. Philos. Trans. Royal Soc. A: Math. Phys. Eng. Sci. , 376 (2119), 20160456.
Smith, R.A.J. and K. Rhiney, 2016: Climate (in)justice, vulnerability and livelihoods in the Caribbean: The case of the indigenous Caribs in northeastern St. Vincent. Geoforum, 73, 22–31, doi:10.1016/j.geoforum.2015.11.008.
Smith, S.D., et al., 2000: Elevated CO2 increases productivity and invasive species success in an arid ecosystem. Nature, 408 (6808), 79–82, doi:10.1038/35040544.
Smith, W., et al., 2020b: Towards an improved methodology for modelling climate change impacts on cropping systems in cool climates. Sci. Total Environ. , 728, 138845, doi:10.1016/j.scitotenv.2020.138845.
Snapp, S.S., C.M. Cox and B.G. Peter, 2019: Multipurpose legumes for smallholders in sub-Saharan Africa: Identification of promising ‘scale out’ options. Glob. Food Secur. Policy Econ. Environ. , 23, 22–32, doi:10.1016/j.gfs.2019.03.002.
Snapp, S.S., et al., 2021: Agroecology and climate change rapid evidence review: Performance of agroecological approaches in low- and middle- income countries. CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS), CGIAR, Wageningen, the Netherlands, 63-63 pp. Available at: https://hdl.handle.net/10568/113487 (accessed 31/8/2021).
Sohn, J.A., S. Saha and J. Bauhus, 2016: Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. , 380, 261–273, doi:10.1016/j.foreco.2016.07.046.
Sohngen, B., R. Mendelsohn and R. Sedjo, 2001: A global model of climate change impacts on timber markets. J. Agric. Resour. Econ. , 26 (2), 326–343.
Solecki, W., M. Pelling and M. Garschagen, 2017: Transitions between risk managemetn regimes in cities. Ecol. Soc. , 22 (2), doi:10.5751/es-09102-220238
Sommerfeld, A., et al., 2018: Patterns and drivers of recent disturbances across the temperate forest biome. Nat Commun, 9 (1), 4355, doi:10.1038/s41467-018-06788-9Sonenshine, D.E., 2018: Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int J Environ Res Public Health, 15 (3), 478, doi:10.3390/ijerph15030478.
Song, Y., et al., 2019: A 20-year journey: Participatory breeding of maize in South-West China. In: Farmers and Plant Breeding[Westengen, O. T. and T. Winge (eds.)]. Routledge, London and New York, pp. 119-134.Sonwa, D.J., et al., 2017a: Drivers of climate risk in African agriculture. Clim. Dev. , 9 (5), 383–398, doi:10.1080/17565529.2016.1167659.
Sonwa, D.J., et al., 2017b: Structure and composition of cocoa agroforests in the humid forest zone of Southern Cameroon. Agrofor. Syst. , 91 (3), 451–470, doi:10.1007/s10457-016-9942-y.
Sorensen, N.N., A.D. Lassen, H. Loje and I. Tetens, 2015: The Danish Organic Action Plan 2020: assessment method and baseline status of organic procurement in public kitchens. Public Health Nutr. , 18 (13), 2350–2357, doi:10.1017/S1368980015001421.
Soroye, P., T. Newbold and J. Kerr, 2020: Climate change contributes to widespread declines among bumble bees across continents. Science, 367 (6478), 685–688, doi:10.1126/science.aax8591.
Sotelo, S., et al., 2020: Pronosticos AClimateColombia: A system for the provision of information for climate risk reduction in Colombia. Comput. Electron. Agric. , 174, doi:10.1016/j.compag.2020.105486.
Soto, D., et al., 2019: Salmon farming vulnerability to climate change in southern Chile: understanding the biophysical, socioeconomic and governance links. Rev. Aquac. , 11 (2), 354–374, doi:10.1111/raq.12336.
Soto, D., et al., 2021: Scientific warnings could help to reduce farmed salmon mortality due to harmful algal blooms. Mar. Policy, 132, 104705, doi:10.1016/j.marpol.2021.104705.
Soto, D., et al., 2018: Chapter 21: Climate change and aquaculture: vulnerability and adaptation options. In: Impacts of climate change on fisheries and aquaculture: synthesis of current knowledge, adaptation and mitigation options[Barange, M., et al.(ed.)]. FAO, Rome, pp. 465–490.
Soto Golcher, C. and I.J. Visseren-Hamakers, 2018: Framing and integration in the global forest, agriculture and climate change nexus. Environ. Plan. C: Polit. Space, 36 (8), 1415–1436, doi:10.1177/2399654418788566.
Soumya, K.V., C.M. Shackleton and S.R. Setty, 2019: Impacts of gum-resin harvest and Lantana camara invasion on the population structure and dynamics of Boswellia serrata in the Western Ghats. India. for. Ecol. Manag. , 453, 117618, doi:10.1016/j.foreco.2019.117618.
Sousa-Silva, R., et al., 2016: Adaptation of forest management to climate change as perceived by forest owners and managers in Belgium. For. Ecosyst. , 3 (1), 22, doi:10.1186/s40663-016-0082-7.
Sousa-Silva, R., et al., 2018: Adapting forest management to climate change in Europe: Linking perceptions to adaptive responses. Policy Econ, 90, 22–30, doi:10.1016/j.forpol.2018.01.004.
Souther, S. and J.B. McGraw, 2014: Synergistic effects of climate change and harvest on extinction risk of American ginseng. Ecol Appl, 24 (6), 1463–1477, doi:10.1890/13-0653.1.
Sovacool, B.K., 2021: Who are the victims of low-carbon transitions? Towards a political ecology of climate change mitigation. Energy Res. Soc. Sci. , 73, 101916, doi:10.1016/j.erss.2021.101916.
Sovacool, B.K., B.O. Linner and M. E. Goodsite, 2015: COMMENTARY: The political economy of climate adaptation. Nat. Clim. Chang. , 5 (7), 616–618, doi:10.1038/nclimate2665.
Sowman, M. and S. Raemaekers, 2018: Socio-ecological vulnerability assessment in coastal communities in the BCLME region. J. Mar. Syst. , 188, 160–171, doi:10.1016/j.jmarsys.2018.01.008.
Specht, K., et al., 2019: How will we eat and produce in the cities of the future? From edible insects to vertical farming—A study on the perception and acceptability of new approaches. Sustainability, 11 (16), 1–22, doi:10.3390/su11164315.
Spellman, K.V. and N.Y. Swenson, 2012: Assessing the vulnerability of Western Alaska ecosystems and subsistence resources to nonnative plant invasion. Report for University of Alaska Fairbanks Contribution to Western Alaska LCC Cooperative Project, Vol. 23. University of Alaska Fairbanks, Alaska, United States.
Spijkers, J. and W.J. Boonstra, 2017: Environmental change and social conflict: the northeast Atlantic mackerel dispute. Reg Environ Change, 17 (6), 1835–1851, doi:10.1007/s10113-017-1150-4.
Spijkers, J., et al., 2018: Marine fisheries and future ocean conflict. Fish. Fish. , 19 (5), 798–806, doi:10.1111/faf.12291.
Spires, M., S. Shackleton and G. Cundill, 2014: Barriers to implementing planned community-based adaptation in developing countries: a systematic literature review. Clim. Dev. , 6 (3), 277–287, doi:10.1080/17565529.2014.886995.
Springmann, M., et al., 2018: Options for keeping the food system within environmental limits. Nature, 562 (7728), 519–525, doi:10.1038/s41586-018-0594-0.
Springmann, M., et al., 2016: Global and regional health effects of future food production under climate change: a modelling study. Lancet , 387 (10031), 1937–1946, doi:10.1016/S0140-6736(15)01156-3.
Srivastava, P., R. Singh, S. Tripathi and A.S. Raghubanshi, 2016: An urgent need for sustainable thinking in agriculture - An Indian scenario. Ecol Indic, 67, 611–622, doi:10.1016/j.ecolind.2016.03.015.
Ssegane, H. and M.C. Negri, 2016: An Integrated Landscape Designed for Commodity and Bioenergy Crops for a Tile-Drained Agricultural Watershed. J. environ. qual. , 45 (5), 1588–1596, doi:10.2134/jeq2015.10.0518.
Ssegane, H., M.C. Negri, J. Quinn and M. Urgun-Demirtas, 2015: Multifunctional landscapes: Site characterization and field-scale design to incorporate biomass production into an agricultural system. Biomass Bioenergy, 80, 179–190, doi:10.1016/j.biombioe.2015.04.012.
Stanimirova, R., et al., 2019: Sensitivity of Global Pasturelands to Climate Variation. Earths Future, 7 (12), 1353–1366, doi:10.1029/2019ef001316.
Stark, F., et al., 2018: Crop-livestock integration determines the agroecological performance of mixed farming systems in Latino-Caribbean farms. Agron. Sustain. Dev. , 38 (1), 4–4, doi:10.1007/s13593-017-0479-x.
Stefan, L., et al., 2021: Positive effects of crop diversity on productivity driven by changes in soil microbial composition. Front. Microbiol. , 12, 808–808.
Steffen, W., et al., 2015: Sustainability. Planetary boundaries: guiding human development on a changing planet. Science, 347 (6223), 1259855, doi:10.1126/science.1259855.
Stehfest, K.M., et al., 2017: Response of Atlantic salmon Salmo salar to temperature and dissolved oxygen extremes established using animal-borne environmental sensors. Sci Rep, 7 (1), 4545, doi:10.1038/s41598-017-04806-2.
Steiner, A., et al., 2020: Actions to transform food systems under climate change. CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS), Wageningen, the Netherlands.
Stemkovski, M., et al., 2020: Bee phenology is predicted by climatic variation and functional traits. Ecol Lett , 23 (11), 1589–1598, doi:10.1111/ele.13583.
Stephen, K., 2018: Societal Impacts of a Rapidly Changing Arctic. Curr Clim Change Rep, 4 (3), 223–237, doi:10.1007/s40641-018-0106-1.
Stevens-Rumann, C.S. and P. Morgan, 2019: Tree regeneration following wildfires in the western US: a review. Fire Ecol. , 15 (1), doi:10.1186/s42408-019-0032-1.
Steward, P.R., C. Thierfelder, A.J. Dougill and I. Ligowe, 2019: Conservation agriculture enhances resistance of maize to climate stress in a Malawian medium-term trial. Agric. Ecosyst. Environ. , 277, 95–104, doi:10.1016/J.AGEE.2018.07.009.
Stewart-Koster, B., et al., 2017: Expert based model building to quantify risk factors in a combined aquaculture-agriculture system. Agric Syst , 157, 230–240, doi:10.1016/j.agsy.2017.08.001.
Stewart-Sinclair, P.J., K.S. Last, B.L. Payne and T.A. Wilding, 2020: A global assessment of the vulnerability of shellfish aquaculture to climate change and ocean acidification. Ecol Evol, 10 (7), 3518–3534, doi:10.1002/ece3.6149.
Stien, A., et al., 2012: Congruent responses to weather variability in high arctic herbivores. Biol. Lett. , 8 (6), 1002–1005, doi:10.1098/rsbl.2012.0764.
Stocker, T.F., D. Qin, G.-K. Plattner, L.V. Alexander, S.K. Allen, N.L. Bindoff, F.-M. Bréon, J.A. Church, U. Cubasch, S. Emori, P. Forster, P. Friedlingstein, N. Gillett, J.M. Gregory, D.L. Hartmann, E. Jansen, B. Kirtman, R. Knutti, K. Krishna Kumar, P. Lemke, J. Marotzke, V. Masson-Delmotte, G.A. Meehl, I.I. Mokhov, S. Piao, V. Ramaswamy, D.Randall, M. Rhein, M. Rojas, C. Sabine, D. Shindell, L.D. Talley, D.G. Vaughan and S.-P. Xie, 2013: Technical Summary. In: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[Stocker, T. F., D. Qin, G. K. Plattner, M. Tignor, S. K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex and P. M. Midgley (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 33-115. ISBN 9781107057991.
Stoeckl, N., et al., 2017: Socioeconomic Impacts of Changes to Marine Fisheries and Aquaculture that are Brought About Through Climate Change. In: Climate Change Impacts on Fisheries and Aquaculture. John Wiley & Sons, Ltd, Chichester, UK, pp. 925–958. ISBN 978-1119154051.
Stoltz, A.D., M. Shivlani and R. Glazer, 2021: Fishing Industry Perspectives on Sea-Level Rise Risk and Adaptation. Water, 13 (8), 1124, doi:10.3390/w13081124.
Stone, G., 2017: Dreading CRISPR: GMO’s, honest brokers, and mertonian transgressions. Geogr Rev, 107, 584–591.
Stone, G.D. and D. Glover, 2017: Disembedding grain: Golden Rice, the Green Revolution, and heirloom seeds in the Philippines. Agric Hum Values, 34 (1), 87–102, doi:10.1007/s10460-016-9696-1.
Stoorvogel, J.J., M. Bakkenes, B.J.E. Brink and A.J.A.M. Temme, 2017: To what extent did we change our soils? A global comparison of natural and current conditions. Land Degrad. Dev. , 28 (7), 1982–1991, doi:10.1002/ldr.2721.
Stranden, I., et al., 2019: Genomic selection strategies for breeding adaptation and production in dairy cattle under climate change. Hered. (edinb) , 123 (3), 307–317, doi:10.1038/s41437-019-0207-1.
Stringer, L.C., et al., 2020: Adaptation and development pathways for different types of farmers. Environ Sci Policy, 104, 174–189, doi:10.1016/j.envsci.2019.10.007.
Stuart-Smith, J., et al., 2018: Southernmost records of two Seriola species in an Australian ocean-warming hotspot. Mar. Biodivers. , 48 (3), 1579–1582, doi:10.1007/s12526-016-0580-4.
Styles, D., et al., 2016: Climate regulation, energy provisioning and water purification: Quantifying ecosystem service delivery of bioenergy willow grown on riparian buffer zones using life cycle assessment. Ambio, 45 (8), 872–884, doi:10.1007/s13280-016-0790-9.
Sudo, K., K. Watanabe, N. Yotsukura and M. Nakaoka, 2020: Predictions of kelp distribution shifts along the northern coast of Japan. Ecol. Res. , 35 (1), 47–60, doi:10.1111/1440-1703.12053.
Sugden, F., et al., 2014: Agrarian stress and climate change in the Eastern Gangetic Plains: Gendered vulnerability in a stratified social formation. Glob. Environ. Change. , 29, 258–269, doi:10.1016/j.gloenvcha.2014.10.008.
Sugiura, T., 2019: Three climate change adaptation strageties for fruit production In: Climate Smart Agriculture for the Small-Scale Farmers in the Asian and Pacific Region. In: National Agriculture and Food Research Organization (NARO) Tsukuba[Shirato, Y. and A. Hasebe(eds.)]. , NARO, Japan. ISBN 978-4908914027 pp. 277).
Sugiura, T., H. Ogawa, N. Fukuda and T. Moriguchi, 2013: Changes in the taste and textural attributes of apples in response to climate change. Sci. Rep. , 3, 2418, doi:10.1038/srep02418.
Sugiura, T., M. Shiraishi, S. Konno and A. Sato, 2018: Prediction of Skin Coloration of Grape Berries from Air Temperature. Hortic. J. , 87 (1), 18–25, doi:10.2503/hortj.OKD-061.
Suhl, J., et al., 2018: An innovative suction filter device reduces nitrogen loss in double recirculating aquaponic systems. Aquac Eng, 82, 63–72, doi:10.1016/j.aquaeng.2018.06.008.
Sullivan, J. and B. Rosenberg, 2018: “Losing Your Land You Feel Like You’re Losing Your Identity, Like You’re Experiencing Slow Death”: An Interview With Chief Thomas Dardar-Houma Nation. New. Solut. , 28 (3), 501–514, doi:10.1177/1048291118795155.
Sullivan, M.J.P., et al., 2020: Long-term thermal sensitivity of Earth’s tropical forests. Science, 368 (6493), 869, doi:10.1126/science.aaw7578.
Sulser, T., et al., 2021a: Climate change and hunger: Estimating costs of adaptation in the agrifood systemFood policy report. International Food Policy Research Institute (IFPRI), Washington, DC.
Sulser, T.B., et al., 2021b: Disability-adjusted life years due to chronic and hidden hunger under food system evolution with climate change and adaptation to 2050. Am J Clin Nutr, doi:10.1093/ajcn/nqab101.
Sultan, B., D. Defrance and T. Iizumi, 2019: Evidence of crop production losses in West Africa due to historical global warming in two crop models. Sci. Rep. , 9 (1), 12834, doi:10.1038/s41598-019-49167-0.
Sultan, B. and M. Gaetani, 2016: Agriculture in west Africa in the twenty-first century: climate change and impacts scenarios, and potential for adaptation. Front. Plant Sci. , 7, 1262, doi:10.3389/fpls.2016.01262.
Sultan, B., et al., 2020: Current needs for climate services in West Africa: Results from two stakeholder surveys. Clim. Serv. , 18, 100166–100166, doi:10.1016/j.cliser.2020.100166.
Sultana, M.N., M.M. Uddin, B.G. Ridoutt and K.J. Peters, 2014: Comparison of water use in global milk production for different typical farms. Agric Syst , 129, 9–21, doi:10.1016/j.agsy.2014.05.002.
Sumaila, U.R., et al., 2019: Benefits of the Paris Agreement to ocean life, economies, and people. Sci. Adv. , 5 (2), eaau3855, doi:10.1126/sciadv.aau3855.
Sumaila, U.R., et al., 2020: Illicit trade in marine fish catch and its effects on ecosystems and people worldwide. Sci. Adv. , 6 (9), eaaz3801, doi:10.1126/sciadv.aaz3801.
Sumby, J., M. Haward, E.A. Fulton and G.T. Pecl, 2021: Hot fish: The response to climate change by regional fisheries bodies. Mar. Policy. , 123, 104284, doi:10.1016/j.marpol.2020.104284.
Sun, Q., et al., 2019: Global heat stress on health, wildfires, and agricultural crops under different levels of climate warming. Environ Int , 128, 125–136, doi:10.1016/j.envint.2019.04.025.
Sun, T., et al., 2021: Current rice models underestimate yield losses from short-term heat stresses. Glob. Chang. Biol. , 27 (2), 402–416.
Sun, W., et al., 2020: Climate drives global soil carbon sequestration and crop yield changes under conservation agriculture. Glob Chang Biol, 26 (6), 3325–3335, doi:10.1111/gcb.15001.
Sunderlin, W.D., A.M. Larson and J.P.S. Barletti, 2018: Land and carbon tenure. In: Transforming REDD+: Lessons and new directions[Angelsen, A., et al.(ed.)]. CIFOR, Bogor, Indonesia, pp. 93–102.
Susaeta, A. and P. Lal, 2018: Impacts of climate change and bioenergy markets on the profitability of slash pine pulpwood production in the southeastern United States. Forests, 9 (10), 656, doi:10.3390/f9100656.
Sweetman, C., et al., 2014: Metabolic effects of elevated temperature on organic acid degradation in ripening Vitis vinifera fruit. J. Exp. Bot. , 65 (20), 5975–5988, doi:10.1093/jxb/eru343.
Swinburn, B.A., et al., 2019: The global syndemic of obesity, undernutrition, and cliamte change: The Lancet Commission report. Lancet , 393, 791–846.
Sylvester, O. and M. Little, 2020: “I came all this way to receive training, am I really going to be taught by a woman?” Factors that support and hinder women’s participation in agroecology in Costa Rica. Agroecol. Sustain. Food Syst. , 1–24, doi:10.1080/21683565.2020.1811830.
Szewczyk, W., J.C.C. Martinez, I. Mongelli and A.S. Ramirez, 2018: JRC PESETA III Project: Economic integration and spillover analysis. Joint Research Centre (Seville site), https://ideas.repec.org/p/ipt/iptwpa/jrc113810.html.
Szopa, S., V. Naik, B. Adhikary, P. Artaxo, T. Berntsen, W.D. Collins, S. Fuzzi, L. Gallardo, A. Kiendler Scharr, Z. Klimont, H. Liao, N. Unger, and P. Zanis, 2021: Short-Lived Climate Forcers. In: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change[Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press.
Tadesse, G., B. Algieri, M. Kalkuhl and J. von Braun, 2014: Drivers and triggers of international food price spikes and volatility. Food Policy, 47, 117–128, doi:10.1016/j.foodpol.2013.08.014.
Tahvanainen, V., J. Miina, M. Kurttila and K. Salo, 2016: Modelling the yields of marketed mushrooms in Picea abies stands in eastern Finland. For. Ecol. Manag. , 362, 79–88, doi:10.1016/j.foreco.2015.11.040.
Taylor, K., 2015: Improving substantive and procedural protections for indigenous rights in REDD projects: Possible lessons from Brazil. J. Sustain. Dev. Law Pol. , 5, 32-54.
Tambo, J.A. and T. Abdoulaye, 2013: Smallholder farmers’ perceptions of and adaptations to climate change in the Nigerian savanna. Reg Environ Change, 13 (2), 375–388, doi:10.1007/s10113-012-0351-0.
Tamburini, G., et al., 2020: Agricultural diversification promotes multiple ecosystem services without compromising yield. Sci. Adv. , 6 (45), eaba1715–eaba1715, doi:10.1126/sciadv.aba1715.
Tamene, L., et al., 2019: Soil structural degradation and nutrient limitations across land use categories and climatic zones in Southern Africa. Land Degrad. Dev. , 30 (11), 1288–1299, doi:10.1002/ldr.3302.
Tamm, O., S. Maasikamae, A. Padari and T. Tamm, 2018: Modelling the effects of land use and climate change on the water resources in the eastern Baltic Sea region using the SWAT model. CATENA, 167, 78–89, doi:10.1016/j.catena.2018.04.029.
Tan, K. and H. Zheng, 2020: Ocean acidification and adaptive bivalve farming. Sci. Total Environ. , 701, 134794, doi:10.1016/j.scitotenv.2019.134794.
Tan, Q., Y. Liu, L. Dai and T. Pan, 2021: Shortened key growth periods of soybean observed in China under climate change. Sci Rep, 11 (1), 8197, doi:10.1038/s41598-021-87618-9.
Tankari, M.R., 2020: Rainfall variability and farm households’ food insecurity in Burkina Faso: nonfarm activities as a coping strategy. Food Secur, 12 (3), 567–578, doi:10.1007/s12571-019-01002-0.
Tanner, T., et al., 2014: Political economy of climate compatible development: artisanal fisheries and climate change in Ghana. Ids Work. Pap. , 2014 (446), 1–30, doi:10.1111/j.2040-0209.2014.00446.x.
Tao, F., et al., 2020: Why do crop models diverge substantially in climate impact projections? A comprehensive analysis based on eight barley crop models. Agric For Meteorol, 281, 107851.
Tao, F., et al., 2017: Designing future barley ideotypes using a crop model ensemble. Eur J Agron, 82, 144–162, doi:10.1016/j.eja.2016.10.012.
Tao, F., et al., 2016: Historical data provide new insights into response and adaptation of maize production systems to climate change/variability in China. Field Crop. Res. , 185, 1–11, doi:10.1016/j.fcr.2015.10.013.
Tarchiani, V., et al., 2017: Smallholder Farmers Facing Climate Change in West Africa: Decision-Making between Innovation and Tradition. J. Innov. Econ. , 3 (24), 151–176, doi:10.3917/jie.pr1.0013.
Taremwa, N.K., D. Gashumba, A. Butera and T. Ranganathan, 2017: Climate Change Adaptation in Rwanda through Indigenous Knowledge Practice. J. Soc. Sci. , 46, 165–175.
Tate, R.D., K. Benkendorff, A. Lah and B.P. Kelaher, 2017: Ocean acidification and warming impacts the nutritional properties of the predatory whelk, Dicathais orbita. J. Exp. Mar. Biol. Ecol. , 493, 7–13, doi:10.1016/j.jembe.2017.03.006.
Tavenner, K., et al., 2019: Intensifying Inequality? Gendered Trends in Commercializing and Diversifying Smallholder Farming Systems in East Africa. Front. Sustain. Food Syst. , 3, doi:10.3389/fsufs.2019.00010.
Taye, Z.M., et al., 2016: Meteorological conditions and site characteristics driving edible mushroom production in Pinus pinaster forests of Central Spain. Fungal Ecol, 23, 30–41, doi:10.1016/j.funeco.2016.05.008.
Taylor, D., et al., 2016: Environmental change and Rift Valley fever in eastern Africa: projecting beyond HEALTHY FUTURES. Geospat Health, 11 (1 Suppl), 387, doi:10.4081/gh.2016.387.
Taylor, M., 2016: Risky Ventures: Financial Inclusion, Risk Management and the Uncertain Rise of Index-Based Insurance. In: Pisking Capitalism, Research in Political Economy.[Soederberg, S. (ed.)]. Emerald Publishing Ltd, Bingley, UK, pp. 237-266. https://doi.org/10.1108/S0161-723020160000031013.
Taylor, M., 2017: Climate-smart agriculture: what is it good for?J Peasant Stud, 45 (1), 89–107, doi:10.1080/03066150.2017.1312355.
Taylor, M. and S. Bhasme, 2020: Between deficit rains and surplus populations: The political ecology of a climate-resilient village in South India. Geoforum, doi:10.1016/j.geoforum.2020.01.007.
Taylor, R.L., et al., 2018: Demography of the Pacific walrus (Odobenus rosmarus divergens) in a changing Arctic. Mar. Mamm. Sci. , 34 (1), 54–86, doi:10.1111/mms.12434.
Teigiserova, D.A., L. Hamelin and M. Thomsen, 2020: Towards transparent valorization of food surplus, waste and loss: Clarifying definitions, food waste hierarchy, and role in the circular economy. Sci. Total Environ. , 706, 136033, doi:10.1016/j.scitotenv.2019.136033.
Tejada, L. and S. Rist, 2018: Seeing land deals through the lens of the ‘land–water nexus’: the case of biofuel production in Piura, Peru. J Peasant Stud, 45 (7), 1247–1271, doi:10.1080/03066150.2016.1259220.
Tesfaye, A., et al., 2019: Estimating the economic value of climate services for strengthening resilience of smallholder farmers to climate risks in Ethiopia: A choice experiment approach. Ecol. Econ. , 162, 157–168, doi:10.1016/j.ecolecon.2019.04.019.
Teslić, N., et al., 2019: Future climatic suitability of the Emilia-Romagna (Italy) region for grape production. Reg Environ Change, 19 (2), 599–614, doi:10.1007/s10113-018-1431-6.
Teslić, N., et al., 2017: Climatic shifts in high quality wine production areas, Emilia Romagna, Italy, 1961-2015. Clim. Res. , 73 (3), 195–206, doi:10.3354/cr01468.
Tester, P.A., R.W. Litaker and E. Berdalet, 2020: Climate change and harmful benthic microalgae. Harmful Algae, 91, 101655, doi:10.1016/j.hal.2019.101655.
Tewari, J.C., K. Pareek, K. Shiran and M.M. Roy, 2017: Non timber tree products: alternative source of livelihood in man-crop/grasslivestock-tree/shrub continuum of arid regions. MOJ Ecol. Environ. Sci. , 2, 1–7.
Thamo, T., et al., 2017: Climate change impacts and farm-level adaptation: Economic analysis of a mixed cropping-livestock system. Agric Syst , 150, 99–108, doi:10.1016/j.agsy.2016.10.013.
Thapa, L.B., H. Thapa and B.G. Magar, 2015: Perception, trends and impacts of climate change in Kailali District, Far West Nepal. Int. J. Environ. , 4, 62–76.
Theusme, C., et al., 2021: Climate change vulnerability of confined livestock systems predicted using bioclimatic indexes in an arid region of Mexico. Sci. Total Environ. , 751, 141779, doi:10.1016/j.scitotenv.2020.141779.
Thiault, L., et al., 2019: Escaping the perfect storm of simultaneous climate change impacts on agriculture and marine fisheries. Sci. Adv. , 5 (11), eaaw9976, doi:10.1126/sciadv.aaw9976.
Thilsted, S.H., et al., 2016: Sustaining healthy diets: The role of capture fisheries and aquaculture for improving nutrition in the post-2015 era. Food Policy, 61, 126–131, doi:10.1016/j.foodpol.2016.02.005.
Thom, D. and R. Seidl, 2016: Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. Camb. Philos. Soc. , 91 (3), 760–781, doi:10.1111/brv.12193.
Thomaier, S., et al., 2015: Farming in and on urban buildings: Present practice and specific novelties of Zero-Acreage Farming (ZFarming). Renew. Agric. Food Syst. , 30 (1), 43–54, doi:10.1017/S1742170514000143.
Thomas, E., C. Alcázar Caicedo, J. Loo and R. Kindt, 2014: The distribution of the Brazil nut (Bertholletia excelsa) through time: from range contraction in glacial refugia, over human-mediated expansion, to anthropogenic climate change. Bol Do Mus Para Emílio Goeldi Ciências Nat , 9, 267–291.
Thomas, K. A., 2020: Shifting baselines of disaster mitigation. Clim. Dev. , 12 (2), 147–150, doi:10.1080/17565529.2019.1605875.
Thomas, P. and U. Buntgen, 2019: A risk assessment of Europe’s black truffle sector under predicted climate change. Sci. Total Environ. , 655, 27–34, doi:10.1016/j.scitotenv.2018.11.252.
Thomas, Y., C. Cassou, P. Gernez and S. Pouvreau, 2018: Oysters as sentinels of climate variability and climate change in coastal ecosystems. Environ. Res. Lett. , 13 (10), doi:10.1088/1748-9326/aae254.
Thompson-Hall, M., E.R. Carr and U. Pascual, 2016: Enhancing and expanding intersectional research for climate change adaptation in agrarian settings. Ambio, 45 (S3), 373–382, doi:10.1007/s13280-016-0827-0.
Thompson, W., et al., 2018: Automatic responses of crop stocks and policies buffer climate change effects on crop markets and price volatility. Ecol. Econ. , 152, 98–105, doi:10.1016/j.ecolecon.2018.04.015.
Thomsen, J., et al., 2017: Naturally acidified habitat selects for ocean acidification-tolerant mussels. Sci. Adv. , 3 (4), e1602411, doi:10.1126/sciadv.1602411.
Thornton, P., et al., 2018: Agriculture in a changing climate: Keeping our cool in the face of the hothouse. Outlook Agric, 47 (4), 283–290, doi:10.1177/0030727018815332.
Thornton, P., et al., 2018: Is agricultural adaptation to global change in lower—income countries on track to meet the future food production challenge?Glob. Environ. Chang. , 52, 37–48.
Thornton, P.K. and M. Herrero, 2014: Climate change adaptation in mixed crop -livestock systems in developing countries. Glob. Food Secur. Policy Econ. Environ. , 3 (2), 99–107, doi:10.1016/j.gfs.2014.02.002.
Thornton, P.K. and M. Herrero, 2015: Adapting to climate change in the mixed crop and livestock farming systems in sub-Saharan Africa. Nat. Clim. Chang. , 5 (9), 830–836, doi:10.1038/Nclimate2754.
Thornton, P.K., M. Herrero and R.B. Boone, 2019: Altered grazing systems: Pastoralism to conventional agriculture. Cambridge University Press, Cambridge, UK, 257–275.
Thornton, P.K., G.C. Nelson, D. Mayberry and M. Herrero, 2021: Increases in extreme heat stress in domesticated livestock species during the twenty-first century. Glob. Change Biol. , doi:10.1111/gcb.15825.
Thornton, T.F. and C. Comberti, 2017: Synergies and trade-offs between adaptation, mitigation and development. Clim Change, 140 (1), 5–18, doi:10.1007/s10584-013-0884-3.
Tian, X., et al., 2016: Global climate change impacts on forests and markets. Environ. Res. Lett. , 11 (035011).
Tibesigwa, B., M. Visser and J. Turpie, 2017: Climate change and South Africa’s commercial farms: an assessment of impacts on specialised horticulture, crop, livestock and mixed farming systems. Environ. Dev. Sustain. , 19 (2), 607–636, doi:10.1007/s10668-015-9755-6.
Tiemann, L.K., et al., 2015: Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecol Lett , 18 (8), 761–771, doi:10.1111/ele.12453.
Tigchelaar, M., D.S. Battisti and J.T. Spector, 2020: Work adaptations insufficient to address growing heat risk for U.S. agricultural workers. Environ. Res. Lett. , 15 (9), 94035–94035, doi:10.1088/1748-9326/ab86 f4.
Tilahun, M., et al., 2015: Rural households’ demand for frankincense forest conservation in Tigray, Ethiopia: a contingent valuation analysis. Land Degrad. Dev. , 26 (7), 642–653, doi:10.1002/ldr.2207.
Tilbrook, B., et al., 2019: An enhanced ocean acidification observing network: from people to technology to data synthesis and information exchange. Front. Mar. Sci. , 6 (JUN), doi:10.3389/fmars.2019.00337.
Tiller, R. and R. Richards, 2018: Ocean futures: exploring stakeholders’ perceptions of adaptive capacity to changing marine environments in northern Norway. Mar. Policy. , 95, 227–238, doi:10.1016/j.marpol.2018.04.001.
Tillotson, M.D., et al., 2019: Artificial selection on reproductive timing in hatchery salmon drives a phenological shift and potential maladaptation to climate change. Evol Appl, 12 (7), 1344–1359, doi:10.1111/eva.12730.
Tirado, C., 2015: Addressing the challenges of climate change to food security, safety, and nutrition. In: Global Climate Change and Human Health: From Science to Practice[Sons, J.W. (ed.)], Jossey-Bass, San Francisco, California, pp. 267-309
Titz, A. and S.S. Chiotha, 2019: Pathways for sustainable and inclusive cities in southern and eastern africa through urban green infrastructure?Sustainability, 11 (10), 2729–2729, doi:10.3390/su11102729.
To, H. and R.Q. Grafton, 2015: Oil prices, biofuels production and food security: past trends and future challenges. Food Secur, 7 (2), 323–336, doi:10.1007/s12571-015-0438-9.
Tom-Dery, D., F. Eller, C. Reisdorff and K. Jensen, 2018: Shea (Vitellaria paradoxa CF Gaertn.) at the crossroads: current knowledge and research gaps. Agrofor. Syst. , 92 (5), 1353–1371.
Toniello, G., et al., 2019: 11,500 y of human-clam relationships provide long-term context for intertidal management in the Salish Sea, British Columbia. Proc. Natl. Acad. Sci. USA, 116 (44), 22106–22114, doi:10.1073/pnas.1905921116.
Toomey, L., P. Fontaine and T. Lecocq, 2020: Unlocking the intraspecific aquaculture potential from the wild biodiversity to facilitate aquaculture development. Rev. Aquac. , 12 (4), 2212–2227, doi:10.1111/raq.12430.
Toreti, A., et al., 2020: Narrowing uncertainties in the effects of elevated CO2 on crops. Nat. Food, 1 (12), 775–782, doi:10.1038/s43016-020-00195-4.
Torres-Vitolas, C.A., et al., 2019: The socio-ecological dynamics of food insecurity among subsistence-oriented indigenous communities in Amazonia: a qualitative examination of coping strategies among riverine communities along the Caqueta river, Colombia. Hum Ecol, 47 (3), 355–368, doi:10.1007/s10745-019-0074-7.
Torres, J.M. and J.A. Casey, 2017: The centrality of social ties to climate migration and mental health. BMC Public Health, 17 (1), 600, doi:10.1186/s12889-017-4508-0.
Toth, G.G., et al., 2017: Malawi’s energy needs and agroforestry: adoption potential of woodlots. Hum Ecol, 45 (6), 735–746, doi:10.1007/s10745-017-9944-z.
Totin, E., et al., 2018: Institutional perspectives of climate-smart agriculture: a systematic literature review. Sustainability, 10 (6), 1990, doi:10.3390/su10061990.
Toufique, K. A. and B. Belton, 2014: Is aquaculture pro-poor? Empirical evidence of impacts on fish consumption in Bangladesh. World Dev, 64, 609–620, doi:10.1016/j.worlddev.2014.06.035.
Toure, A., B. Diekkruger, A. Mariko and A.S. Cisse, 2017: Assessment of groundwater resources in the context of climate change and population growth: case of the Klela basin in southern Mali. Climate, 5 (3), 45–45, doi:10.3390/cli5030045.
Toussaint, P. and A. Martínez Blanco, 2020: A human rights-based approach to loss and damage under the climate change regime. Clim. Policy, 20 (6), 743–757, doi:10.1080/14693062.2019.1630354.
Trainer, V.L., et al., 2020: Pelagic harmful algal blooms and climate change: Lessons from nature’s experiments with extremes. Harmful Algae, 91, 101591, doi:10.1016/j.hal.2019.03.009.
Tramel, S., 2018: Convergence as political strategy: social justice movements, natural resources and climate change. Third World Q, 39 (7), 1290–1307, doi:10.1080/01436597.2018.1460196.
Tran, L. and K. Brown, 2019: The importance of ecosystem services to smallholder farmers in climate change adaptation: learning from an ecosystem-based adaptation pilot in Vietnam. Agrofor. Syst. , 93 (5), 1949–1960, doi:10.1007/s10457-018-0302-y.
Tran, N., et al., 2020: Profitability and perceived resilience benefits of integrated shrimp-tilapia-seaweed aquaculture in North Central Coast, Vietnam. Mar. Policy, 120, 104153, doi:10.1016/j.marpol.2020.104153.
Traore, B., et al., 2013: Effects of climate variability and climate change on crop production in southern Mali. Eur J Agron, 49, 115–125, doi:10.1016/j.eja.2013.04.004.
Traore, B., et al., 2015: Climate variability and change in southern Mali: learning from farmer perceptions and on-farm trials. Exp. Agric. , 51 (4), 615–634, doi:10.1017/S0014479714000507.
Triguero-Mas, M., et al., 2015: Natural outdoor environments and mental and physical health: relationships and mechanisms. Environ Int , 77, 35–41, doi:10.1016/j.envint.2015.01.012.
Trisos, C.H., C. Merow and A.L. Pigot, 2020: The projected timing of abrupt ecological disruption from climate change. Nature, 580 (7804), 496–501, doi:10.1038/s41586-020-2189-9.
Troell, M., et al., 2017: Seafood from a changing Arctic. Ambio, 46 (Suppl 3), 368–386, doi:10.1007/s13280-017-0954-2.
Troell, M., et al., 2014: Does aquaculture add resilience to the global food system?Proc. Natl. Acad. Sci. USA, 111 (37), 13257–13263, doi:10.1073/pnas.1404067111.
Trouche, G., K. Vom Brocke, L. Temple and M. Guillet, 2016: Analyse de l’impact des programmes de sélection participative du sorgo conduits au Burkina Faso de 1995 à 2015. Montpellier, France, http://agritrop.cirad.fr/5809. (205 pp). (accessed August 31, 2021).
Tschakert, P., et al., 2017: Climate change and loss, as if people mattered: values, places, and experiences. Wiley. Interdiscip. Rev. Clim. Change. , 8 (5), 1–19, doi:10.1002/wcc.476.
Tschakert, P., et al., 2016: Micropolitics in collective learning spaces for adaptive decision making. Glob. Environ. Chang. , 40, 182–194, doi:10.1016/j.gloenvcha.2016.07.004.
Tseng, Y.-H., R. Ding and X. Huang, 2017: The warm Blob in the northeast Pacific—the bridge leading to the 2015/16 El Niño. Environ. Res. Lett. , 12 (5), 54019, doi:10.1088/1748-9326/aa67c3.
Tsikata, D. and J.A. Yaro, 2014: When a good business model is not enough: land transactions and gendered livelihood prospects in rural Ghana. Fem Econ, 20 (1), 202–226, doi:10.1080/13545701.2013.866261.
Turhan, E., C. Zografos and G. Kallis, 2015: Adaptation as biopolitics: why state policies in Turkey do not reduce the vulnerability of seasonal agricultural workers to climate change. Glob. Environ. Chang. , 31, 296–306, doi:10.1016/j.gloenvcha.2015.02.003.
Turner, C., et al., 2018: Concepts and critical perspectives for food environment research: a global framework with implications for action in low- and middle-income countries. Glob. Food Secur. Policy Econ. Environ. , 18, 93–101, doi:10.1016/j.gfs.2018.08.003.
Turner, M.G., K.H. Braziunas, W.D. Hansen and B.J. Harvey, 2019: Short-interval severe fire erodes the resilience of subalpine lodgepole pine forests. Proc. Natl. Acad. Sci. USA, 116 (23), 11319–11328, doi:10.1073/pnas.1902841116.
Turner, M.G., et al., 2020: Climate change, ecosystems and abrupt change: science priorities. Philos. Trans. R. Soc. Lond. B Biol. Sci. , 375 (1794), 20190105, doi:10.1098/rstb.2019.0105.
Turner, N.J. and H. Clifton, 2009: “it’s so different today”: climate change and indigenous lifeways in British Columbia, Canada. Glob. Environ. Change. , 19 (2), 180–190, doi:10.1016/j.gloenvcha.2009.01.005.
Turnhout, E., et al., 2017: Envisioning REDD plus in a post-Paris era: between evolving expectations and current practice. Wiley. Interdiscip. Rev. Clim. Change. , 8 (1), e425, doi:10.1002/wcc.425.
Turnock, S.T., et al., 2018: The impact of future emission policies on tropospheric ozone using a parameterised approach. Atmos. Chem. Phys. , 18 (12), 8953–8978, doi:10.5194/acp-18-8953-2018.
Tyler, N.J.C., 2010: Climate, snow, ice, crashes, and declines in populations of reindeer and caribou (Rangifer tarandus L.). Ecol Monogr, 80 (2), 197–219, doi:10.1890/09-1070.1.
Ubilava, D., 2018: The role of El Nino southern oscillation in commodity price movement and predictability. Am J Agric Econ, 100 (1), 239–263, doi:10.1093/ajae/aax060.
Ubilava, D. and M. Abdolrahimi, 2019: The El Nino impact on maize yields is amplified in lower income teleconnected countries. Environ. Res. Lett. , 14 (5), doi:10.1088/1748-9326/ab0cd0.
Uchiyama, T., F.J. Mueter and G.H. Kruse, 2020: Multispecies biomass dynamics models reveal effects of ocean temperature on predation of juvenile pollock in the eastern Bering Sea. Fish Oceanogr, 29 (1), 10–22, doi:10.1111/fog.12433.
Ujiie, K., et al., 2019: How elevated CO2 affects our nutrition in rice, and how we can deal with it. PLoS ONE, 14 (3), e212840, doi:10.1371/journal.pone.0212840.
UK, T. G. F. S. P., 2015: Extreme Weather and Resilience of the lobal Food System. Final Project Report . UK-US Taskforce on Extreme Weather and Global Food System Resilience, London, UK.
ul Haq, S., I. Boz and P. Shahbaz, 2021: Adoption of climate-smart agriculture practices and differentiated nutritional outcome among rural households: a case of Punjab province, Pakistan. Food Secur, doi:10.1007/s12571-021-01161-z.
Ulrichs, M., R. Slater and C. Costella, 2019: Building resilience to climate risks through social protection: from individualised models to systemic transformation. Disasters, 43 (S3), 368–S387, doi:10.1111/disa.12339.
UN, 2018: The World’s Cities in 2018 - Data Booklet (ST/ESA/SER.A/417). United Nations, Available at: https://digitallibrary.un.org/record/3799524 (accessed August 31, 2021).
UNDRR, 2019: Global Assessment Report on Disaster Risk Reduction 2019. United Nations, 472-472 pp. Available at: https://www.un-ilibrary.org/content/books/9789210041805 (accessed 31/8/2021)..
UNDRR (United Nations Office for Disaster Risk Reduction - Regional Office for Asia and Pacific), 2020: COVID-19 Brief: Combating the dual challenges of climate-related disasters and COVID-19. https://www.undrr.org/publication/undrr-asia-pacific-covid-19-brief-combating-dual-challenges-climate-related-disasters.(accessed August 31, 2021).
UNEP, 2016: The Adaptation Finance Gap Report . UNEP, Programme, U. N. E., Nairobi, Kenya, https://climateanalytics.org/media/agr2016.pdf.(accessed August 31, 2021)
UNEP, 2019: Emissions Gap Report 2019. Nairobi, https://www.unenvironment.org/resources/emissions-gap-report-2019.(accessed August 31, 2021).
UNEP, 2021: Adaptation Gap Report 2020. Nairobi, https://www.unep.org/resources/adaptation-gap-report-2020.(accessed August 31, 2021).
UNFCCC, 2013: Best Practices and Available Tools for the Use of Indigenous and Traditional Knowledge and Practices for Adaptation, and the Application of Gender-Sensitive Approaches and Tools for Understanding and Assessing Impacts, Vulnerability and Adaptation to Climate Change. United Nations, https://unfccc.int/documents/7927. Accessed 2021 . (62 pp).
UNFCCC, 2018: Summary and recommendations by the Standing Committee on Finance on the 2018 Biennial Assessment and Overview of Climate Finance Flows. https://unfccc.int/sites/default/files/resource/51904. (UNFCCC BA 2018 - Summary Final.pdf.).(accessed August 31, 2021).
UNHRC, 2017: Human Rights and Climate Change. A/HRC/35/L.32, The Office of the High Commissioner for Human Rights (OHCHR), Geneva, Switzerland. Available at: https://documents-dds-ny.un.org/doc/UNDOC/LTD/G17/167/92/PDF/G1716792.pdf?OpenElement (accessed January 6, 2022).
Upgupta, S., et al., 2015: Climate change impact and vulnerability assessment of forests in the Indian Western Himalayan region: A case study of Himachal Pradesh, India. Clim. Risk Manag. , 10, 63–76, doi:10.1016/j.crm.2015.08.002.
Uson, M.A.M., 2017: Natural disasters and land grabs: the politics of their intersection in the Philippines following super typhoon Haiyan. Can. J. Dev. Stud. , 38 (3), 414–430, doi:10.1080/02255189.2017.1308316.
Uyttendaele, M., C. Liu and N. Hofstra, 2015: Special issue on the impacts of climate change on food safety. Food. Res. Int. , 68, 1–6, doi:10.1016/j.foodres.2014.09.001.
Vaara, M., O. Saastamoinen and M. Turtiainen, 2013: Changes in wild berry picking in Finland between 1997 and 2011. Scand. J. For. Res. , 28 (6), 586–595, doi:10.1080/02827581.2013.786123.
Vaillancourt, J.P. and N.H. Ogden, 2016: Emergence of infectious diseases associated with climate change: rural and urban challenges. Environnement Risques Sante, 15 (4), 312–316, doi:10.1684/ers.2016.0889.
Valdivia, R.O., et al., 2015: Representative agricultural pathways and scenarios for regional integrated assessment of climate change impacts, vulnerability, and adaptation. Handb. Clim. Chang. Agroecosystems, 3, 101–156.
Valencia, V., H. Wittman and J. Blesh, 2019: Structuring markets for resilient farming systems. Agron. Sustain. Dev. , 39 (2), 25–25, doi:10.1007/s13593-019-0572-4.
Valverde, P., et al., 2015: Climate change impacts on irrigated agriculture in the Guadiana river basin (Portugal). Agric. Water Manag. , 152, 17–30, doi:10.1016/j.agwat.2014.12.012.
van Berkel, D.B. and P.H. Verburg, 2014: Spatial quantification and valuation of cultural ecosystem services in an agricultural landscape. Ecol Indic, 37, 163–174, doi:10.1016/j.ecolind.2012.06.025.
Van der Fels-Klerx, H.J., L.C. Vermeulen, A.K. Gavai and C. Liu, 2019: Climate change impacts on aflatoxin B1 in maize and aflatoxin M1 in milk: A case study of maize grown in Eastern Europe and imported to the Netherlands. PLoS ONE, 14 (6), e218956, doi:10.1371/journal.pone.0218956.
van der Gaast, K., E. van Leeuwen and S. Wertheim-Heck, 2020: City-region food systems and second tier cities: from garden cities to garden regions. Sustainability, 12 (6), doi:10.3390/su12062532.
van der Werf, H.M.G., M.T. Knudsen and C. Cederberg, 2020: Towards better representation of organic agriculture in life cycle assessment. Nat. Sustain. , 3 (6), 419–425, doi:10.1038/s41893-020-0489-6.
van Dijk, M., T. Morley, M.L. Rau and Y. Saghai, 2021: A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food, 2 (7), 494–501, doi:10.1038/s43016-021-00322-9.
Van Dingenen, R. et al., 2018: Global trends of methane emissions and their impacts on ozone concentrations. JRC Science for Policy Report. Office for Official Publications of the European Union, Luxembourg.
van Dorst, R.M., et al., 2018: Warmer and browner waters decrease fish biomass production. Glob Chang Biol, 25 (4), 1395–1408, doi:10.1111/gcb.14551.
van Gameren, V. and E. Zaccai, 2015: Private forest owners facing climate change in Wallonia: Adaptive capacity and practices. Environ Sci Policy, 52, 51–60, doi:10.1016/j.envsci.2015.05.004.
van Keulen, H. and H. Schiere (eds.), New Directions for a Diverse Planet . 4th Int. Crop Science Congress, Brisbane, Australia. The Regional Institute Ltd, Gosford, Australia. ISBN 9781920842215.
van Leeuwen, C. and P. Darriet, 2016: The impact of climate change on viticulture and wine quality. J. Wine Econ. , 11, 150–167.
van Leeuwen, C. and A. Destrac-Irvine, 2017: Modified grape composition under climate change conditions requires adaptations in the vineyard. Oeno One, 51, 147.
van Noordwijk, M., et al., 2018: SDG synergy between agriculture and forestry in the food, energy, water and income nexus: reinventing agroforestry?Curr. Opin. Environ. Sustain. , 34, 33–42, doi:10.1016/j.cosust.2018.09.003.
Van Wijk, M., et al., 2014: Farm household models to analyse food security in a changing climate: A review. Glob. Food Secur. , 3 (2), 77–84.
van Zeist, W.-J., et al., 2020: Are scenario projections overly optimistic about future yield progress?Glob. Environ. Chang. , 64, doi:10.1016/j.gloenvcha.2020.102120.
van Zonneveld, M., M.-S. Turmel and J. Hellin, 2020: Decision-Making to Diversify Farm Systems for Climate Change Adaptation. Front. Sustain. Food Syst. , 4, doi:10.3389/fsufs.2020.00032.
Vanclay, F., 2017: Principles to gain a social licence to operate for green initiatives and biodiversity projects. Curr. Opin. Environ. Sustain. , 29, 48–56, doi:10.1016/j.cosust.2017.11.003.
Vandergeten, E., et al., 2016: Agricultural outsourcing or land grabbing: a meta-analysis. Landsc Ecol, 31 (7), 1395–1417, doi:10.1007/s10980-016-0365-y.
Vannuccini, S., et al., 2018: Chapter 3: Understanding the impacts of climate change for fisheries and aquaculture: global and regional supply and demand trends and prospects. In: Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options[Kalikoski, D.C., et al.(ed.)]. FAO, Rome, pp. 41–61.
Vanos, J., D.J. Vecellio and T. Kjellstrom, 2019: Workplace heat exposure, health protection, and economic impacts: A case study in Canada. Am. J. Ind. Med. , 62 (12), 1024–1037, doi:10.1002/ajim.22966.
Varshney, R.K., C. Ojiewo and E. Monyo, 2019: A decade of Tropical Legumes projects: Development and adoption of improved varieties, creation of market-demand to benefit smallholder farmers and empowerment of national programmes in sub-Saharan Africa and South Asia. Plant Breed, 138 (4), 379–388, doi:10.1111/pbr.12744.
Varshney, R.K., et al., 2018: Can genomics deliver climate-change ready crops?Curr. Opin. Plant Biol. , 45 (Pt B), 205–211, doi:10.1016/j.pbi.2018.03.007.
Vasconcelos, V.V., P.M. Hannam, S.A. Levin and J.M. Pacheco, 2020: Coalition-structured governance improves cooperation to provide public goods. Sci Rep, 10 (1), 9194, doi:10.1038/s41598-020-65960-8.
Vaughan, C., S. Dessai and C. Hewitt, 2018: Surveying climate services: What can we learn from a bird’s-eye view?Weather. Clim. Soc. , 10, 373–395.
Vaughan, C., et al., 2019: Evaluating agricultural weather and climate services in Africa: Evidence, methods, and a learning agenda. Wiley. Interdiscip. Rev. Clim. Change. , 10 (4), doi:10.1002/wcc.586.
Veldman, J.W., et al., 2015: Where tree planting and forest expansion are bad for Biodiversity and ecosystem services. BioScience, 65 (10), 1011–1018, doi:10.1093/biosci/biv118.
Veldwisch, G.J., J. Franco and L. Mehta, 2018: Water grabbing: Practices of contestation and appropriation of water resources in the context of expanding global capital. Water Justice, 59–70, doi:10.1017/9781316831847.004.
Velenturf, A.P.M., et al., 2019: Circular economy and the matter of integrated resources. Sci. Total Environ. , 689, 963–969, doi:10.1016/j.scitotenv.2019.06.449.
Venkatesh Mannar, M.G., R. Micha and I.E. Group, 2020: 2020 Global Nutrition Report: Action on equity to end malnutrition. Development Initiatives, Bristol, UK.
Verdolini, E., C. Bak, J. Ruet and A. Venkatachalam, 2018: Innovative green-technology SMEs as an opportunity to promote financial de-risking. Econ. Open Access Open-assessment E-journal, 12, doi:10.5018/economics-ejournal.ja.2018-14.
Verheecke-Vaessen, C., et al., 2019: Interacting climate change environmental factors effects on Fusarium langsethiae growth, expression of Tri genes and T-2/HT-2 mycotoxin production on oat-based media and in stored oats. Fungal Biol, 123 (8), 618–624, doi:10.1016/j.funbio.2019.04.008.
Verkerk, P.J., et al., 2020: Climate-Smart Forestry: the missing link. Elsevier B.V., ISBN 102164102164.
Verma, M., T. Hertel and N. Diffenbaugh, 2014: Market-oriented ethanol and corn-trade policies can reduce climate-induced US corn price volatility. Environ. Res. Lett. , 9 (6), 64028, doi:10.1088/1748-9326/9/6/064028.
Vermeulen, S.J., et al., 2018: Transformation in practice: a review of empirical cases of transformational adaptation in agriculture under climate change. Front. Sustain. Food Syst. , 2, doi:10.3389/fsufs.2018.00065.
Verschuur, J., S. Li, P. Wolski and F.E.L. Otto, 2021: Climate change as a driver of food insecurity in the 2007 Lesotho-South Africa drought. Sci Rep, 11 (1), 3852–3852, doi:10.1038/s41598-021-83375-x.
Vianna, G.M.S., D. Zeller and D. Pauly, 2020: Fisheries and policy implications for human nutrition. Curr Environ Health Rep, 7 (3), 161–169, doi:10.1007/s40572-020-00286-1.
Vidal-González, P. and B. Nahhass, 2018: The use of mobile phones as a survival strategy amongst nomadic populations in the Oriental region (Morocco). GeoJournal, 83, 1079–1090, doi:10.1007/s10708-017-9823-6.
Vijge, M.J., M. Brockhaus, M. Di Gregorio and E. Muharrom, 2016: Framing national REDD plus benefits, monitoring, governance and finance: A comparative analysis of seven countries. Glob. Environ. Change. , 39, 57–68, doi:10.1016/j.gloenvcha.2016.04.002.
Villasante, S., et al., 2015: Are provisioning ecosystem services from rural aquaculture contributing to reduce hunger in Africa?Ecosyst. Serv. , 16, 365–377, doi:10.1016/j.ecoser.2015.07.003.
Vinyeta, K., F.K. Lake and K. Norgaard, 2016: Vulnerabilities of Traditional Foods and Cultural Use Species. Karuk Tribe Climate Vulnerability Assessment- Assessing vulnerabilities from the increased frequency of high severity fire, Karuk Tribe Department of Natural Resources, USA. Available at: https://karuktribeclimatechangeprojects.com/chapter-3-vulnerabilities-of-traditional-foods-and-cultural-use-species/ (accessed August 31, 2021).
Vira, B., C. Wildburger and S. Mansourian, 2016: Forests and Food: Addressing Hunger and Nutrition Across Sustainable Landscapes. Open Book Publishers, UK and USA, ISBN 978-1783741939.
Visch, W., et al., 2019: Underpinning the development of seaweed biotechnology: cryopreservation of brown algae (Saccharina latissima) Gametophytes. Biopreserv Biobank, 17 (5), 378–386, doi:10.1089/bio.2018.0147.
Vitasse, Y., et al., 2018: Increase in the risk of exposure of forest and fruit trees to spring frosts at higher elevations in Switzerland over the last four decades. Agric. For. Meteorol. , 248, 60–69, doi:10.1016/j.agrformet.2017.09.005.
Vitt, R., et al., 2017: Modelled performance of energy saving air treatment devices to mitigate heat stress for confined livestock buildings in Central Europe. Biosys. Eng. , 164, 85–97, doi:10.1016/j.biosystemseng.2017.09.013.
Vogel, E., et al., 2019: The effects of climate extremes on global agricultural yields. Environ. Res. Lett. , 14 (5), doi:10.1088/1748-9326/ab154b.
Voggesser, G., et al., 2013: Cultural impacts to tribes from climate change influences on forests. In: Climate change and indigenous peoples in the United States. [Maldonado, J. K., B. Colombi and R. Pandya (eds.)]. Springer, Cham, Switzerland, pp. 107-118.
Vogliano, C., et al., 2021: Progress towards SDG 2: Zero hunger in melanesia – A state of data scoping review. Glob. Food Secur. , 29, 100519, doi:10.1016/j.gfs.2021.100519.
Von Lampe, M., et al., 2014: Why do global long-term scenarios for agriculture differ? An overview of the AgMIP global economic model intercomparison. Agric. Econ. , 45 (1), 3–20.
Voss-Fels, K. and R.J. Snowdon, 2016: Understanding and utilizing crop genome diversity via high-resolution genotyping. Plant Biotechnol J, 14, 1086–1094, doi:10.1111/pbi.12456.
Voss, R.C., 2021: On- and non-farm adaptation in Senegal: understanding differentiation and drivers of farmer strategies. Clim. Dev. , 1–15, doi:10.1080/17565529.2021.1881424.
Waha, K., et al., 2013: Adaptation to climate change through the choice of cropping system and sowing date in sub-Saharan Africa. Glob. Environ. Change. , 23 (1), 130–143, doi:10.1016/j.gloenvcha.2012.11.001.
Waha, K., et al., 2018: Agricultural diversification as an important strategy for achieving food security in Africa. Glob Chang Biol, 24 (8), 3390–3400, doi:10.1111/gcb.14158.
Wako, G., M. Tadesse and A. Angassa, 2017: Camel management as an adaptive strategy to climate change by pastoralists in southern Ethiopia. Ecol Process, 6, 26, doi:10.1186/s13717-017-0093-5.
Waldbusser, G.G., et al., 2015: Ocean acidification has multiple modes of action on bivalve larvae. PLoS ONE, 10 (6), e128376, doi:10.1371/journal.pone.0128376.
Waldron, A., et al., 2017: Agroforestry can enhance food security while meeting other sustainable development goals. Trop. Conserv. Sci, 10, 194008291772066–194008291772066, doi:10.1177/1940082917720667.
Wallbott, L., 2014: Indigenous peoples in UN REDD plus negotiations: “importing power” and lobbying for rights through discursive interplay management. Ecol. Soc. , 19 (1), doi:10.5751/Es-06111-190121.
Walsh, M.G., A.W. de Smalen and S.M. Mor, 2018: Climatic influence on anthrax suitability in warming northern latitudes. Sci Rep, 8 (1), 9269, doi:10.1038/s41598-018-27604-w.
Wang, C.J., Z.X. Zhang and J.Z. Wan, 2019a: Vulnerability of global forest ecoregions to future climate change. Glob. Ecol. Conserv. , 20, e760, doi:10.1016/j.gecco.2019.e00760.
Wang, G.J., et al., 2020a: A unique feature of the 2019 extreme positive Indian ocean dipole event. Geophys. Res. Lett. , 47 (18), doi:10.1029/2020GL088615.
Wang, J., et al., 2019b: Changes in grain protein and amino acids composition of wheat and rice under short-term increased [CO2 ] and temperature of canopy air in a paddy from East China. New Phytol. , 222 (2), 726–734, doi:10.1111/nph.15661.
Wang, J., et al., 2020b: Global warming effects on climate zones for wine grape in Ningxia region, China. Theor Appl Climatol, 140 (3-4), 1527–1536, doi:10.1007/s00704-020-03170-y.
Wang, W.L., et al., 2018: Elevated CO2 cannot compensate for japonica grain yield losses under increasing air temperature because of the decrease in spikelet density. Eur J Agron, 99, 21–29, doi:10.1016/j.eja.2018.06.005.
Wang, X., H. Li and I. García de Cortázar Atauri, 2020c: Assessing grapevine phenological models under Chinese climatic conditions. Oeno One, 54 (3), doi:10.20870/oeno-one.2020.54.3.3195Warnatzsch, E.A. and D.S. Reay, 2020: Climate Change Impact on Aflatoxin Contamination Risk in Malawi’s Maize Crops. Front. Sustain. Food Syst , 4, doi:10.3389/fsufs.2020.591792.
Warren, R., et al., 2018: The implications of the United Nations Paris Agreement on climate change for globally significant biodiversity areas. Clim Change, 147 (3), 395–409, doi:10.1007/s10584-018-2158-6.
Wartenberg, A.C., et al., 2020: Soil fertility and Theobroma cacao growth and productivity under commonly intercropped shade-tree species in Sulawesi, Indonesia. Plant Soil, 453 (1), 87–104.
Waryszak, P., T.I. Lenz, M.R. Leishman and P.O. Downey, 2018: Herbicide effectiveness in controlling invasive plants under elevated CO2: Sufficient evidence to rethink weeds management. J Environ Manage, 226, 400–407, doi:10.1016/j.jenvman.2018.08.050.
Watkiss, P., 2015: A review of the economics of adaptation and climate resilient development . Working Paper, No. 205, Grantham Research Institute on Climate Change and the Environment London, UK. Available at: http://www.vfmadaptation.com/Working-Paper-205-Watkiss.pdf.
Watson, C., et al., 2015: Finance for reducing disaster risk: 10 things to know. Oversea Development Institute, London, . Available at: https://www.undp.org/content/dam/undp/library/crisis%20prevention/disaster/Finance%20for%20reducing%20disaster%20risk-10-Things-to-know-report.pdf (accessed August 31, 2021).
Watson, E.E., H.H. Kochore and B.H. Dabasso, 2016a: Camels and Climate Resilience: Adaptation in Northern Kenya. Hum Ecol, 44 (6), 701–713, doi:10.1007/s10745-016-9858-1.
Watson, J.R., F. Armerin, D.H. Klinger and B. Belton, 2018: Resilience through risk management: cooperative insurance in small-holder aquaculture systems. Heliyon, 4 (9), e799, doi:10.1016/j.heliyon.2018.e00799.
Watson, S., et al., 2016b: Dietary exposure to aflatoxin and micronutrient status among young children from Guinea. Mol. Nutr. Food Res. , 60 (3), 511–518, doi:10.1002/mnfr.201500382.
Watts, N., et al., 2021: The 2020 report of The Lancet Countdown on health and climate change: responding to converging crises. Lancet , 397 (10269), 129–170, doi:10.1016/S0140-6736(20)32290-X.
Watts, N., et al., 2018: The Lancet Countdown on health and climate change: from 25 years of inaction to a global transformation for public health. Lancet , 391 (10120), 581–630, doi:10.1016/S0140-6736(17)32464-9.
Weatherdon, L.V., et al., 2016: Observed and Projected Impacts of Climate Change on Marine Fisheries, Aquaculture, Coastal Tourism, and Human Health: An Update. Front. Mar. Sci. , 3 (APR), doi:10.3339/fmars.2076.00048.
Webber, H., et al., 2018: Diverging importance of drought stress for maize and winter wheat in Europe. Nat. Commun. , 9 (1), 4249, doi:10.1038/s41467-018-06525-2.
Webber, S., 2017: Circulating climate services: commercializing science for climate change adaptation in pacific islands. Geoforum, 85 (November 2016), 82–91, doi:10.1016/j.geoforum.2017.07.009.
Webster, C.R., M. A. Jenkins and S. Jose, 2006: Woody invaders and the challenges they pose to forest ecosystems in the eastern United States. J. For. , 104 (7), 366–374, doi:10.1093/jof/104.7.366.
Weerakoon, D.K. and S. Wijesundara (eds.), 2012: The National Red List 2012 of Sri Lanka; Conservation Status of the Fauna and Flora. Ministry of the Environment, Colombo, Sri Lanka, ISBN 978-9550033553. 452 pp.
WEF, 2017: The Global Risks Report 2017. World Economic Forum. 78 pp.
Wegner, K., et al., 2016: Effects of temperature and temperature-humidity index on the reproductive performance of sows during summer months under a temperate climate. Anim. Sci. J. , 87 (11), 1334–1339, doi:10.1111/asj.12569.
Weidner, T. and A.D. Yang, 2020: The potential of urban agriculture in combination with organic waste valorization: Assessment of resource flows and emissions for two european cities. J Clean Prod, 244, doi:10.1016/j.jclepro.2019.118490.
Wein, E.E., M.M.R. Freeman and J.C. Makus, 1996: Use of and preference for traditional foods among the Belcher Island Inuit. Arctic, 49 (3), 256–264.
Weindl, I., et al., 2017: Livestock production and the water challenge of future food supply: Implications of agricultural management and dietary choices. Glob. Environ. Change. , 47, 121–132, doi:10.1016/j.gloenvcha.2017.09.010.
Weindl, I., et al., 2015: Livestock in a changing climate: production system transitions as an adaptation strategy for agriculture. Environ. Res. Lett. , 10 (9), 94021, doi:10.1088/1748-9326/10/9/094021.
Weinreb, A., G. Stecklov and A. Arslan, 2020: Effects of changes in rainfall and temperature on age- and sex-specific patterns of rural-urban migration in sub-Saharan Africa. Popul Environ, 42 (2), 219–254, doi:10.1007/s11111-020-00359-1.
Weitz, N., C. Strambo, E. Kemp-Benedict and M. Nilsson, 2017: Closing the governance gaps in the water-energy-food nexus: Insights from integrative governance. Glob. Environ. Change. , 45, 165–173, doi:10.1016/j.gloenvcha.2017.06.006.
Wells, M.L., et al., 2020: Future HAB science: directions and challenges in a changing climate. Harmful Algae, 91, 101632, doi:10.1016/j.hal.2019.101632.
Weltzien, E., et al., 2019: Long-term collaboration between farmers’ organizations and plant breeding programmes: Sorghum and pearl millet in West Africa. In: Farmers and Plant Breeding[Westengen, O. T. and T. Winge (eds.)]. Routledge, London and New York, pp. 29-48Wenndt, A.J., H.K. Sudini, P. Pingali and R. Nelson, 2021: Farmer research networks enable community-based mycotoxin management in rural Indian villages. Agric Syst , 192, 103192, doi:10.1016/j.agsy.2021.103192.
Wesche, S.D. and H.M. Chan, 2010: Adapting to the impacts of climate change on food security among Inuit in the Western Canadian Arctic. EcoHealth, 7 (3), 361–373, doi:10.1007/s10393-010-0344-8.
West, T.A.P., 2016: Indigenous community benefits from a de-centralized approach to REDD plus in Brazil. Clim. Policy, 16 (7), 924–939, doi:10.1080/14693062.2015.1058238.
Westengen, O.T. and T. Berg, 2016: Crop adaptation to climate change in SSA: the role of genetic resources and seed systems. In: Climate Change and Multi-Dimensional Sustainability in African Agriculture. [Lal, R., D. Kraybill, D. O. Hansen, B. R. Singh, T. Mosogoya and L. O. Eik (eds.)]. Springer International Publishing, Cham, pp. 327-343.Westengen, O.T. and A.K. Brysting, 2014: Crop adaptation to climate change in the semi-arid zone in Tanzania: the role of genetic resources and seed systems. Agric. Food Secur. , 3 (1), 3–3, doi:10.1186/2048-7010-3-3.
Westengen, O.T., R. Haug, P. Guthiga and E. Macharia, 2019: Governing Seeds in East Africa in the Face of Climate Change: Assessing Political and Social Outcomes. Front. Sustain. Food Syst. , 3, doi:10.3389/fsufs.2019.00053.
Westengen, O.T., et al., 2018: A climate for commerce: the political agronomy of conservation agriculture in Zambia. Agric Hum Values, 35 (1), 255–268, doi:10.1007/s10460-017-9820-x.
Westmeijer, G., et al., 2019: Mechanistic niche modelling to identify favorable growth sites of temperate macroalgae. Algal Res. Biofuels Bioprod. , 41, doi:10.1016/j.algal.2019.101529.
Westoby, R., K.E. McNamara, R. Kumar and P.D. Nunn, 2020: From community-based to locally led adaptation: Evidence from Vanuatu. Ambio, 49 (9), 1466–1473, doi:10.1007/s13280-019-01294-8.
Weyant, C., et al., 2018: Anticipated burden and mitigation of carbon-dioxide-induced nutritional deficiencies and related diseases: A simulation modeling study. PLoS Med, 15 (7), e1002586, doi:10.1371/journal.pmed.1002586.
WFP-FSIN, 2020: Global Report on Food Crises: Joint Analysis for Better Decisions. United Nations World Food Programme, Food Security Information Network. FAO, WFP, IFPRI, Rome, Italy and Washington, DC.
White, D., 2014: A perfect storm? Indigenous rights within a national REDD+ readiness process in Peru. Mitig Adapt Strateg Glob Change, 19, 657–676, doi:10.1007/s11027-013-9523-6.
White, J.C., et al., 2017: A nationwide annual characterization of 25 years of forest disturbance and recovery for Canada using Landsat time series. Remote. Sens. Environ. , 194, 303–321, doi:10.1016/j.rse.2017.03.035.
White, J.T. and C. Bunn, 2017: Growing in Glasgow: Innovative practices and emerging policy pathways for urban agriculture. Land Use Policy, 68 (August), 334–344, doi:10.1016/j.landusepol.2017.07.056.
Whitfield, S., J.L. Dixon, B.P. Mulenga and H. Ngoma, 2015: Conceptualising farming systems for agricultural development research: Cases from Eastern and Southern Africa. Agric Syst , 133, 54–62, doi:10.1016/j.agsy.2014.09.005.
Whitmee, S., et al., 2015: Safeguarding human health in the Anthropocene epoch: report of The Rockefeller Foundation–Lancet Commission on planetary health. Lancet , 386 (10007), 1973–2028, doi:10.1016/s0140-6736(15)60901-1.
Whitney, C.K., T. Conger, N.C. Ban and R. McPhie, 2020: Synthesizing and communicating climate change impacts to inform coastal adaptation planning. FACETS, 5 (1), 704–737, doi:10.1139/facets-2019-0027.
WHO, 2013: WHO traditional medicine strategy: 2014-2023. World Health Organization, Geneva, Switzerland, ISBN 978-9241506090.
WHO, 2021: The Global Health Observatory. Available at: https://www.who.int/data/gho/data/. (accessed August 31, 2021)
Wichelns, D., 2017: The water-energy-food nexus: Is the increasing attention warranted, from either a research or policy perspective?Environ Sci Policy, 69, 113–123, doi:10.1016/j.envsci.2016.12.018.
Widianingsih, N.N., L.H. Schmidt and I. Theilade, 2019: Jernang (Daemonorops spp.) commercialization and its role for rural incomes and livelihoods in Southern Sumatra, Indonesia. For. Trees Livelihoods, 28 (3), 143–159.
Wiebe, K., et al., 2015: Climate change impacts on agriculture in 2050 under a range of plausible socioeconomic and emissions scenarios. Environ. Res. Lett. , 10 (8), 85010, doi:10.1088/1748-9326/10/8/085010.
Wiesmeier, M., et al., 2019: Soil organic carbon storage as a key function of soils - A review of drivers and indicators at various scales. Geoderma, 333, 149–162, doi:10.1016/j.geoderma.2018.07.026.
Wijngaard, R.R., et al., 2017: The impact of climate change on metal transport in a lowland catchment. Water Air Soil Pollut , 228 (3), 107, doi:10.1007/s11270-017-3261-4.
Wild, C.P., J.D. Miller and J.D. Groopman (eds.), 2015: Mycotoxin Control in Low- and Middle-Income Countries, IARC World Group Reports. International Agency for Research on Cancer, Lyon, France, ISBN 978-9283225102. 66 pp.
Wilkes, A., et al., 2017: Is cross-breeding with indigenous sheep breeds an option for climate-smart agriculture?Small. Rumin. Res. , 147, 83–88, doi:10.1016/j.smallrumres.2016.12.036.
Willett, W., et al., 2019: Food in the Anthropocene: the EAT-Lancet Commission on healthy diets from sustainable food systems. Lancet , 393 (10170), 447–492, doi:10.1016/S0140-6736(18)31788-4.
Williams, A., et al., 2016: Soil water holding capacity mitigates downside risk and volatility in US rainfed maize: time to invest in soil organic matter?PLoS ONE, 11 (8), e160974–e160974, doi:10.1371/journal.pone.0160974.
Williams, A., et al., 2018: A regionally-adapted implementation of conservation agriculture delivers rapid improvements to soil properties associated with crop yield stability. Sci Rep, 8 (1), 8467–8467, doi:10.1038/s41598-018-26896-2.
Williams, A., et al., 2015: Quantifying the response of cotton production in eastern Australia to climate change. Clim Change, 129 (1-2), 183–196, doi:10.1007/s10584-014-1305-y.
Williams, P.H. and J.L. Osborne, 2009: Bumblebee vulnerability and conservation world-wide. Apidologie, 40 (3), 367–387, doi:10.1051/apido/2009025.
Williamson, B., 2021: Cultural Burning in NSW: Challenges and Opportunities for Policy Makers and Aboriginal Peoples. CAEPR Working Paper. ACT: Australian National University, Centre for Aboriginal Economic Policy Research (CAEPR), Canberra.
Wilson, M.J., et al., 2015: Invasive slug pests and their parasites-temperature responses and potential implications of climate change. Biol Fertil Soils, 51 (6), 739–748, doi:10.1007/s00374-015-1022-3.
Winans, K., A. Kendall and H. Deng, 2017: The history and current applications of the circular economy concept. Renew. Sustain. Energy Rev. , 68, 825–833, doi:10.1016/j.rser.2016.09.123.
Winkler, J.A., et al., 2018: Potential impacts of climate change on storage conditions for commercial agriculture: an example for potato production in Michigan. Clim Change, 151 (2), 275–287, doi:10.1007/s10584-018-2301-4.
Witasari, A.I., 2016: Trust building to coordinate collective action in reforestation program. Manajemen Hutan Trop. , 22 (3), 192–201, doi:10.7226/jtfm.22.3.192.
Wolde, A., T. Amsalu and M.M. Alemu, 2016: Social and economic impacts of community managed reforestation and natural regeneration of forestry development, the case of Humbo district, Ethiopia. Environ. Nat. Resour. Res. , 6 (4), 36, doi:10.5539/enrr.v6n4p36.
Wolz, K.J., et al., 2018: Frontiers in alley cropping: Transformative solutions for temperate agriculture. Glob Chang Biol, 24 (3), 883–894, doi:10.1111/gcb.13986.
Wood, S.A. and F. Baudron, 2018: Soil organic matter underlies crop nutritional quality and productivity in smallholder agriculture. Agric Ecosyst Environ, 266, 100–108, doi:10.1016/j.agee.2018.07.025.
Woodard, J.D., B. Sherrick, J. Coppess and D.J. Muth, 2019: The Role of the Banking and Financing Sector in Encouraging Conservation Practices and Organic Transitions Meridian Institute. Available at: https://foodandagpolicy.org/wp-content/uploads/sites/4/2019/09/2019-March-Role-of-Banking-and-Finance-in-Encouraging-Conservation-Practices.pdf (accessed August 31, 2021)Woodworth-Jefcoats, P.A., J.L. Blanchard and J.C. Drazen, 2019: Relative Impacts of Simultaneous Stressors on a Pelagic Marine Ecosystem. Front. Mar. Sci. , 6, doi:10.3389/fmars.2019.00383.
Woolf, D., D. Solomon and J. Lehmann, 2018: Land restoration in food security programmes: synergies with climate change mitigation. Clim. Policy, 18 (10), 1260–1270, doi:10.1080/14693062.2018.1427537.
Woolway, R.I. and S.C. Maberly, 2020: Climate velocity in inland standing waters. Nat. Clim. Chang. , 10 (12), 1124–1129, doi:10.1038/s41558-020-0889-7.
Work, C., V. Rong, D. Song and A. Scheidel, 2019: Maladaptation and development as usual? Investigating climate change mitigation and adaptation projects in Cambodia. Clim. Policy, 19 (sup1), 47–S62, doi:10.1080/14693062.2018.1527677.
World Bank, 2010: Economics of adaptation to climate change - Synthesis report . World Bank, Washington, D.C., http://documents.worldbank.org/curated/en/646291468171244256/Economics-of-adaptation-to-climate-change-Synthesis-report.(accessed August 31, 2021)
World Bank, 2013: Fish to 2030: Prospects for Fisheries and Aquaculture. Agriculture and Environmental Services Discussion Paper, Vol. 3, 1. World Bank Group, Washington DC, USA. 80 pp.
World Bank, 2016: Forest Action Plan Fy16-20. Vol. 1. World Bank Group, Washington D.C., USA. 89 pp.
World Bank, 2018: Realigning Agricultural Support to Promote Climate-Smart Agriculture. Agriculture Global Practice Note, Vol. 1. World Bank Group, Washington D.C., USA. 4 pp.
World Bank, 2019: The World Governance Indicators. Available at: https://info.worldbank.org/governance/wgi/ (accessed May 5, 2020).
World Bank, 2020: The World Development Indicators: GDP (current US$). Available at: https://data.worldbank.org/indicator/NY.GDP.MKTP.CD (accessed June 26, 2021).Wreford, A. and C.F.E. Topp, 2020: Impacts of climate change on livestock and possible adaptations: A case study of the United Kingdom. Agric Syst , 178, 102737, doi:10.1016/j.agsy.2019.102737.
WRI, 2018: Creating a Sustainable Food Future: A Menu of Solutions to Feed Nearly 10 Billion People by 2050. World Resources Institute, Available at: https://www.wri.org/publication/creating-sustainable-food-future (accessed August 31, 2021).
Wu, J., 2016: The distributions of Chinese yak breeds in response to climate change over the past 50 years. Anim. Sci. J. , 87 (7), 947–958, doi:10.1111/asj.12526.
Wu, J.G., 2015: The response of the distributions of Asian buffalo breeds in China to climate change over the past 50 years. Livest. Sci. , 180, 65–77, doi:10.1016/j.livsci.2015.07.005.
Wu, K.S., Y.R. He, Q.J. Chen and Y.M. Zheng, 2020: Analysis on the damage and recovery of typhoon disaster based on UAV orthograph. Microelectron. Reliab. , 107, doi:10.1016/j.microrel.2019.06.029.
Xiang, J., A. Hansen, D. Pisaniello and P. Bi, 2016: Workers’ perceptions of climate change related extreme heat exposure in South Australia: a cross-sectional survey. BMC Public Health, 16, 549, doi:10.1186/s12889-016-3241-4.
Xin, Y. and F. Tao, 2019: Optimizing genotype-environment-management interactions to enhance productivity and eco-efficiency for wheat-maize rotation in the North China Plain. Sci. Total Environ. , 654, 480–492, doi:10.1016/j.scitotenv.2018.11.126.
Xinhua, Y., et al., 2017: Fishery and Aquaculture Insurance in China. FAO Fisheries and Aquaculture Circular, Vol. 1139. FAO, Rome, 1–30.
Xu, H.Q., et al., 2019: Future increases in irrigation water requirement challenge the water-food nexus in the northeast farming region of China. Agric. Water Manag. , 213, 594–604, doi:10.1016/j.agwat.2018.10.045.
Xu, J., et al., 2008: Understanding land use, livelihoods, and health transitions among Tibetan nomads: a case from Gangga Township, Dingri County, Tibetan Autonomous Region of China. EcoHealth, 5 (2), 104–114, doi:10.1007/s10393-008-0173-1.
Yamamoto, T., et al., 2018: Application and development of genome editing technologies to the Solanaceae plants. Plant. Physiol. Biochem. , 131, 37–46, doi:10.1016/j.plaphy.2018.02.019.
Yan, Y.J., et al., 2017: Range shifts in response to climate, change of Ophiocordyceps sinensis, a fungus endemic to the Tibetan Plateau. Biol. Conserv. , 206, 143–150, doi:10.1016/j.biocon.2016.12.023.
Yang, G., J. Roy, S.D. Veresoglou and M.C. Rillig, 2021: Soil biodiversity enhances the persistence of legumes under climate change. New Phytol. , 229 (5), 2945–2956, doi:10.1111/nph.17065.
Yang, J., et al., 2018: Quantifying the Sustainability of Water Availability for the Water-Food-Energy-Ecosystem Nexus in the Niger River Basin. Earths Future, 6 (9), 1292–1310, doi:10.1029/2018EF000923.
Yang, X.F., et al., 2012: Climate change effects fruiting of the prize matsutake mushroom in China. Fungal Divers. , 56 (1), 189–198, doi:10.1007/s13225-012-0163-z.
Yang, Y.M., et al., 2014: Prediction of cotton yield and water demand under climate change and future adaptation measures. Agric. Water Manag. , 144, 42–53, doi:10.1016/j.agwat.2014.06.001.
Yeboah, T., et al., 2020: Hard work and hazard: Young people and agricultural commercialisation in Africa. J Rural Stud, 76, 142–151, doi:10.1016/j.jrurstud.2020.04.027.
Yengoh, G.T. and F.A. Armah, 2016: Land access constraints for communities affected by large-scale land acquisition in Southern Sierra Leone. GeoJournal, 81 (1), 103–122, doi:10.1007/s10708-014-9606-2.
Yengoh, G.T., F.A. Armah and K. Steen, 2015: Women’s Bigger Burden: Disparities in Outcomes of Large Scale Land Acquisition in Sierra Leone. Gend. Issues, 32 (4), 221–244, doi:10.1007/s12147-015-9140-7.
Yeni, F. and H. Alpas, 2017: Vulnerability of global food production to extreme climatic events. Food. Res. Int. , 96 (27), 27–39, doi:10.1016/j.foodres.2017.03.020.
Yetty, Y., B. Hariyadi and P. Murni, 2013: Studi Etnobotani Jernang (Daemonorops spp.) pada Masyarakat Desa Lamban Sigatal dan Sepintun Kecamatan Pauh Kabupaten Sarolangun Jambi. Biospecies, 6 (1), 38-43.
Yi, Y.J., X. Cheng, Z.F. Yang and S.H. Zhang, 2016: Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol Eng, 92, 260–269, doi:10.1016/j.ecoleng.2016.04.010.
Yi, Y.J., et al., 2018: The influence of climate change on an endangered riparian plant species: The root of riparian Homonoia. Ecol Indic, 92, 40–50, doi:10.1016/j.ecolind.2017.05.004.
Yokohata, T. et al., 2020: MIROC-INTEG-LAND version 1 : a global biogeochemical land surface model with human water management , crop growth , and land-use change. Geosci. Model Dev. , 13 (10), 4713-4747, doi:10.5194/gmd-13-4713-2020.
York, L., C. Heffernan, C. Rymer and N. Panda, 2017: A deterministic evaluation of heat stress mitigation and feed cost under climate change within the smallholder dairy sector. Animal, 11 (5), 900–909, doi:10.1017/S1751731116002706.
Young, H. and M. A. Ismail, 2019: Complexity, continuity and change: livelihood resilience in the Darfur region of Sudan. Disasters, 43 (Suppl 3), S318–S344, doi:10.1111/disa.12337.
Young, N. and M. Liston, 2010: (Mis)managing a risk controversy: the Canadian salmon aquaculture industry’s responses to organized and local opposition. J. Risk Res. , 13 (8), 1043–1065, doi:10.1080/13669877.2010.514429.
Young, T., et al., 2019: Adaptation strategies of coastal fishing communities as species shift poleward. ICES J. Mar. Sci. , 76 (1), 93–103, doi:10.1093/icesjms/fsy140.
Yousefi, A., et al., 2020: Biometric indices of wild pistachio (Pistacia atlantica Desf.) trees under resin extraction in Western Iran. Agrofor. Syst. , 94 (5), 1977–1988.
Yu, H., et al., 2019: Targeted gene disruption in pacific oyster based on CRISPR/Cas9 Ribonucleoprotein complexes. Mar Biotechnol (ny) , 21 (3), 301–309, doi:10.1007/s10126-019-09885-y.
Zabaniotou, A., 2018: Redesigning a bioenergy sector in EU in the transition to circular waste-based Bioeconomy-A multidisciplinary review. J Clean Prod, 177, 197–206, doi:10.1016/j.jclepro.2017.12.172.
Zabel, F., et al., 2021: Large potential for crop production adaptation depends on available future varieties. Glob Chang Biol, doi:10.1111/gcb.15649.
Zahra, W., et al., 2020: Economic Importance of Medicinal Plants in Asian Countries. In: Bioeconomy for Sustainable Development . [Keswani, C. (ed.)]. Springer, Singapore, pp. 359-377.
Zaidi, S.S., et al., 2019: New plant breeding technologies for food security. Science, 363 (6434), 1390–1391, doi:10.1126/science.aav6316.
Zalesny, R.S., et al., 2019: Positive water linkages of producing short rotation poplars and willows for bioenergy and phytotechnologies. Wiley Interdiscip. Rev. Energy Environ. , 8 (5), doi:10.1002/wene.345.
Zampieri, M., A. Ceglar, F. Dentener and A. Toreti, 2017: Wheat yield loss attributable to heat waves, drought and water excess at the global, national and subnational scales. Environ. Res. Lett. , 12 (6), doi:10.1088/1748-9326/aa723b.
Zandalinas, S.I., et al., 2018: Plant adaptations to the combination of drought and high temperatures. Physiol Plant , 162 (1), 2–12, doi:10.1111/ppl.12540.
Zander, K.K., et al., 2015: Heat stress causes substantial labour productivity loss in Australia. Nat. Clim. Chang. , 5 (7), 647, doi:10.1038/Nclimate2623.
Zavaleta, C., et al., 2018: Multiple non-climatic drivers of food insecurity reinforce climate change maladaptation trajectories among Peruvian Indigenous Shawi in the Amazon. PLoS ONE, 13 (10), e205714, doi:10.1371/journal.pone.0205714.
Zellweger, F., et al., 2020: Forest microclimate dynamics drive plant responses to warming. Science, 368 (6492), 772–775, doi:10.1126/science.aba6880.
Zenger, K.R., et al., 2018: Genomic selection in aquaculture: application, limitations and opportunities with special reference to marine shrimp and pearl oysters. Front. Genet. , 9 (693), 693, doi:10.3389/fgene.2018.00693.
Zhang, K. et al., 2019a: Unexpected groundwater recovery with decreasing agricultural irrigation in the Yellow River Basin. Agric. Water Manage., 213, 858-867, doi:10.1016/j.agwat.2018.12.009.
Zhang, L., et al., 2016: Ethnobotanical study of traditional edible plants used by the Naxi people during droughts. J Ethnobiol Ethnomed, 12 (1), 39, doi:10.1186/s13002-016-0113-z.
Zhang, S., F.L. Tao and Z. Zhang, 2014: Rice reproductive growth duration increased despite of negative impacts of climate warming across China during 1981-2009. Eur J Agron, 54, 70–83, doi:10.1016/j.eja.2013.12.001.
Zhang, S.Z., et al., 2017a: Effects of elevated CO2 concentration and temperature on some physiological characteristics of cotton (Gossypium hirsutum L.) leaves. Environ. Exp. Bot. , 133, 108–117, doi:10.1016/j.envexpbot.2016.10.001.
Zhang, Y., A.A. Malzahn, S. Sretenovic and Y. Qi, 2019b: The emerging and uncultivated potential of CRISPR technology in plant science. Nat. Plants, 5 (8), 778–794, doi:10.1038/s41477-019-0461-5.
Zhang, Y., Y. Wang and H. Niu, 2017b: Spatio-temporal variations in the areas suitable for the cultivation of rice and maize in China under future climate scenarios. Sci. Total Environ. , 601-602, 518–531, doi:10.1016/j.scitotenv.2017.05.232.
Zhang, Y.Q.W., B.A. McCarl, Y.B. Luan and U. Kleinwechter, 2018: Climate change effects on pesticide usage reduction efforts: a case study in China. Mitig Adapt Strateg Glob Change, 23 (5), 685–701, doi:10.1007/s11027-017-9755-y.
Zhao, C., et al., 2017: Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA, 114 (35), 9326–9331, doi:10.1073/pnas.1701762114.
Zhao, C., et al., 2019: A SIMPLE crop model. Eur J Agron, 104, 97–106, doi:10.1016/j.eja.2019.01.009.
Zhao, Q., et al., 2018: Modeling impacts of climate change on the geographic distribution of medicinal plant <i>Fritillaria cirrhosa</i> D. Don. Plant Biosyst. - Int. J. Deal. With All Aspects Plant Biol. , 152, 349–355, doi:10.1080/11263504.2017.1289273.
Zhao, Z., et al., 2020: Potential distribution of Notopterygium incisum Ting ex H. T. Chang and its predicted responses to climate change based on a comprehensive habitat suitability model. Ecol Evol, 10 (6), 3004–3016, doi:10.1002/ece3.6117.
Zhou, X., et al., 2014: Nutrient removal by prairie filter strips in agricultural landscapes. J. Soil Water Conserv. , 69 (1), 54–64, doi:10.2489/jswc.69.1.54.
Zhu, C., et al., 2018a: Carbon dioxide (CO2) levels this century will alter the protein, micronutrients, and vitamin content of rice grains with potential health consequences for the poorest rice-dependent countries. Sci. Adv. , 4 (5), eaaq1012, doi:10.1126/sciadv.aaq1012.
Zhu, P., et al., 2018b: The important but weakening maize yield benefit of grain filling prolongation in the US Midwest. Glob Chang Biol, 24 (10), 4718–4730, doi:10.1111/gcb.14356.
Zickfeld, K., et al., 2013: Long-term climate change commitment and reversibility: an EMIC Intercomparison. J Clim, 26 (16), 5782–5809, doi:10.1175/Jcli-D-12-00584.1.
Ziervogel, G., 2019: Building transformative capacity for adaptation planning and implementation that works for the urban poor: Insights from South Africa. Ambio, 48 (5), 494–506, doi:10.1007/s13280-018-1141-9.
Zimová, S., et al., 2020: Reducing rotation age to address increasing disturbances in Central Europe: Potential and limitations. For. Ecol. Manag. , 475, doi:10.1016/j.foreco.2020.118408.
Zin, W.Y.L., P. Teartisup and P. Kerdseub, 2019: Evaluating traditional knowledge on climate change (TKCC): a case study in the central dry zone of Myanmar. Environ. Nat. Resour. J. , 17 (2), 1–29, doi:10.32526/ennrj.17.2.2019.09.
Zinsstag, J., et al., 2018: Climate change and One Health. FEMS Microbiol. Lett. , 365 (11), doi:10.1093/femsle/fny085.
Ziska, L.H., 2003: Evaluation of the growth response of six invasive species to past, present and future atmospheric carbon dioxide. J. Exp. Bot. , 54 (381), 395–404, doi:10.1093/jxb/erg027.
Ziska, L.H., 2014: Increasing minimum daily temperatures are associated with enhanced pesticide use in cultivated soybean along a latitudinal gradient in the mid-western United States. PLoS One, 9 (2), e98516, doi:10.1371/journal.pone.0098516.
Ziska, L.H., 2016: The role of climate change and increasing atmospheric carbon dioxide on weed management: Herbicide efficacy. Agric Ecosyst Environ, 231, 304–309, doi:10.1016/j.agee.2016.07.014.
Ziska, L.H., 2020: Climate change and the herbicide paradigm: visiting the future. Agronomy, 10 (12), doi:10.3390/agronomy10121953.
Ziska, L.H. and L.L. McConnell, 2016: Climate change, carbon dioxide, and pest biology: monitor, mitigate, manage. J. Agric. Food Chem. , 64 (1), 6–12, doi:10.1021/jf506101h.
Ziska, L.H., et al., 2016: Rising atmospheric CO2 is reducing the protein concentration of a floral pollen source essential for North American bees. Proc. Biol. Sci. , 283 (1828), 20160414, doi:10.1098/rspb.2016.0414.
Zomer, R.J., et al., 2016: Global tree cover and biomass carbon on agricultural land: the contribution of agroforestry to global and national carbon budgets. Sci Rep, 6, 29987, doi:10.1038/srep29987.
Zoomers, E.B. and K. Otsuki, 2017: Addressing the impacts of large-scale land investments: Re-engaging with livelihood research. Geoforum, 83, 164–171, doi:10.1016/j.geoforum.2017.01.009.
Zougmoré, R., et al., 2016: Toward climate-smart agriculture in West Africa: a review of climate change impacts, adaptation strategies and policy developments for the livestock, fishery and crop production sectors. Agric. Food Secur. , 5 (1), doi:10.1186/s40066-016-0075-3.
Zouré, C., et al., 2019: Modelling the water balance on farming practices at plot scale: Case study of Tougou watershed in Northern Burkina Faso. CATENA, 173, 59–70, doi:10.1016/j.catena.2018.10.002.
Zumpf, C., et al., 2017: Yield and water quality impacts of field-scale integration of willow into a continuous corn rotation system. J. environ. qual. , 46 (4), 811–818, doi:10.2134/jeq2017.02.0082.
1 In this Report, the following summary terms are used to describe the available evidence: limited, medium, or robust; and for the degree of agreement: low, medium, or high. A level of confidence is expressed using five qualifiers: very low, low, medium, high, and very high, and typeset in italics, e.g., medium confidence. For a given evidence and agreement statement, different confidence levels can be assigned, but increasing levels of evidence and degrees of agreement are correlated with increasing confidence.
2 2 In this Report, the following terms have been used to indicate the assessed likelihood of an outcome or a result: virtually certain 99–100% probability, very likely 90–100%, likely 66–100%, about as likely as not 33–66%, unlikely 0–33%, very unlikely 0–10%, and exceptionally unlikely 0–1%. Additional terms (extremely likely 95–100%, more likely than not >50–100%, and extremely unlikely 0–5%) may also be used when appropriate. Assessed likelihood is typeset in italics, e.g., very likely . This Report also uses the term ‘likely range’ to indicate that the assessed likelihood of an outcome lies within the 17–83% probability range.
3 Disability-adjusted life years or DALYs.
4 For reference, the SEI Aid-Atlas (https://aid-atlas.org) only reports flows where adaptation is the principal objective, and therefore adaptation spending on agriculture, forestry and fisheries for the same period is significantly lower with USD 16.52 billion, that is, 21.4% of total adaptation spending.
Table of Contents
5.1.1 Scope of the Chapter 720
5.1.2 Starting Point: AR5 and Recent IPCC Special Reports 720
5.2 Observed Impacts and Key Risks 723
5.2.1 Detection and Attribution of Observed Impacts 723
5.3 Methodologies and Associated Uncertainties 725
5.3.1 Methodologies for Assessing Impacts and Risks 725
5.3.2 Methodologies for Assessing Vulnerabilities and Adaptation 727
Box 5.1: Evidence for Simultaneous Crop Failures Due to Climate Change 731
5.4.2 Assessing Vulnerabilities within Production Systems 733
5.5 Livestock-Based Systems 746
5.5.2 Assessing Vulnerabilities 747
5.5.4 Adaptation in Livestock-Based Systems 751
Box 5.5: Alternative Sources of Protein for Food and Feed 753
Box 5.6: Contributions of Indigenous and Local Knowledge: An Example 755
5.7 Other Natural Products 757
5.7.4 Observed and Projected Impacts 758
5.8 Ocean-Based and Inland Fisheries Systems 760
5.8.2 Assessing Vulnerabilities 764
Cross-Chapter Box: MOVING PLATE: Sourcing Food When Species Distributions Change 768
5.9 Ocean-Based and Inland Aquaculture Systems 773
5.9.2 Assessing Vulnerabilities 774
5.9.4 Aquaculture Adaptation 779
5.9.5 Contributions of Indigenous, Traditional and Local Knowledge 782
5.10.2 Assessing Vulnerabilities 783
Box 5.7: Perspectives of Crop and Livestock Farmers on Observed Changes in Climate in the Sahel 785
5.10.4 Adaptation Strategies 786
5.11 The Supply Chain from Post-harvest to Food 787
Box 5.8: Climate Adaptation and Maladaptation in Cocoa and Coffee Production 788
5.11.1 Current and Future Climate Change Impacts on Food Safety 789
5.11.4 Adaptation in the Post-harvest Supply Chain 791
5.12 Food Security, Consumption and Nutrition 792
5.12.2 Mechanisms for Climate Change Impacts on Food Security 794
Box 5.9: Desert Locust Case Study: Climate as Compounding Effect on Food Security 795
5.12.4 Projected Impacts on Food Security 796
Box 5.10: Food Safety Interactions with Food Security and Malnutrition 798
5.12.5 Adaptation Options for Food Security and Nutrition 798
5.12.6 Changing Dietary Patterns 799
5.12.7 Integrated Multisectoral Food Security and Nutrition Adaptation Options 801
5.12.8 Incorporating Human Rights-Based Approaches into Food Systems 801
5.13 Climate-Change-Triggered Competition, Trade-offs and Nexus Interactions in Land and Ocean 802
5.13.2 Trade-offs Generated by Agricultural Intensification and Expansion 804
5.13.3 Competition between Food Systems in Land and Ocean 805
5.13.4 Maladaptation Responses and Sustainable Solutions 806
Case Studies of Maladaptation 806
Large-Scale Irrigation Project in Navarre, Spain 806
Constraining Adaptation: Previous Agricultural Development Pathways in India 806
5.13.5 Climate Change and Climate Response Impacts on Indigenous People 806
5.13.6 Increased Presence of Financial Actors in the Agrifood System 810
5.13.7 Climate Change Interactions with other Drivers—Food–Water–Health–Energy–Security Nexus 811
5.14 Implementation Pathways to Adaptation and Co-benefits 811
5.14.1 State of Adaptation of Food, Feed, Fibre and Other Ecosystem Products 811
Box 5.11: Agroecology as a Transformative Climate Change Adaptation Approach 815
5.14.2 Enabling Conditions for Implementing Adaptation 818
Box 5.12: Is Climate-Smart Agriculture Overlooking Gender and Power Relations?819
Box 5.13: Supporting Youth Adaptation in Food Systems 820
5.14.3 Climate Resilient Development Pathways 825
Cross-Working Group Box BIOECONOMY: Mitigation and Adaptation via the Bioeconomy 828
FAQ 5.4: What are effective adaptation strategies for improving food security in a warming world?836





